|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ГЛАВА 1. МОРФОФУНКЦИОНАЛЬНЫЕ ОСНОВЫ НЕРВНОЙ СИСТЕМЫ1.1. Нервная система: общая структура Нервная система - это система организма, объединяющая и регулирующая различные физиологические процессы в соответствии с меняющимися условиями внешней и внутренней среды. Нервная система состоит из сенсорных компонентов, реагирующих на раздражители, исходящие из окружающей среды, интегративных - перерабатывающих и хранящих сенсорные и другие данные, и двигательных, управляющих движениями и секреторной деятельностью желез. Нервная система воспринимает сенсорные стимулы, перерабатывает информацию и формирует поведение. Особые виды переработки информации - научение и память, благодаря которым при изменениях окружающей среды поведение адаптируется с учетом предшествующего опыта. В этих функциях участвуют и другие системы, такие как эндокринная и иммунная, но нервная система специализирована для выполнения указанных функций. Под переработкой информации подразумевают передачу информации в нейронных сетях, трансформацию сигналов путем их объединения с другими сигналами (нервная интеграция), хранение информации в памяти и извлечение информации из памяти, использование сенсорной информации для восприятия, мышление, обучение, планирование (подготовка) и выполнение двигательных команд, формирование эмоций. Взаимодействия между нейронами осуществляются посредством как электрических, так и химических процессов. Поведение - это комплекс реакций организма на меняющиеся условия внешней и внутренней среды. Поведение может быть сугубо внутренним, скрытым процессом (познание) или доступным для внешнего наблюдения (двигательные или вегетативные реакции). У человека особенно важен набор тех поведенческих актов, которые связаны с речью. Каждую реакцию, простую или комплексную, обеспечивают нервные клетки, организованные в нейронные сети (нервные ансамбли и пути). Нервную систему подразделяют на центральную и периферическую (рис. 1.1). Центральная нервная система (ЦНС) состоит из головного и спинного мозга. В периферическую нервную систему входят корешки, сплетения и нервы.
А - Центральная нервная система. Б - Ствол мозга: 1 - конечный мозг; 2 - промежуточный мозг; 3 - средний мозг; 4 - мост и мозжечок, 5 - продолговатый мозг, 6 - конечный мозг срединные структуры. В - Спинной мозг: 7 - спинномозговой конус; 8 - терминальные нити. Г - Периферическая нервная система: 9 - вентральный корешок; 10- дорзальный корешок; 11 - спинальный ганглий; 12 - спинномозговой нерв; 13 - смешанный периферический нерв; 14 - эпиневрий; 15 - периневрий; 16 - миелиновый нерв; 17 - фиброцит; 18 - эндоневрий; 19 - капилляр; 20 - безмиелиновый нерв; 21 - кожные рецепторы; 22 - окончание мотонейрона; 23 - капилляр; 24 - мышечные волокна; 25 - ядро шванновской клетки; 26 - перехват Ранвье; 27 - симпатический ствол; 28 - соединительная ветвь Центральная нервная система ЦНС собирает и перерабатывает поступающую от рецепторов информацию об окружающей среде, формирует рефлексы и другие поведенческие реакции, планирует и осуществляет произвольные движения. Кроме того, ЦНС обеспечивает так называемые высшие познавательные (когнитивные) функции. В ЦНС происходят процессы, связанные с памятью, обучением и мышлением. Головной мозг в процессе онтогенеза формируется из мозговых пузырей, возникающих в результате неравномерного роста передних отделов медуллярной трубки (рис. 1.2). Из этих пузырей формируются передний мозг (prosencephalon), средний мозг (mesencephalon) и ромбовидный мозг (rhombencephalon). В дальнейшем из переднего мозга образуются конечный (telencephalon) и промежуточный (diencephalon) мозг, а ромбовидный мозг разделяется на задний (metencephalon) и продолговатый (myelencephalon, или medulla oblongata) мозг. Из конечного мозга, соответственно, формируются полушария большого мозга, базальные ганглии, из промежуточного мозга - таламус, эпиталамус, гипоталамус, метаталамус, зрительные тракты и нервы, сетчатка. Зрительные нервы и сетчатка - отделы ЦНС, как бы вынесенные за пределы головного мозга. Из среднего мозга образуются пластинка четверохолмия и ножки мозга. Из заднего мозга формируются мост и мозжечок. Мост мозга граничит внизу с продолговатым мозгом. Задняя часть медуллярной трубки формирует спинной мозг, а ее полость превращается в центральный канал спинного мозга. Спинной мозг состоит из шейного, грудного, поясничного, крестцового и копчикового отделов, каждый из которых в свою очередь складывается из сегментов. В ЦНС различают серое и белое вещество. Серое вещество представляет собой скопление тел нейронов, белое вещество - отростки нейронов, покрытые миелиновой оболочкой. В головном мозге серое вещество находится в коре больших полушарий, в подкорковых ганглиях, ядрах ствола мозга, коре мозжечка и его ядрах. В спинном мозге серое вещество концентрируется в его середине, белое - на периферии. Периферическая нервная система Периферическая нервная система (ПНС) отвечает за сопряжение между окружающей средой (или возбудимыми клетками) и ЦНС. В состав ПНС входят сенсорные (рецепторы и первичные афферентные нейроны) и двигательные (соматические и вегетативные мотонейроны) компоненты.
Б. I - Общий вид сбоку. Б. II - Вид сверху: 10 - конечный мозг; 11 - боковой желудочек; 12 - промежуточный мозг; 13 - глазной стебелек; 14 - хрусталик; 15 - зрительный нерв; 16 - средний мозг; 17 - задний мозг; 18 - продолговатый мозг; 19 - спинной мозг; 20 - центральный канал; 21 - четвертый желудочек; 22 - водопровод мозга; 23 - третий желудочек. III - Вид сбоку: 24 - новая кора; 25 - межжелудочковая перегородка; 26 - полосатое тело; 27 - бледный шар; 28 - гиппокамп; 29 - таламус; 30 - шишковидное тело; 31 - верхние и нижние холмики; 32 - мозжечок; 33 - задний мозг; 34 - спинной мозг; 35 - продолговатый мозг; 36 - мост; 37 - средний мозг; 38 - нейрогипофиз; 39 - гипоталамус; 40 - миндалевидное тело; 41 - обонятельный тракт; 42 - обонятельная кора Сенсорная часть ПНС. Сенсорное восприятие - это преобразование энергии внешнего стимула в нервный сигнал. Оно осуществляется специализированными структурами - рецепторами, воспринимающими воздействие на организм разнообразных видов внешней энергии, включая механическую, свет, звук, химические стимулы, изменения температуры. Рецепторы расположены на периферических окончаниях первичных афферентных нейронов, передающих получаемую информацию в ЦНС по чувствительным волокнам нервов, сплетений, спинальных нервов и, наконец, по задним корешкам спинного мозга (либо по черепным нервам). Тела клеток задних корешков и черепных нервов находятся в спинномозговых (спинальных) ганглиях либо в ганглиях черепных нервов. Двигательная часть ПНС. К двигательному компоненту ПНС относятся соматические и вегетативные (автономные) мотонейроны. Соматические мотонейроны иннервируют поперечнополосатую мускулатуру. Тела клеток находятся в передних рогах спинного мозга или в стволе мозга, у них длинные дендриты, получающие много синаптических «входов». Мотонейроны каждой мышцы составляют определенное двигательное ядро - группу нейронов ЦНС, имеющих сходные функции. Например, от ядра лицевого нерва иннервируются мышцы лица. Аксоны соматических мотонейронов покидают ЦНС через передний корешок либо через черепной нерв. Вегетативные (автономные) мотонейроны посылают нервы к волокнам гладкой мускулатуры и к железам - преганглионарные и постганглионарные нейроны симпатической и парасимпатической нервной системы. Преганглионарные нейроны расположены в ЦНС - в спинном мозге либо в стволе мозга. В отличие от соматических мотонейронов, вегетативные преганглионарные нейроны образуют синапсы не на эффекторных клетках (гладкой мускулатуре или железах), а на постганглионарных нейронах, которые в свою очередь синаптически контактируют непосредственно с эффекторами. 1.2. Микроскопическое строение нервной системы Основу нервной системы составляют нервные клетки, или нейроны, специализирующиеся на получении входящих сигналов и на передаче сигналов к другим нейронам или эффекторным клеткам. Кроме нервных клеток, в нервной системе имеются глиальные клетки и элементы соединительной ткани. Клетки нейроглии (от греч. «глия» - клей) вы- полняют в нервной системе опорные, трофические, регуляторные функции, участвуя практически во всех видах активности нейронов. Количественно они преобладают над нейронами и занимают весь объем между сосудами и нервными клетками. Нервная клетка Основной структурно-функциональной единицей нервной системы является нейрон (рис. 1.3). В нейроне различают тело (сому) и отростки: дендриты и аксон. Сома и дендриты представляют воспринимающую поверхность клетки. Аксон нервной клетки образует синаптические связи с другими нейронами или с эффекторными клетками. Нервный импульс распространяется всегда в одном направлении: по дендритам к телу клетки, по аксону - от тела клетки (закон динамической поляризации нервной клетки Рамон-иКахаля). Как правило, нейрон имеет множество «входов», осуществляемых дендритами, и только один «выход» (аксон) (см. рис. 1.3). Нейроны сообщаются друг с другом с помощью потенциалов действия, которые распространяются по аксонам. Потенциалы действия поступают от одного нейрона к следующему в результате синаптической передачи. Достигший пресинаптического окончания потенциал действия обычно запускает высвобождение нейромедиатора, который либо возбуждает постсинаптическую клетку, так что в ней возникает разряд из одного или нескольких потенциалов действия, либо тормозит ее активность. Аксоны не только передают информацию в нервных
