|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология
|
Отношение полов
Численным отношением полов или просто отношением полов называется частное от деления числа самцов на число самок. Эта величина у большинства раздельнополых организмов близка к единице, что является прямым результатом хромосомного механизма определения пола (рис. 3.16). Самки (точнее, гомогаметный пол) продуцируют га-
| Рис. 3.16. Хромосомный механизм определения отношения численности полов. Количество самок и самцов приблизительно одинаково у тех организмов, у которых один пол гомогаметный, а другой - гетерогаметный. Гомогаметные особи продуцируют в отношении половых хромосом только один тип гамет, а гетерогаметные - два типа гамет в равном количестве.
| 
|
3. Хромосомные основы наследственности 85
меты одного типа, X, самцы - двух типов, X и Y, причем в равном количестве. В результате зиготы XX и XY также образуются в одинаковом количестве.
У пчел и родственных им насекомых отношение полов определяется долей оплодотворенных яиц; не удивительно, что оно не равно единице, а обычно много меньше. У тех видов, у которых пол определяется внешними условиями, отношение полов обычно отлично от единицы, причем часто, как, например, у L. dimidiatus, бывает значительно меньшим единицы.
На отношение полов могут влиять некоторые гены. Например, рецессивные летальные гены (т.е. гены, приводящие к гибели организма до достижения половозрелости) в Х-хромосоме дрозофил убивают гемизиготных самцов, но оставляют невредимыми гомозиготных самок. У Drosophila pseudoobscura существует линия, в которой потомство состоит почти исключительно из самок, вероятно, вследствие того, что содержащие Y-хромосому гаметы самцов либо не возникают вовсе, либо не способны оплодотворять яйцеклетки.
Литература
| Bridges C.B. (1916). Nondisjunction as proof of the chromosome theory of heredity, Genetics, 1, 1-52, 107-163.
Bridges C.B. (1925). Sex in relation to chromosomes and genes, Amer. Nat., 59, 127-137.
Galan F. (1964). Teoria genética del sexo zigotico en el caso de Ecballium elaterium, Revista de Biologia, 4, 187-220.
Miller O.J., Siniscalco M. (1982). Report of the Committee on the Genetic Constitution of the X and Y Chromosome, Cytogenet. Cell Genet., 32, 179-190.
Morgan L. V. (1922). Non criss-cross inheritance in
| Drosophila melanogaster, Biol. Bull., 42, 267-274.
Morgan Т.Н. (1910). Sex-limited inheritance in Drosophila, Science, 32, 120-122.
Morreale S.J., Ruiz G.J., Spotila J.R., Standora E. A. (1982). Temperature-dependent sex determination, Science, 216, 1245-1247.
Sutton W.S. (1903). The chromosomes in heredity, Biol. Bull., 4, 231-251.
Voeller В., 1968. The Chromosome Theory of Inheritance, Appleton-Century-Crofts, New York.
Zulueta A. de (1925). La herencia ligada al sexo en el coleoptero Phytodecta variabilis (Ol.), EOS, Revista Espanola de Entomologia. 1, 203-209.
| Ключевые слова и понятия
| Гапло-диплоидия
Гемизиготность
Гермафродитизм
Голандрическое наследование
| Нерасхождение первичное и вторичное
Половые хромосомы
Сцепленное с полом наследование
Хромосомная теория наследственности
|
86 Организация и передача генетического материала
Задачи
| 3.1. У супругов, зрение каждого из которых нормально, четверо детей: две дочери и два сына. У первой дочери зрение нормальное; у нее есть три сына, два из которых дальтоники. У второй дочери, а также у ее пяти сыновей зрение нормальное. Первый сын дальтоник; у него две дочери и два сына, и все видят нормально. Второй сын и четверо его сыновей также имеют нормальное зрение. Каковы генотипы дедушки с бабушкой, всех их детей, их супругов и внуков?
3.2. Если женщина, отец которой страдал гемофилией, вышла замуж за здорового мужчину, то какова вероятность того, что у ее ребенка будет гемофилия? Предположим теперь, что отец мужа также был болен гемофилией; какова вероятность в этом случае?