цепях, но и доставляют путем аксонного транспорта химические вещества к синаптическим окончаниям. Существуют многочисленные классификации нейронов в соответствии с формой их тела, протяженностью и формой дендритов и другими признаками (рис. 1.4). По функциональному значению нервные клетки подразделяются на афферентные (чувствительные, сенсорные), доставляющие импульсы к центру, эфферентные (двигательные, моторные), несущие информацию от центра к периферии, и вставочные (интернейроны), в которых происходит переработка импульсов и организуются коллатеральные связи. Нервная клетка осуществляет две основные функции: специфическую переработку поступающей информации и передачу нервного импульса и биосинтетическую, направленную на поддержание своей жизнедеятельности. Это находит выражение и в ультраструктуре нервной клетки. Передача информации от одной нервной клетки к другой, объединение нервных клеток в системы и комплексы различной сложности осуществляются структурами нейрона: аксонами, дендритами и синапсами. Органеллы, связанные с обеспечением энергетического обмена, белоксинтезирующей функцией клетки, встречаются в большинстве клеток; в нервных клетках они выполняют функции энергообеспечения клетки, переработки и передачи информации (см. рис. 1.3). Структура нейрона. Сома. Тело нервной клетки имеет округлую или овальную форму, в центре (или слегка эксцентрично) располагается ядро. Оно содержит ядрышко и окружено наружной и внутренней ядерными мембранами толщиной около 70 Ǻ каждая, разделенных пери-
А - Псевдоуниполярный нейрон. Б - клетка Пуркинье (дендриты, аксон). В - пирамидная клетка (аксон). Г - мотонейрон переднего рога (аксон) нуклеарным пространством, размеры которого вариабельны. В кариоплазме распределены глыбки хроматина, локализованные преимущественно у внутренней ядерной мембраны. В цитоплазме нервных клеток располагаются элементы зернистой и незернистой цитоплазматичсской сети, полисомы, рибосомы, митохондрии, лизосомы, многопузырчатые тельца и другие органеллы (рис. 1.5). К аппарату биосинтеза в нейронах относятся тельца Ниссля - плотно прилегающие друг к другу сплюснутые цистерны гранулярного эндоплазматического ретикулума, а также хорошо выраженный аппарат Гольджи. Кроме того, сома содержит многочисленные митохондрии, определяющие ее энергетический обмен, и элементы цитоскелета, в том числе нейрофиламенты и микротрубочки. Лизосомы и фагосомы являются основными органеллами «внутриклеточного пищеварительного тракта». Дендриты. Дендриты и их разветвления определяют рецептивное поле той или иной клетки (см. рис. 1.5). При электронно-микроскопическом исследовании обнаруживается, что тело нейрона постепенно переходит в дендрит. Резкой границы и выраженных различий в ультраструктуре сомы и начального отдела крупного дендрита не наблюдается. Дендриты очень вариабельны по форме, величине, разветвленности и ультраструктуре. Обычно от тела клетки отходит несколько дендритов. Длина дендрита может превышать 1 мм, на их долю приходится более 90% площади поверхности нейрона. Основными компонентами цитоплазмы дендритов являются микротрубочки и нейрофиламенты; в проксимальных частях дендритов (ближе к клеточному телу) содержатся тельца Ниссля и участки аппарата Гольджи. Ранее считалось, что дендриты электрически невозбудимы, в настоящее время доказано, что дендриты многих
1 - ядро; 2 - гранулярная эндоплазматическая сеть; 3 - пластинчатый комплекс (Гольджи); 4 - митохондрии; 5 - лизосомы; 6 - мультивезикулярное тело; 7 - полисомы нейронов обладают потенциалзависимой проводимостью, что обусловлено присутствием на их мембранах кальциевых каналов, при активации которых генерируются потенциалы действия. Аксон. Аксон берет начало у аксонного холмика - специализированного участка клетки (чаще сомы, но иногда - дендрита) (см. рис. 1.3). Аксон и аксонный холмик отличаются от сомы и проксимальных участков дендритов отсутствием гранулярного эндоплазматического ретикулума, свободных рибосом и аппарата Гольджи. В аксоне присутствуют гладкий эндоплазматический ретикулум и выраженный цитоскелет. Аксоны покрываются миелиновой оболочкой, образуя миелиновые волокна. Пучки волокон (в которых могут быть отдельные немиелинизированные волокна) составляют белое вещество мозга, черепные и периферические нервы. При переходе аксона в пресинаптическое окончание, наполненное синаптическими пузырьками, аксон образует колбовидное расширение. Переплетения аксонов, дендритов и отростков глиальных клеток создают сложные, не повторяющиеся картины нейропиля. Распределение аксонов и дендритов, их взаиморасположение, афферентно-эфферентные взаимоотношения, закономерности синаптоархитектоники определяют механизмы интегративной функции мозга. Типы нейронов. Полиморфизм строения нейронов определяется их различной ролью в системной деятельности мозга в целом. Так, нейроны ганглиев задних корешков спинного мозга (спинальных ганглиев) получают информацию не путем синаптической передачи, а от сенсорных нервных окончаний в рецепторных органах. В соответствии с этим клеточные тела этих нейронов лишены дендритов и не получают синаптических окончаний (биполярные клетки; рис. 1.6). Выйдя из клеточного тела, аксон такого нейрона разделяется на две ветви, одна из которых (периферический отросток) направляется в составе периферического нерва к рецептору, а другая ветвь (центральный отросток) входит в спинной мозг (в составе заднего корешка) либо в ствол мозга (в составе черепного нерва). Нейроны другого типа, такие, как пирамидные клетки коры больших полушарий и клетки Пуркинье коры мозжечка, заняты переработкой информации. Их дендриты покрыты дендритными шипиками и имеют обширную поверхность; к ним поступает огромное количество синаптических входов (мультиполярные клетки; см. рис. 1.4, 1.6). Можно классифицировать нейроны по длине их аксонов. У нейронов 1-го типа по Гольджи аксоны короткие, оканчивающиеся, так же как дендриты, близко к соме. Нейроны 2-го типа имеют длинные аксоны, иногда длиннее 1 м. Нейроглия Другая группа клеточных элементов нервной системы - нейроглия (рис. 1.7). В ЦНС человека число нейроглиальных клеток на порядок больше, чем число нейронов: 1013 и 1012 соответственно. Тесная морфологическая взаимосвязь является основой для физиологических и патологических взаимодействий глии и нейронов. Их взаимоотношения описываются концепцией динамических нейронально-глиальных сигнальных процессов. Возможность передачи сигналов от нейронов к глии и, таким образом, к другим нейронам открывает много вариантов для межклеточных «перекрестных разговоров». Существует несколько типов нейроглии, в ЦНС нейроглия представлена астроцитами и олигодендроцитами, а в ПНС - шванновскими клетками и клетками-сателлитами. Кроме того, центральными глиальными клетками считаются клетки микроглии и клетки эпендимы. Астроциты (получившие название благодаря своей звездчатой форме) регулируют состояние микросреды вокруг нейронов ЦНС. Их отростками окружены группы синаптических окончаний, которые в результате изолированы от соседних синапсов. Особые отростки - «ножки» астроцитов образуют контакты с капиллярами и соединительной тканью на поверхности головного и спинного мозга (мягкой мозговой оболочкой) (рис. 1.8). Ножки ограничивают свободную диффузию веществ в ЦНС. Астроциты могут активно поглощать К+ и нейромедиаторы, затем метаболизируя их. Благодаря избирательно повышенной проницаемости для ионов К+ астроглия регулирует активацию ферментов, необходимых для поддержания метаболизма нейронов, а также для удаления медиаторов и других агентов, выделяющихся в процессе нейро-
А -биполярный. Б - псевдоуниполярный. В - мультиполярный. 1 - дендриты; 2 - аксон
А - Протоплазматический астроцит. Б - микроглиальная клетка. В - олигодердроцит. Г - фиброзный астроцит нальной активности. Астроглия участвует в синтезе иммунных медиаторов: цитокинов, других сигнальных молекул (циклический гуанозинмонофосфат - сОМР, оксид азота - NO), передаваемых затем нейронам, - в синтезе глиальных ростовых факторов (GDNF), участвующих в трофике и репарации нейронов. Астроциты способны реагировать на увеличение синаптической концентрации нейротрансмиттеров и изменение электрической активности нейронов изменениями внутриклеточной концентрации Са2+. Это создает «волну» миграции Са2+ между астроцитами, способную модулировать состояние многих нейронов. Таким образом, астроглия, не являясь лишь трофическим компонентом нервной системы, участвует в специфическом функционировании нервной ткани. В цитоплазме астроцитов находятся глиальные филаменты, выполняющие в ткани ЦНС механическую опорную функцию. В случае повреждения отростки астроцитов, содержащие глиальные филаменты, подвергаются гипертрофии и формируют глиальный рубец. Основной функцией олигодендроцитов является обеспечение электрической изоляции аксонов путем формирования миелиновой оболочки (рис. 1.9). Это многослойная обертка, спирально намотанная поверх плазматической мембраны аксонов. В ПНС миелиновая оболочка образована мембранами шванновских клеток (см. рис. 1.18). Миелин представляет собой упаковку листов специфических плазматических мембран, богатых фосфолипидами, а также содержит протеины нескольких типов, различных в ЦНС и ПНС. Белковые структуры позволяют плазматическим мембранам плотно упаковываться вместе. При росте мембраны глиальной клетки происходит ее вращение вокруг аксона нейрона с образованием слоистой спирали с двойной плазматической мембраной вокруг аксона. Толщина миелиновой оболочки может составлять 50-100 мембран, играющих роль электрического изолятора аксона, предотвращающего ионообмен между цитозолем аксона и внеклеточной средой. Кроме того, в состав нейроглии входят клетки-сателлиты, которые инкапсулируют нейроны ганглиев спинальных и черепных нервов, регулируя микросреду вокруг этих нейронов наподобие того, как это делают астроциты (рис. 1.10). Еще один вид клеток - микроглия, или латентные фагоциты. Микроглия является единственным в ЦНС представительством иммунокомпетентных клеток. Она широко представлена во всей ткани мозга человека и составляет 9-12% общей глиальной популяции в сером веществе и 7,5-9% - в белом веществе. В отличие от астроцитов, микроглиальные клетки происходят из стволовых клеток и в нормальных условиях имеют вет-
1 - таницит; 2 - полость желудочка; 3 - эпендимальные клетки; 4 - капилляр; 5 - нейрон; 6 - миелинизированный аксон; 7 - мягкая мозговая оболочка; 8 - субарахноидальное пространство. На рисунке изображены два астроцита и их взаимосвязь с эпендимальными клетками, выстилающими желудочек, перикарионом, дендритами нейрона, капилляром, а также плоским эпителием мягкой мозговой оболочки. Следует отметить, что данный рисунок схематичен и связь нейрона одновременно с желудочком и субарахноидальным пространством маловероятна
вистую форму со множеством отростков. Активация микроглии, в частности в условиях гипоксииишемии, сопровождается продукцией провоспалительных медиаторов с токсичными свойствами. Поддерживаемая ими хроническая воспалительная реакция в ткани головного мозга ведет к отсроченным нейрональным потерям, микроциркуляторным нарушениям, изменениям функции гематоэнцефалического барьера. В патологических условиях микроглиальные клетки втягивают отростки и принимают амебоидную форму, чему соответствует их выраженная функциональная активация вплоть до состояния фагоцитоза. При повреждении мозговой ткани микроглия наряду с проникающими в ЦНС из кровотока фагоцитами способствует удалению продуктов клеточного распада. Ткань ЦНС отделена от цереброспинальной жидкости (ЦСЖ), заполняющей желудочки мозга, эпителием, который сформирован эпендимальными клетками. Эпендима обеспечивает диффузию многих веществ между внеклеточным пространством мозга и ЦСЖ. ЦСЖ секретируют специализированные эпендимальные клетки сосудистых сплетений в системе желудочков. Поступление к клеткам мозга питательных веществ и удаление продуктов жизнедеятельности клеток происходят через сосудистую систему. Хотя нервная ткань изобилует капиллярами и другими кровеносными сосудами, гематоэнцефалический барьер (ГЭБ) ограничивает диффузию многих веществ между кровью и тканью ЦНС. 1.3. Электрическая передача информации между нейронами Нормальная деятельность нервной системы зависит от возбудимости ее нейронов. Возбудимость - это способность клеточных мембран отвечать на действие адекватных раздражителей специфическими изменениями ионной проводимости и мембранного потенциала. Возбуждение - электрохимический процесс, идущий исключительно на цитоплазматической мембране клетки и характеризующийся изменениями ее электрического состояния, что запускает специфическую для каждой ткани функцию. Так, возбуждение мембраны мышцы вызывает ее сокращение, а возбуждение мембраны нейрона - проведение электрического сигнала по аксонам. Нейроны имеют не только потенциалуправляемые, т.е. регулируемые действием электрического возбудителя ионные каналы, но и хемоуправляемые и механоуправляемые. Существуют различия во взаимоотношениях между мембранным потенциалом/проницаемостью мембраны и типом раздражителя. При воздействии электрического раздражителя цепь событий следующая: раздражитель (электрический ток) => сдвиг мембранного потенциала (до критического потенциала) => активация потенциалуправляемых ионных каналов => изменение ионной проницаемости мембраны => изменение ионных токов через мембрану => дальнейший сдвиг мембранного потенциала (формирование потенциала действия). При воздействии химического раздражителя происходит принципиально другая цепь событий: раздражитель (химическое вещество) => химическое связывание раздражителя и рецептора хемоуправляемого ионного канала => изменение конформации лигандрецепторного комплекса и открытие рецепторуправляемых (хемоуправляемых) ионных каналов => изменение ионной проницаемости мембраны => изменение ионных токов через мембрану => сдвиг мембранного потенциала (формирование, например, локального потенциала). Цепь событий под воздействием механического раздражителя похожа на предыдущую, так как в этом случае также активируются рецепто- руправляемые ионные каналы: раздражитель (механический стресс) => изменение натяжения мембраны => открытие рецепторуправляемых (механоуправляемых) ионных каналов => изменение ионной проницаемости мембраны => изменение ионных токов через мембрану => сдвиг мембранного потенциала (формирование механоиндуцированного потенциала). Пассивные электрические свойства клетки связаны с электрическими свойствами ее мембраны, цитоплазмы и внешней среды. Электрические свойства клеточной мембраны определяются ее емкостными и резистивными характеристиками, так как липидный бислой впрямую можно уподобить и конденсатору, и резистору. Емкостные характеристики липидного бислоя и реальной мембраны сходны, а резистивные различаются вследствие присутствия прежде всего белков, образующих ионные каналы. У большинства клеток входное сопротивление ведет себя нелинейно: для тока, текущего в одном направлении, оно больше, чем для противоположно направленного. Это свойство асимметрии отражает активную реакцию и называется выпрямлением. Ток, протекающий через мембрану, определяется емкостным и резистивным компонентами. Резистивный компонент описывает собственно ионный ток, поскольку в клетке электричество переносится ионами. Движению ионов в клетку или из клетки препятствует плазматическая мембрана. Поскольку мембрана представляет липидный бислой, непроницаемый для ионов, она обладает сопротивлением. Наоборот, мембрана имеет некоторую проводимость для ионов, которые проходят через ионные каналы. Из-за препятствия свободному перемещению ионов снаружи и внутри клетки находятся одинаковые ионы, но в разных концентрациях. Существует два принципиальных механизма перемещения веществ через мембрану - посредством простой диффузии (рис. 1.11) и при по-
А - простая диффузия. Б - облегченная диффузия. В - активный транспорт: 1- мембрана мощи специфических переносчиков, встроенных в мембрану и представляющих трансмембранные интегральные белки. К последнему механизму относятся облегченная диффузия и активный ионный транспорт, который может быть первично-активным и вторично-активным. Посредством простой диффузии (без помощи переносчика) может осуществляться транспорт водонерастворимых органических соединений и газов (кислорода и углекислого газа) через липидный бислой путем их растворения в липидах клеточной мембраны; ионов Na+, Ca2+, К+, Cl- через ионные каналы клеточной мембраны, соединяющие цитоплазму клеток с внешней средой (пассивный ионный транспорт, который определяется электрохимическим градиентом и направлен от большего электрохимического потенциала к меньшему: внутрь клетки для ионов Na+, Ca2+, Cl-, наружу - для ионов К+); молекул воды через мембрану (осмос). При помощи специфических переносчиков осуществляется энергетически независимая облегченная диффузия ряда соединений (см. рис. 1.11). Ярким примером облегченной диффузии является транспорт глюкозы через мембрану нейрона. Без специализированного астроцитарного переносчика поступление глюкозы в нейроны было бы практически невозможным, так как она является относительно большой полярной молекулой. Вследствие ее быстрого превращения в глюкозу-6- фосфат внутриклеточный уровень глюкозы более низкий, чем внеклеточный уровень, и, таким образом, сохраняется градиент, обеспечивающий непрерывный поток глюкозы в нейроны. Энергетически зависимый первично-активный транспорт ионов Na+, Ca2+, К+, и Н+ - это энергозависимый перенос веществ против их электрохимических градиентов (см. рис. 1.11). Благодаря ему клетки могут накапливать ионы в концентрациях, более высоких по сравнению с окружающей средой. Движение от более низкой к более высокой концентрации и поддержание установившегося градиента возможны лишь при непрерывном энергетическом обеспечении транспортного процесса. При первичном активном транспорте происходит прямое потребление АТФ. АТФ-энергетические насосы (АТФазы) транспортируют ионы против их концентрационного градиента. На основании особенностей молекулярной организации различают 3 класса - Р, V и F (рис. 1.12). Все три класса АТФаз имеют одно или несколько мест связывания с АТФ на цитозольной поверхности мембраны. К классу Р относятся Са2+-АТФаза и Na+/К+-АТФаза. Переносчики активного ионного транспорта специфичны для транспортируемого вещества и насыщаемы, т.е. их поток максимален, когда все специфические места связывания с переносимым веществом заняты. Многие градиенты электрохимического потенциала клетки, являющиеся необходимым условием для пассивного переноса ионов, появляются в результате их активного транспорта. Так, градиенты К+ и Na+ возникают в результате их активного переноса Na+/К+ - насосом (рис. 1.13). Вследствие активности Na+/К+-насоса внутри клетки ионы К+ присутствуют в большей концентрации, но они стремятся посредством диффузии перейти во внеклеточную среду по градиенту концентрации. Для сохранения равенства положительных и отрицательных зарядов внутри клетки выход во внешнюю среду ионов К+ должен быть компенсирован входом внутрь клетки ионов Na+. Поскольку мембрана в покое значительно меньше проницаема для ионов Na+, чем для ионов К+, калий должен выходить из клетки по градиенту концентраций. В результате на внешней стороне мембраны накапливается положительный заряд, а на внутренней - отрицательный. Так поддерживается потенциал покоя мембраны. Вторично-активный транспорт ряда ионов и молекул тоже использует энергию, накопленную в результате потребления АТФ и затраченную на создание градиента концентрации. Градиент концентрации ионов относительно мембраны используется как источник энергии, созданный первичным активным транспортом (рис. 1.14). Таким образом, вторично-активный транспорт включает котранспорт и противотранспорт: поток ионов от более высокой (более высокое энергетическое состояние) к более низкой (более низкое энергетическое состояние) концентрации обеспечивает энергию для перемещения активно транспортируемого вещества из области его низкой концентрации в область высокой.