3.3. У дрозофилы ген редуцированных крыльев (vg) рецессивен и расположен в аутосоме; ген желтой окраски тела (у) также рецессивен, но сцеплен с полом. Если гомозиготную по этим генам самку скрестить с нормальным самцом, то как будет выглядеть потомство в F1 и F2?
3.4. Предположим, что у желтой самки с редуцированными крыльями из условия предыдущей задачи Х-хромосомы связаны общей центромерой. Какое потомство можно ожидать от скрещивания с нормальным самцом?
3.5. Петух гетерозиготен по сцепленной с полом рецессивной летали. Каково отношение полов в потомстве от скрещивания такого петуха с нормальными курами?
3.6. Иногда у кур яичники не развиваются или не функционируют, а вместо них развиваются семенники. У некоторых из таких «петухов» с переопределенным полом могут быть цыплята. Какого типа потомство можно ожидать от скрещивания таких петухов с нормальными курами? Каково будет отношение полов в потомстве с учетом того, что яйца типа WW не способны к развитию?
3.7. Рябую курицу скрещивали
| с гладким петухом. В F2 проводились все возможные типы скрещиваний. Каково будет потомство в F3?
3.8. Какие типы потомства и в каком отношении возникают при скрещивании гладкой курицы с гетерозиготным рябым петухом?
3.9. У Ecballium elaterium однодомные (гермафродитные) растения классифицируют как вариант elaterium, а двудомные (мужские и женские) - как вариант dioicum. Обычно используются следующие генетические обозначения: Е. elaterium elaterium обозначается  , мужские растения Е. elaterium dioicum - , мужские растения Е. elaterium dioicum -  , женские растения Ε. е. dioicum- , женские растения Ε. е. dioicum-  . Предположим, что в природе растения обычно имеют генетическую конституцию дикого типа. Каковы будут результаты скрещиваний: a) dioicum . Предположим, что в природе растения обычно имеют генетическую конституцию дикого типа. Каковы будут результаты скрещиваний: a) dioicum  x dioicum x dioicum  ; б) elaterium x x elaterium; в) elaterium x dioicum ; б) elaterium x x elaterium; в) elaterium x dioicum  ; г) dioicum ; г) dioicum 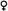 x elaterium?
3.10. Предположим, что в потомстве от скрещивания типа в) из условий предыдущей задачи плодовиты любые типы перекрестного опыления. Какие типы растений и в каких пропорциях будут присутствовать в потомстве от всех типов такого скрещивания? Ответьте на тот же вопрос применительно к результатам скрещивания типа г).
3.11. В потомстве от скрещивания двух растений Ecballium elaterium возникают исключительно гермафродитные растения. При их скрещивании в F2 одну четверть потомства составляют двудомные женские растения и три четверти - однодомные растения. Каковы генотипы представителей всех трех поколений?
3.12. Самка медоносной пчелы Apis mellifera диплоидна, а трутни гаплоидны. Существует, однако, локус со множественными аллелями, влияющий на определение пола у диплоидных особей. Все диплоидные особи, гетерозиготные по этому локусу, - самки, а гомозиготные по любому из аллелей - самцы. Гомози- x elaterium?
3.10. Предположим, что в потомстве от скрещивания типа в) из условий предыдущей задачи плодовиты любые типы перекрестного опыления. Какие типы растений и в каких пропорциях будут присутствовать в потомстве от всех типов такого скрещивания? Ответьте на тот же вопрос применительно к результатам скрещивания типа г).
3.11. В потомстве от скрещивания двух растений Ecballium elaterium возникают исключительно гермафродитные растения. При их скрещивании в F2 одну четверть потомства составляют двудомные женские растения и три четверти - однодомные растения. Каковы генотипы представителей всех трех поколений?