Потенциалы клетки, определяемые пассивным ионным транспортом В ответ на подпороговый, близкий к порогу и пороговые импульсы электрического тока возникают соответственно пассивный электротонический потенциал, локальный ответ и потенциал действия (рис. 1.15). Все эти потенциалы определяются пассивным ионным транспортом через мембрану. Для их возникновения требуется поляризация мембраны клетки, которая может осуществляться внеклеточно (обычно наблюдается на нервных волокнах) и внутриклеточно (обычно отмечается на теле клетки). Пассивный электротонический потенциал возникает в ответ на подпороговый импульс, который не приводит к открытию ионных каналов и определяется только емкостными и резистивными свойствами мембраны клетки. Пассивный электротонический потенциал характеризуется постоянной времени, которая отражает пассивные свойства мембраны временной ход изменений мембранного потенциала, т.е. скорость, с которой он меняется при переходе от одного значения к другому. Пас-
сивному электротоническому потенциалу присуще равенство скоростей нарастания и спада экспоненты. Существует линейная зависимость между амплитудами электрического стимула и пассивного электротонического потенциала, причем увеличение длительности импульса не меняет этой закономерности. Пассивный электротонический потенциал распространяется по аксону с затуханием, которое определяется постоянной длины мембраны. При приближении силы электрического импульса к пороговой величине возникает локальный ответ мембраны, который проявляется изменением формы пассивного электротонического потенциала и развитием самостоятельного пика небольшой амплитуды, по форме напоминающего S-образную кривую (см. рис. 1.15). Первые признаки локального ответа регистрируются при действии стимулов, составляющих примерно 75% пороговой величины. При усилении раздражающего тока амплитуда локального ответа увеличивается нелинейно и может не только достигать критического потенциала, но и превышать его, не перерастая, однако, в потенциал действия. Самостоятельное развитие локального ответа связано с повышением натриевой проницаемости мембраны через натриевые каналы, обеспечивающие входящий ток, который при пороговом раздражителе вызывает фазу деполяризации потенциала действия. Однако при подпороговом стимуле это повышение проницаемости недостаточно для запуска процесса регенеративной деполяризации мембраны, поскольку открывается лишь небольшая часть натриевых каналов. Начавшаяся де-
А - Динамика изменения мембранного потенциала в зависимости от силы деполяризующего импульса электрического тока. Б - Дискретное нарастание силы деполяризующего импульса поляризация приостанавливается. В результате выхода из клетки ионов К+ потенциал возвращается на уровень потенциала покоя. В отличие от потенциала действия локальный ответ не имеет четкого порога возникновения и не подчиняется закону «все или ничего»: с увеличением силы электрического импульса амплитуда локального ответа растет. В организме локальный ответ является электрофизиологическим выражением местного возбуждения и, как правило, предшествует потенциалу действия. Иногда локальный ответ может существовать самостоятельно в виде возбуждающего постсинаптического потенциала. Примерами самостоятельного значения локального потенциала являются проведение возбуждения от амакриновых клеток сетчатки - нейронов ЦНС, лишенных аксонов, к синаптическим окончаниям, а также ответ постсинаптической мембраны химического синапса и коммуникативная передача информации между нервными клетками, генерирующими синаптические потенциалы. При пороговой величине раздражающего электрического импульса возникает потенциал действия, состоящий из фаз деполяризации и реполяризации (рис. 1.16). Потенциал действия начинается в результате смещения под действием прямоугольного импульса электрического тока потенциала покоя (например, от -90 мВ) до уровня критического потенциала (разного для клеток различных типов). В основе фазы деполяризации лежит активация всех потенциалуправляемых натриевых каналов, вслед-
Кривые проводимости ионов взаимосвязаны с кривой потенциала действия ствие чего нарастает пассивный транспорт ионов Na+ внутрь клетки и возникает смещение мембранного потенциала до 35 мВ (этот пиковый уровень различен для клеток разных типов). Превышение потенциала действия над нулевой линией называется овершутом. По достижении пика величина потенциала падает в отрицательную область, достигая потенциала покоя (фаза реполяризации). В основе реполяризации лежат инактивация потенциалуправляемых натриевых каналов и активация потенциалуправляемых калиевых каналов. Ионы К+ пассивным транспортом выходят из клетки и возникающий при этом ток ведет к смещению мембранного потенциала в отрицательную область. Фаза реполяризации завершается следовой гиперполяризацией или следовой деполяризацией - альтернативными ионными механизмами возвращения мембранного потенциала на уровень потенциала покоя (см. рис. 1.16). При первом механизме реполяризация достигает величины покоя и продолжается дальше в более отрицательную область, после чего возвращается к уровню потенциала покоя (следовая гиперполяризация); при втором - реполяризация происходит медленно и плавно переходит в потенциал покоя (следовая деполяризация). Развитие потенциала действия сопровождается фазными изменениями возбудимости клетки - от повышенной возбудимости к абсолютной и относительной рефрактерности. Биоэлектрическая активность нейронов Первый тип биоэлектрической активности клеток присущ молчащим нейронам, не способным самостоятельно генерировать потенциалы действия. Потенциал покоя этих клеток не меняется (рис. 1.17). Нейроны второго типа способны самостоятельно генерировать потенциалы действия. Среди них выделяют клетки, генерирующие регулярную и нерегулярную ритмическую или пачечную (пачку составляет несколько потенциалов действия, после чего наблюдается короткий период покоя) активность. К третьему типу биоэлектрической активности относятся нейроны, способные самостоятельно генерировать флюктуации потенциала покоя синусоидальной или пилообразной формы, не достигающие критического потенциала. Только редкие осцилляции могут достигнуть порога и вызвать генерацию одиночных потенциалов действия. Эти нейроны получили название пейсмекерных (рис. 1.17). «Поведение» отдельных нейронов и межнейрональные взаимодействия находятся под влиянием длительной поляризации (деполяризации или гиперполяризации) постсинаптических мембран клеток. Стимуляция нейронов постоянным деполяризующим электрическим током вызывает ответы ритмическими разрядами потенциалов действия. После прекращения длительной деполяризации мембраны наступает постактивационное торможение, при котором клетка не способна генерировать потенциалы действия. Длительность стадии постактивационного торможения прямо коррелирует с амплитудой стимулирующего тока. Затем клетка постепенно восстанавливает привычный ритм генерации потенциалов. Напротив, постоянный гиперполяризующий ток тормозит развитие потенциала действия, что имеет особое значение по отношению к нейронам, обладающим спонтанной активностью. Нарастание гиперполяризации мембраны клетки приводит к уменьшению частоты спайковой активности и увеличению амплитуды каждого потенциала действия; следующей стадией является полное прекращение генерации потенциалов. После прекращения длительной гиперполяризации мембраны наступает фаза посттормозной активации, когда клетка начинает самопроизвольно генерировать с более высокой частотой, чем обычно, потенциалы действия. Длительность стадии постактивационной активации прямо коррелирует с амплитудой гиперполяризующего тока, после чего клетка постепенно восстанавливает привычный ритм генерации потенциалов.
1.4. Проведение возбуждения по нервному волокну Закономерности проведения возбуждения по нервным волокнам определяются как электрическими, так и морфологическими особенностями аксонов. Нервные стволы состоят из миелинизированных и немиелинизированных волокон. Мембрана немиелинизированного нервного волокна прямо контактирует с внешней средой, т.е. обмен ионами между внутриклеточной и внеклеточной средой может происходить в любой точке немиелинизированного волокна. Миелинизированное нервное волокно на большем протяжении покрыто жировой (миелиновой) оболочкой, выполняющей функции изолятора (см. рис. 1.18). Миелин от одной глиальной клетки формирует регион миелинизированного нервного волокна, отделенный от следующего региона, образованного другой глиальной клеткой, немиелинизированным участком - перехватом Ранвье (рис. 1.19). Длина перехвата Ранвье составляет всего 2 мкм, а длина миелинизированного участка волокна между соседними перехватами Ранвье достигает 2000 мкм. Перехваты Ранвье полностью свободны от миелина и могут контактировать с внеклеточной жидкостью, т.е. электрическая активность миелинизированного нервного волокна ограничена мембраной перехватов Ранвье, через которую способны проникать ионы. В этих участках мембраны отмечается наибольшая плотность потенциалуправляемых натриевых каналов. Пассивный электротонический потенциал распространяется по нервному волокну на небольшие расстояния (рис. 1.20), при этом его ампли-
1 - плазматическая мембрана аксона; 2 - миелиновые мембраны; 3 - цитозоль шванновской клетки; 4 - зона перехвата Ранвье; 5 - плазматическая мембрана шванновской клетки туда, скорость нарастания и падения с расстоянием уменьшаются (феномен затухания возбуждения). Распространение возбуждения в форме потенциала действия не сопровождается изменением формы или амплитуды потенциала, так как при пороговой деполяризации активизируются потенциалуправляемые ионные каналы, чего не происходит при распространении пассивного электротонического потенциала. Процесс распространения потенциала действия зависит от пассивных (емкость, сопротивление) и активных (активация потенциалуправляемых каналов) свойств мембраны нервного волокна. И внутренняя, и внешняя среда аксона является хорошим проводником. Мембрана аксона, несмотря на изолирующие свойства, также может проводить ток из-за наличия каналов «утечки» ионов. При раздражении немиелинизированного волокна в месте раздражения открываются потенциалуправляемые натриевые каналы, что вызывает возникновение входящего тока и генерацию фазы деполяризации потенциала действия на этом участке аксона. Входящий ток Na+ индуцирует локальные круги тока между деполяризованной и недеполяризованной областями мембраны. Благодаря описанному механизму в немиелинизированном волокне потенциал действия распространяется в обе стороны от места возбуждения. В миелинизированном нервном волокне потенциалы действия генерируются только в перехватах Ранвье. Электрическое сопротивление участков, покрытых миелиновой оболочкой, высокое и не допускает развития локальных круговых токов, которые необходимы для генерации потенциала действия. При распространении возбуждения по миелинизированному волокну нервный импульс перескакивает с одного перехвата Ранвье на другой (сальтаторное проведение) (см. рис. 1.20). При этом потенциал действия может распространяться в обе стороны от места раздражения, как и в немиелинизированном волокне. Сальтаторное проведе-
A - Распространение потенциала действия по немиелинизированному аксону: а - аксон в покое; б - инициация потенциала действия и возникновение локальных токов; в - распространение локальных токов; г - распространение потенциала действия по аксону. Б - Распространение потенциала действия от тела нейрона к терминальному окончанию. B - Сальтаторное проведение импульса по миелинизированному волокну. Перехваты Ранвье разделяют сегменты миелиновой оболочки аксона ние импульса обеспечивает в 5-50 раз более высокую скорость проведения возбуждения по сравнению с немиелинизированным волокном. Кроме того, оно более экономично, так как локальная деполяризация мембраны аксона только в месте перехвата Ранвье приводит к потере в 100 раз меньшего количества ионов, чем при формировании локальных токов в немиелинизированном волокне. Кроме того, при сальтаторном проведении минимально задействованы потенциалуправляемые калиевые каналы, вследствие чего у потенциалов действия миелинизированных волокон часто нет фазы следовой гиперполяризации. Законы проведения возбуждения по нервному волокну Первый закон: при раздражении нервного волокна возбуждение по нерву распространяется в обе стороны. Второй закон: распространение возбуждения в обе стороны происходит с одинаковой скоростью. Третий закон: возбуждение по нерву распространяется без феномена затухания, или без декремента. Четвертый закон: проведение возбуждения по нервному волокну возможно лишь при его анатомической и физиологической целостности. Любая травма поверхностной мембраны нервного волокна (перерезка, сдавление вследствие воспаления и отека окружающих тканей) нарушает проведение раздражения. Проведение нарушается и при изменении физиологического состояния волокна: блокаде ионных каналов, охлаждении и т.д. Пятый закон: возбуждение распространения по нервным волокнам изолировано, т.е. не переходит с одного волокна на другое, а возбуждает лишь те клетки, с которыми контактируют окончания данного нервного волокна. В связи с тем, что в состав периферического нерва обычно входит множество разных волокон (двигательных, чувствительных, вегетативных), иннервирующих разные органы и ткани и выполняющих разные функции, изолированное проведение по каждому волокну имеет особое значение. Шестой закон: нервное волокно не утомляется; потенциал действия волокна имеет одинаковую амплитуду очень длительное время. Седьмой закон: скорость проведения возбуждения различна в разных нервных волокнах и определяется электрическим сопротивлением внутри- и внеклеточной среды, мембраны аксона, а также диаметром нервного волокна. С увеличением диаметра волокна скорость проведения раздражения возрастает. Классификация нервных волокон На основании скорости проведения возбуждения по нервным волокнам, длительности фаз потенциала действия и особенностей строения выделяют три основных типа нервных волокон: А, В и С. Все волокна типа А миелинизированные; их разделяют на 4 подгруппы: α, β, γ и δ. Наибольший диаметр у αА-волокон (12-22 мкм), что определяет высокую скорость проведения возбуждения по ним (70- 170 м/с). Волокна типа αА у человека проводят возбуждение от двигательных нейронов передних рогов спинного мозга к скелетным мышцам, а также от проприоцептивных рецепторов мышц к чувствительным центрам ЦНС. Другие волокна типа А (β, γ и δ) имеют меньший диаметр, меньшую скорость проведения возбуждения и более длительный потенциал действия. К этим группам волокон относятся преимущественно чувствительные волокна, проводящие импульсы от различных рецепторов в ЦНС; исключение составляют волокна γА, которые проводят возбуждение от γ-нейронов передних рогов спинного мозга к интрафузальным мышечным волокнам. Волокна типа В также миелинизированные, относящиеся преимущественно к преганглионарным волокнам вегетативной нервной системы. Скорость проведения по ним составляет 3-18 м/с, длительность потенциала действия почти в 3 раза превышает аналогичный показатель волокон типа А. Для этих волокон не характерна фаза следовой деполяризации. Волокна типа С немиелинизированные, имеют малый диаметр (около 1 мкм) и низкую скорость проведения возбуждения (до 3 м/с). Большинство волокон типа С являются постганглионарными волокнами симпатической нервной системы, некоторые волокна типа С участвуют в проведении возбуждения от болевых, температурных и других рецепторов. 1.5. Кодирование Информация, передаваемая по аксону тем или иным способом, кодируется. Совокупность нейронов, обеспечивающих определенную функцию (например, конкретную сенсорную модальность), формирует проекционный путь (первый способ кодирования). Так, зрительный путь включает в себя нейроны сетчатки, латеральное коленчатое тело таламуса и зрительные области коры больших полушарий. Аксоны, проводящие зрительные сигналы, входят в состав зрительного нерва, зрительного тракта, зрительной лучистости. Физиологическим стимулом для активации зрительной системы служит свет, попадающий на сетчатку. Нейроны сетчатки преобразуют эту информацию и передают сигнал далее по зрительному пути. Однако при механическом или электрическом раздражении нейронов зрительного пути тоже возникает зрительное ощущение, хотя, как правило, искаженное. Итак, нейроны зрительной системы составляют проекционный путь, при активации которого возникает зрительное ощущение. Двигательные пути также представляют проекционные структуры. Например, при активации определенных нейронов коры больших полушарий генерируются разряды в мотонейронах мышц кисти, и эти мышцы сокращаются. Второй способ кодирования обусловлен принципом упорядоченной пространственной (соматотопической) организации ЦНС. Соматотопические карты составлены определенными группами нейронов сенсорной и двигательной систем. Эти группы нейронов, во-первых, получают информацию от соответствующим образом локализованных областей поверхности тела и, во-вторых, посылают двигательные команды к определенным частям тела. В зрительной системе участки сетчатки представлены в коре мозга группами нейронов, образующими ретинотопические карты. В слуховой системе частотные характеристики звуков отражены в тонотопичсских картах. Третий способ кодирования информации основан на варьировании характеристик последовательностей (серий) нервных импульсов, направ- ляемых в результате синаптической передачи к следующей группе нейронов, при этом кодирующий механизм - временная организация разряда нервных импульсов. Возможны разные виды такого кодирования. Часто кодом служит средняя частота разряда: во многих сенсорных системах увеличение интенсивности стимула сопровождается повышением частоты разряда сенсорных нейронов. Кроме того, кодом могут служить длительность разряда, разнообразное группирование импульсов в разряде, продолжительность высокочастотных залпов импульсов и т.д. 1.6. Проведение возбуждения между клетками. Синапсы Взаимосвязи между нервными клетками осуществляются межнейрональными контактами, или синапсами. Информация в виде серии потенциалов действия поступает от первого (пресинаптического) нейрона на второй (постсинаптический) либо путем формирования локального тока между соседними клетками (электрические синапсы), либо опосредованно химическими веществами - медиаторами, нейротрансмиттерами (химические синапсы), либо с помощью обоих механизмов (смешанные синапсы). Быстрая передача сигналов осуществляется электрическими синапсами, более медленная - химическими. Типичные синапсы - это образования, сформированные терминалями аксона одного нейрона и дендритами другого (аксодендритические синапсы). Кроме того, существуют аксосоматические, аксо-аксональные и дендродендритические синапсы (рис. 1.21). Некоторые ассоциативные нейроны имеют разнообразные синаптические связи (рис. 1.22). Синапс между аксоном мотонейрона и волокном скелетной мышцы называется двигательной концевой пластинкой, или нервномышечным соединением. У электрического синапса (рис. 1.23) клеточные мембраны соседних нейронов тесно прилегают друг к другу, щель между ними составляет около 2 нм. Участки мембран соседних клеток, образующие щелевой контакт, содержат специфические белковые комплексы, состоящие из 6 субъединиц (коннексонов), располагающихся в таком порядке, что в центре контакта образуют заполненную водой пору. Коннексоны мембран соседних клеток, выстраиваясь друг против друга, образуют открытую связь - «каналы», расстояние между которыми составляет около 8 нм.
А - а - электрический синапс; б - шипиковый синапс, содержащий электронно-плотные везикулы; в - «en passant»-синапс, или синаптическая «почка»; г - тормозной синапс, расположенный на начальной части аксона (содержит эллипсоидные везикулы); д - дендритный шипик; е - шипиковый синапс; ж - тормозный синапс; з - аксо-аксональный синапс; и - реципрокный синапс; к - возбуждающий синапс. Б - Нетипичные синапсы: 1 - аксо-аксональный синапс. Окончание одного аксона может регулировать активность другого; 2 - дендродендрический синапс; 3 - сомасоматический синапс Электрические синапсы наиболее часто образуются в эмбриональной стадии развития, у взрослого их количество уменьшается. Однако и во взрослом организме значимость электрических синапсов сохраняется для клеток глии и амакринных клеток сетчатки глаза; электрические синапсы можно обнаружить в стволе головного мозга, особенно в нижних оливах, в сетчатке глаза, вестибулярных корешках. Деполяризация пресинаптической мембраны приводит к образованию разности потенциалов с недеполяризованной постсинаптической мембраной. В результате через каналы, образованные коннексонами, начинается движение положительных ионов по градиенту разности потенциалов в постсинаптическую клетку или движение анионов в обратном направлении. При достижении на постсинаптической мембране
1 - аксонный холмик, переходящий в аксон; 2 - миелиновая оболочка; 3 - аксодендритический синапс; 4 - ядро; 5 - дендрит; 6 - аксосоматический синапс
А - Щелевой контакт между участками мембран соседних клеток. Б - Коннексоны мембран соседних клеток образуют межнейрональный «канал». 1 - протеиновый комплекс; 2 - ионный канал. 3 - канал; 4 - коннексон клетки 1; 5 - каждые шесть субъединиц; 6 - коннексон клетки 2 суммарной деполяризации пороговой величины возникает потенциал действия. Важно отметить, что в электрическом синапсе ионные токи возникают с минимальной временной задержкой, составляющей 10-5 с, что объясняет высокую синхронизацию ответа даже очень большого числа клеток, соединенных щелевым контактом. Проведение тока через электрический синапс также возможно в обоих направлениях (в противоположность химическому синапсу). Функциональное состояние электрических синапсов регулируется ионами Са2+ и уровнем мембранного потенциала клеток, что создает условия для влияния на распространение возбуждения вплоть до его прекращения. К особенностям деятельности электрических синапсов следует отнести невозможность прямого переноса возбуждения на отдаленные клетки, так как с возбужденной клеткой непосредственно связаны лишь немногие другие; уровень возбуждения в пресинаптической и постсинаптической клетках одинаков; затормозить распространение возбуждения невозможно, в связи с этим головной мозг новорожденных и детей раннего возраста, содержащий значительно больше электрических синапсов, чем мозг взрослого, оказывается значительно более возбудимым для электрических процессов: быстро распространяющееся электрическое возбуждение не подвергается тормозной коррекции и почти мгновенно становится генерализованным, что объясняет его особую уязвимость и подверженность развитию пароксизмальной активности. Следует отметить, что при некоторых формах демиелинизирующих полиневропатий аксоны, входящие в состав одного нервного ствола, начинают тесно соприкасаться друг с другом, формируя патологические зоны (эфапсы), внутри которых становится возможным «перепрыгивание» потенциала действия с одного аксона на другой. В результате возможно появление симптоматики, отражающей поступление «псевдоинформации» в головной мозг, - ощущение боли без раздражения периферических болевых рецепторов и т.д. Химический синапс также передает электрический сигнал от пресинаптической к постсинаптической клетке, но в нем ионные каналы на постсинаптической мембране открываются или закрываются при помощи химических веществ-переносчиков (медиаторов, нейротрансмиттеров), высвобождающихся из пресинаптической мембраны (рис. 1.24). Изменение возможности проведения определенных ионов через постсинаптическую мембрану является основой функционирования химических синапсов. Ионные токи изменяют потенциал постсинаптической мембраны, т.е. вызывают развитие постсинаптического потенциала. В зависимости от того, проводимость каких ионов изменяется при действии нейротрансмиттера, его эффект может быть тормозящим (гиперполяризация постсинаптической мембраны вследствие дополнительного выходящего тока ионов К+ или входящего тока ионов С1-) или возбуждающим (деполяризация постсинаптической мембраны при дополнительном входящем токе ионов Ca2+ или Na+). В синапсе (рис. 1.25) выделяют пресинаптический отросток, содержащий пресинаптические пузырьки (везикулы), и постсинаптическую часть (дендрит, тело клетки или аксон). В пресинаптическом нервном окончании в везикулах аккумулируются нейротрансмиттеры. Синаптические везикулы фиксируются в основном на цитоскелете посредством белков синапсина, локализованного на цитоплазматической поверхности каждой везикулы, и спектрина, расположенного на волокнах F- актина цитоскелета (рис. 1.26). Меньшая часть везикул связана с преси- наптической мембраной посредством белка везикулы синаптобревина и белка пресинаптической мембраны синтаксина. Одна везикула содержит 6000-8000 молекул трансмиттера, что составляет 1 квант трансмиттера, т.е. минимальное количество, освобождающееся в синаптическую щель. Когда серия потенциалов действия достигает нервного окончания (пресинаптической мембраны), ионы Са2+ устремляются внутрь клетки. На связанных с пресинаптической мембраной везикулах ионы Са2+ связываются с белком везикул синаптотагми-
Дата добавления: 2015-12-16 | Просмотры: 1829 | Нарушение авторских прав |
 Рис. 1.1. Общее строение нервной системы.
Рис. 1.1. Общее строение нервной системы. Рис. 1.2. Эмбриональное развитие нервной системы млекопитающего. Схема развития нервной рубки на стадии трех (А) и пяти (Б) мозговых пузырей. А. I - Общий вид сбоку: 1 - краниальный изгиб; 2 - шейный изгиб; 3 - спинномозговой узел. II - Вид сверху: 4 - передний мозг; 5 - средний мозг; 6 - ромбовидный мозг; 7 - невроцель; 8 - стенка нервной трубки; 9 - зачаточный спинной мозг.
Рис. 1.2. Эмбриональное развитие нервной системы млекопитающего. Схема развития нервной рубки на стадии трех (А) и пяти (Б) мозговых пузырей. А. I - Общий вид сбоку: 1 - краниальный изгиб; 2 - шейный изгиб; 3 - спинномозговой узел. II - Вид сверху: 4 - передний мозг; 5 - средний мозг; 6 - ромбовидный мозг; 7 - невроцель; 8 - стенка нервной трубки; 9 - зачаточный спинной мозг. Рис. 1.3. Строение нейрона. А - Типичный нейрон, состоящий из собственно тела, дендритов и аксона: 1 - начало аксона; 2 - дендриты; 3 - тело нейрона; 4 - аксон; 5 - шванновская клетка; 6 - разветвление аксона. Б - Увеличенное тело нейрона. Аксональный холмик не содержит субстанции Ниссля: 7 - ядро; 8 - аппарат Гольджи; 9 - митохондрии; 10 - аксональный холмик; 11 - субстанция Ниссля
Рис. 1.3. Строение нейрона. А - Типичный нейрон, состоящий из собственно тела, дендритов и аксона: 1 - начало аксона; 2 - дендриты; 3 - тело нейрона; 4 - аксон; 5 - шванновская клетка; 6 - разветвление аксона. Б - Увеличенное тело нейрона. Аксональный холмик не содержит субстанции Ниссля: 7 - ядро; 8 - аппарат Гольджи; 9 - митохондрии; 10 - аксональный холмик; 11 - субстанция Ниссля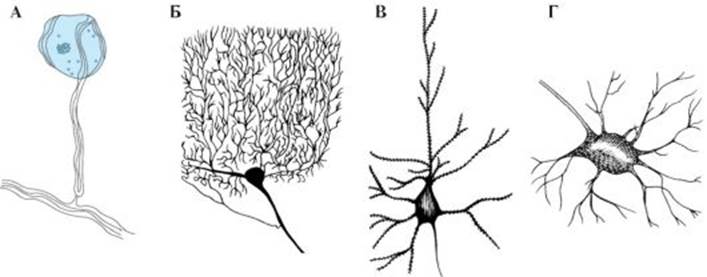 Рис. 1.4. Варианты нейронов разной формы.
Рис. 1.4. Варианты нейронов разной формы. Рис. 1.5. Ультраструктура нервной клетки.
Рис. 1.5. Ультраструктура нервной клетки.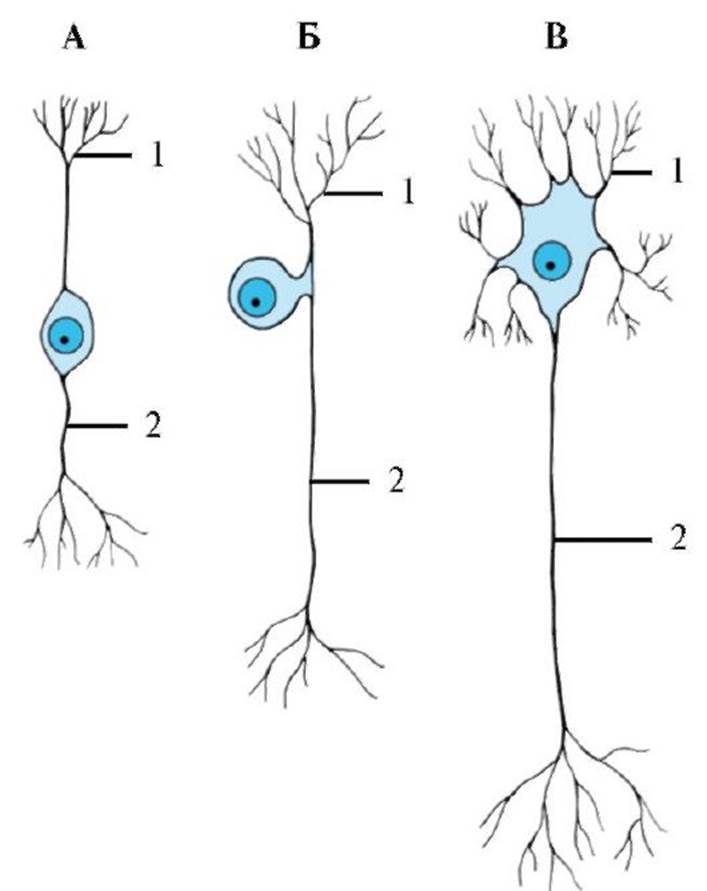 Рис. 1.6. Классификация нейронов по количеству отростков, отходящих от тела клетки.
Рис. 1.6. Классификация нейронов по количеству отростков, отходящих от тела клетки.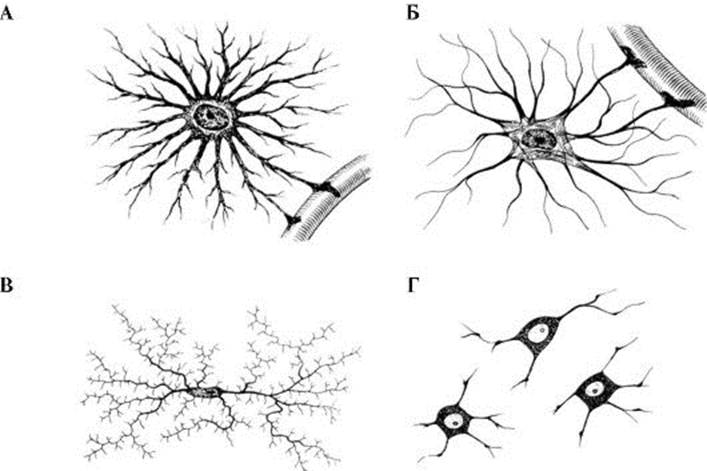 Рис. 1.7. Основные типы глиальных клеток.
Рис. 1.7. Основные типы глиальных клеток.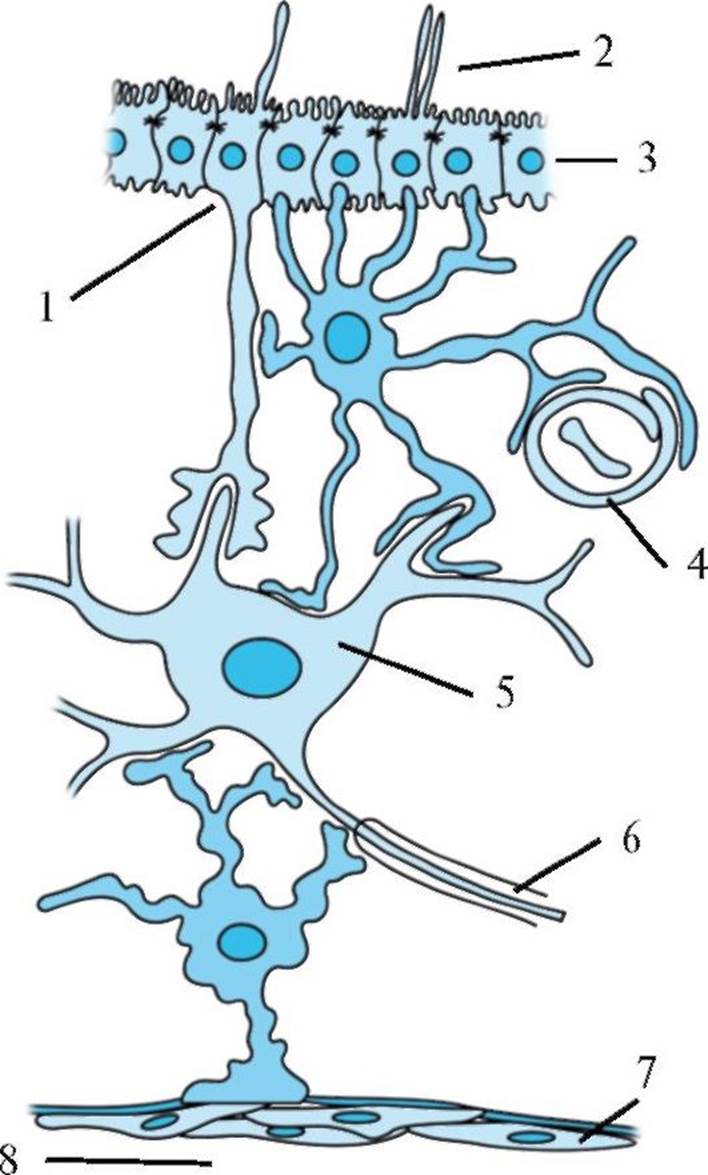 Рис. 1.8. Взаимодействие астроцитов с окружающими клеточными элементами.
Рис. 1.8. Взаимодействие астроцитов с окружающими клеточными элементами. Рис. 1.9. Олигодендроцит: формирование миелиновой оболочки аксона. 1 - аксон; 2 - миелин; 3 - гладкий эндоплазматический ретикулум; 4 - нейрофиламенты; 5 - митохондрии
Рис. 1.9. Олигодендроцит: формирование миелиновой оболочки аксона. 1 - аксон; 2 - миелин; 3 - гладкий эндоплазматический ретикулум; 4 - нейрофиламенты; 5 - митохондрии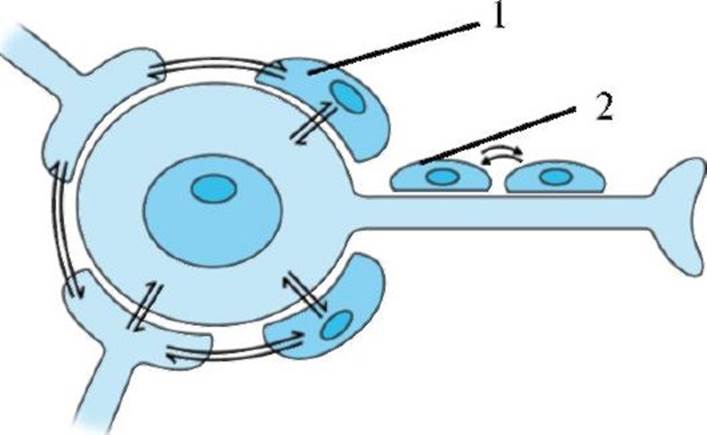 Рис. 1.10. Взаимодействие глиальных клеток и нейронов. Схематично изображено стрелками. 1 - сателлитная глиальная клетка; 2 - глиальная клетка, синтезирующая миелин
Рис. 1.10. Взаимодействие глиальных клеток и нейронов. Схематично изображено стрелками. 1 - сателлитная глиальная клетка; 2 - глиальная клетка, синтезирующая миелин Рис. 1.11. Транспорт веществ через клеточную мембрану.
Рис. 1.11. Транспорт веществ через клеточную мембрану.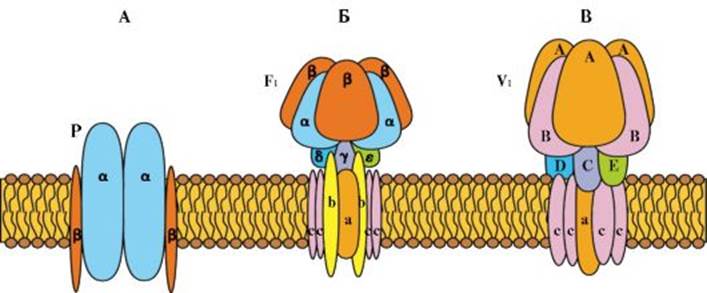 Рис. 1.12. Три класса АТФ-зависимых ионных насосов. А - P-класс. Б - F1-класс В - V1-класс
Рис. 1.12. Три класса АТФ-зависимых ионных насосов. А - P-класс. Б - F1-класс В - V1-класс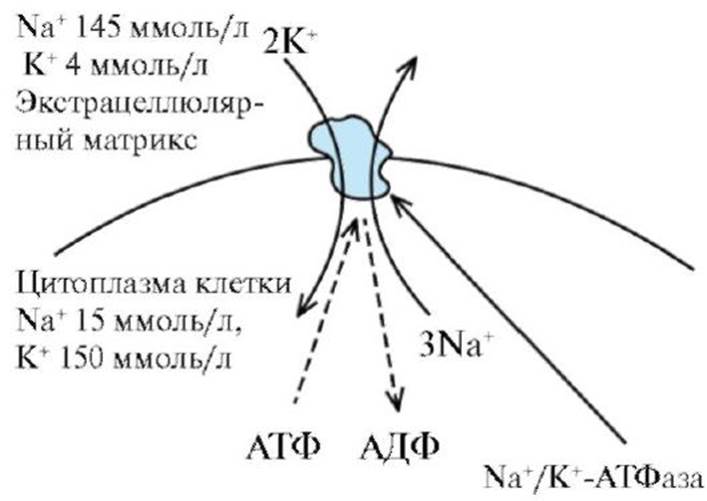 Рис. 1.13. Механизм работы Na+/K+ насоса
Рис. 1.13. Механизм работы Na+/K+ насоса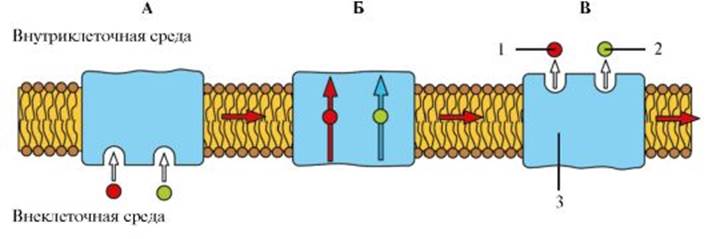 Рис. 1.14. Механизм работы вторично-активного транспорта. А - Этап 1. Б - Этап 2. В - Этап 3: 1 - Na+; 2 - молекула вещества, которая должна быть перенесена против градиента концентрации; 3 - транспортер. При связывании Na+ с переносчиком возникают аллостерические изменения в связывающем центре белка-переносчика для молекулы переносимого вещества, что вызывает конформационные изменения белка-переносчика, позволяющие ионам Na+ и связанному веществу выйти на другой стороне мембраны
Рис. 1.14. Механизм работы вторично-активного транспорта. А - Этап 1. Б - Этап 2. В - Этап 3: 1 - Na+; 2 - молекула вещества, которая должна быть перенесена против градиента концентрации; 3 - транспортер. При связывании Na+ с переносчиком возникают аллостерические изменения в связывающем центре белка-переносчика для молекулы переносимого вещества, что вызывает конформационные изменения белка-переносчика, позволяющие ионам Na+ и связанному веществу выйти на другой стороне мембраны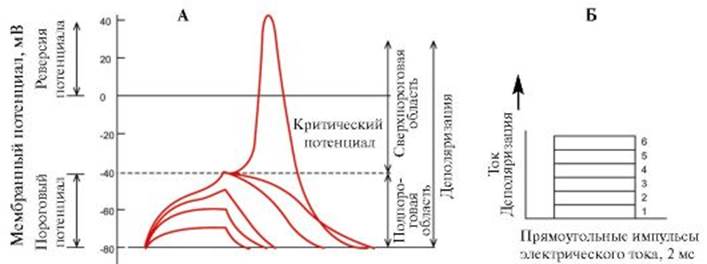 Рис. 1.15. Потенциалы мембраны клетки.
Рис. 1.15. Потенциалы мембраны клетки.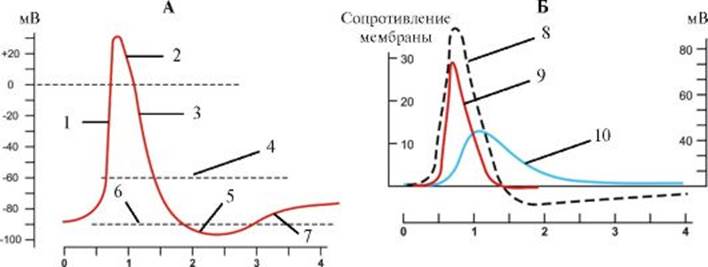 Рис. 1.16. Изменения мембранного потенциала нейрона (А) и проводимости ионов через плазмолемму (Б) при возникновении потенциала действия. 1 - быстрая деполяризация; 2 - овершут; 3 - реполяризация; 4 - пороговый потенциал; 5 - гиперполяризация; 6 - потенциал покоя; 7 - медленная деполяризация; 8 - потенциал действия; 9 - проницаемость для ионов натрия; 10 - проницаемость для ионов калия.
Рис. 1.16. Изменения мембранного потенциала нейрона (А) и проводимости ионов через плазмолемму (Б) при возникновении потенциала действия. 1 - быстрая деполяризация; 2 - овершут; 3 - реполяризация; 4 - пороговый потенциал; 5 - гиперполяризация; 6 - потенциал покоя; 7 - медленная деполяризация; 8 - потенциал действия; 9 - проницаемость для ионов натрия; 10 - проницаемость для ионов калия.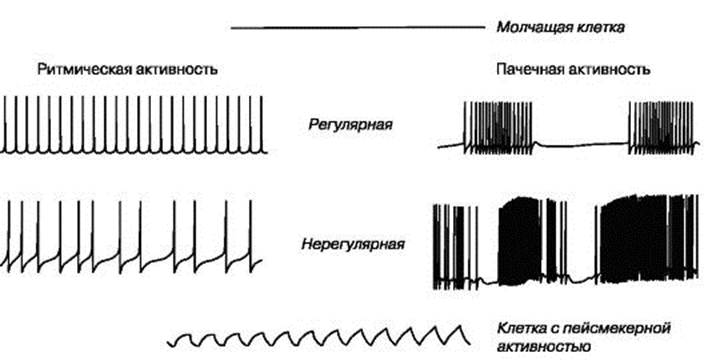 Рис. 1.17. Типы биоэлектрической активности нервных клеток
Рис. 1.17. Типы биоэлектрической активности нервных клеток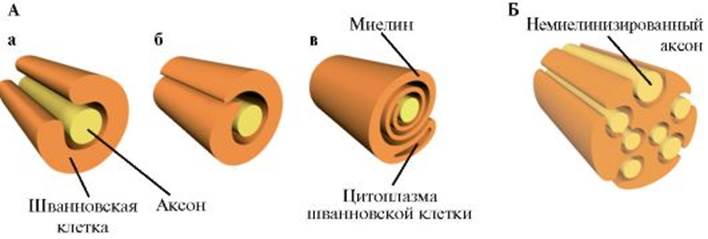 Рис. 1.18. Схема миелинизации периферического нервного волокна. А - Этапы миелинизации. а - аксон обхватывается отростком шванновской клетки; б - отросток шванновской клетки накручивается вокруг аксона; в - шванновская клетка теряет большую часть цитоплазмы, превращаясь в пластинчатую оболочку вокруг аксона. Б - Немиелинизированные аксоны, окруженные отростком шванновской клетки
Рис. 1.18. Схема миелинизации периферического нервного волокна. А - Этапы миелинизации. а - аксон обхватывается отростком шванновской клетки; б - отросток шванновской клетки накручивается вокруг аксона; в - шванновская клетка теряет большую часть цитоплазмы, превращаясь в пластинчатую оболочку вокруг аксона. Б - Немиелинизированные аксоны, окруженные отростком шванновской клетки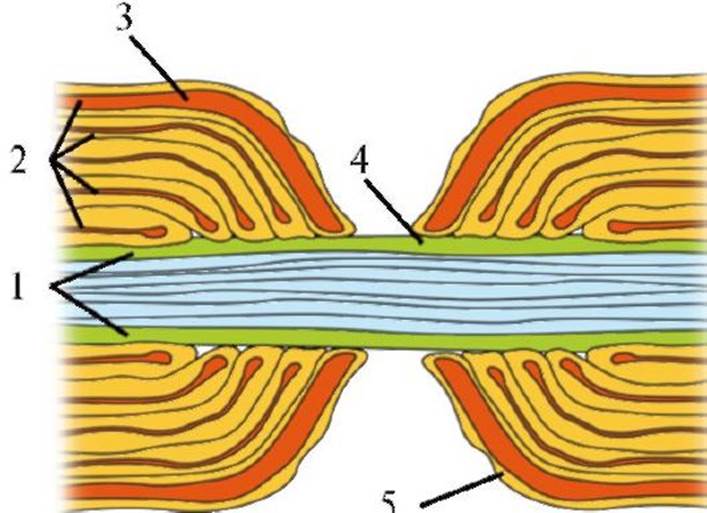 Рис. 1.19. Структура перехвата Ранвье.
Рис. 1.19. Структура перехвата Ранвье. Рис. 1.20. Схема распространения электрического потенциала по нервному волокну.
Рис. 1.20. Схема распространения электрического потенциала по нервному волокну. Рис. 1.21. Основные виды синапсов.
Рис. 1.21. Основные виды синапсов.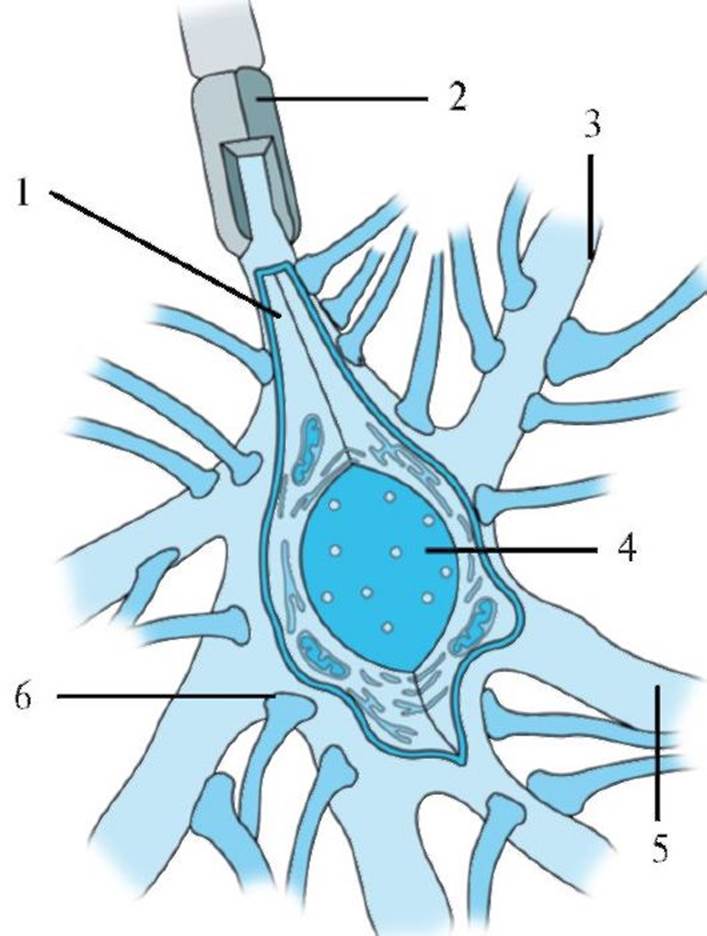 Рис. 1.22. Ассоциативный нейрон со множественными синаптическими связями.
Рис. 1.22. Ассоциативный нейрон со множественными синаптическими связями.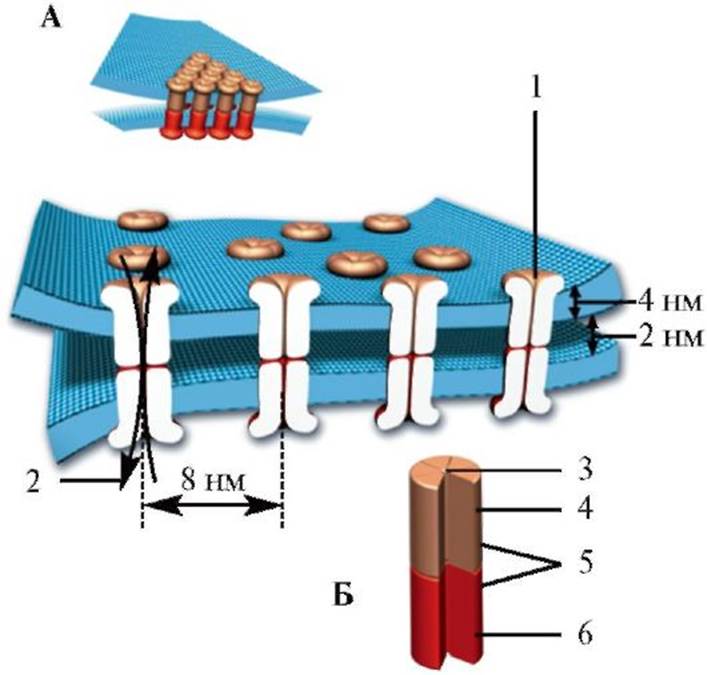 Рис. 1.23. Строение электрического синапса.
Рис. 1.23. Строение электрического синапса. Рис. 1.24. Основные этапы передачи через химический синапс: 1 - потенциал действия достигает пресинаптического окончания; 2 - деполяризация пресинаптической мембраны приводит к открытию потенциалзависимых Са2+-каналов; 3 - ионы Са2+ опосредуют слияние везикул с пресинаптической мембраной; 4 - молекулы медиатора высвобождаются в синаптическую щель путем экзоцитоза; 5 - молекулы медиатора связываются с постсинаптическими рецепторами, активируя ионные каналы; 6 - происходит изменение проводимости мембраны для ионов и в зависимости от свойств медиатора возникает возбуждающий (деполяризация) или тормозной (гиперполяизация) потенциал постсинаптической мембраны; 7 - ток ионов распространяется по постсинаптической мембране; 8 - медиаторные молекулы возвращаются в пресинаптическое окончание путем обратного захвата или 9 - диффундируют во внеклеточную жидкость
Рис. 1.24. Основные этапы передачи через химический синапс: 1 - потенциал действия достигает пресинаптического окончания; 2 - деполяризация пресинаптической мембраны приводит к открытию потенциалзависимых Са2+-каналов; 3 - ионы Са2+ опосредуют слияние везикул с пресинаптической мембраной; 4 - молекулы медиатора высвобождаются в синаптическую щель путем экзоцитоза; 5 - молекулы медиатора связываются с постсинаптическими рецепторами, активируя ионные каналы; 6 - происходит изменение проводимости мембраны для ионов и в зависимости от свойств медиатора возникает возбуждающий (деполяризация) или тормозной (гиперполяизация) потенциал постсинаптической мембраны; 7 - ток ионов распространяется по постсинаптической мембране; 8 - медиаторные молекулы возвращаются в пресинаптическое окончание путем обратного захвата или 9 - диффундируют во внеклеточную жидкость