3.12. Самка медоносной пчелы Apis mellifera диплоидна, а трутни гаплоидны. Существует, однако, локус со множественными аллелями, влияющий на определение пола у диплоидных особей. Все диплоидные особи, гетерозиготные по этому локусу, - самки, а гомозиготные по любому из аллелей - самцы. Гомози-
|
3. Хромосомные основы наследственности 87
| готные самцы не достигают половозрелости, поскольку обычно поедаются рабочими пчелами на личиночной стадии в первые трое суток после вылупления из яичек. Таким образом, гомозиготный генотип в естественных условиях детален. Однако изъятых из улья личинок таких самцов можно довести до стадии половозрелости. В этом локусе насчитывается около двадцати аллелей, обозначаемых символами A1, А2,, А20. Пусть в потомстве некоторой оплодотворенной гаплоидным трутнем матки лишь половина яичек способна к развитию (это хорошо заметно, поскольку половина ячеек в сотах остается пустой, и пустые ячейки распределены по сотам случайным образом). Что можно сказать о генетической конституции матки и трутня?
3.13. Предположим, что диплоидный трутень оплодотворяет матку, у которой в локусе, определяющем пол, один из аллелей идентичен аллелю трутня. Какой процент потомства будет жизнеспособен в улье?
3,14. Пусть число аллелей в локусе, определяющем пол, равно двадцати. Сколько существует генетически различных типов гаплоидных самцов? Диплоидных самцов? Самок?
3.15. Локализация гена в Y-хромосоме впервые была обнаружена Антонио де Зулуэта в 1925 году. Существуют четыре фенотипических класса жуков-листоедов Phytodecta variabilis: полосатый, желтый, красный и черный, отличающиеся окраской надкрылий. Все четыре фенотипа определяются аллелями одного гена, обозначаемыми еl для полосатых жуков, еу для желтых жуков, еr для красных и еb для черных. Характер доминирования среди этих аллелей можно описать неравенством еb > еr > еy > еl, где знак «>» означает «доминантен по отношению к». Ген расположен в половых хромосомах, причем все четыре аллеля могут быть локализованы как в Х-, так и в Y-хромосоме. Зулуэта обнаружил, что полосатый фенотип очень редко встречается у самцов (0,5% всех самцов) и широко распро-
| странен у самок (59% всех самок). Как вы можете объяснить этот факт?
3.16. При скрещивании полосатой самки Phytodecta variabilis с желтым самцом в f1 обнаружено 13 полосатых самок и 11 желтых самцов. В F2 все самки (31) были полосатыми, а все самцы (29) -желтыми. Каковы генотипы родителей и потомства в F1 и F2? Используйте метод хи-квадрат для проверки вашей гипотезы.
3.17. Допустим, что вам неизвестен пол особей в поколениях F1 и F2 из скрещивания, описанного в предыдущей задаче. Могли бы такие результаты получиться, если бы ген окраски надкрылий не был сцеплен с полом? Используйте метод хиквадрат для проверки гипотезы о несцепленности с полом данного гена уже с учетом половой принадлежности особей.
3.18. При скрещивании красной самки Phytodecta variabilis с красным самцом в потомстве оказалось 15 желтых самок, 15 красных самок и 34 красных самца. При скрещивании отдельных желтых самок из поколения F1 с отдельными красными самцами из того же поколения соотношение фенотипов в потомстве оказалось в разных скрещиваниях различным: примерно в половине скрещиваний все самцы и самки в потомстве были красными, в другой половине самцы имели красную окраску, а самки примерно в равном числе были.желтыми и полосатыми. Каковы вероятные генотипы родителей?
3.19. У некоторых тропических рыб, таких как меченосцы и гуппии, в некоторых линиях гетерогаметными бывают самцы, а в других - самки. В диких линиях самки часто бывают типа XX, самцы типа XY; в некоторых аквариумных линиях самки имеют генотип ZW, самцы - ZZ. При перекрестных скрещиваниях можно получить самцов с комбинациями половых хромосом типа ZZ, XZ, XY или YY, а самок с комбинациями XX, XW, ZW или YW. Каково будет отношение полов в следующих типах скрещиваний: а)  XX x XX x  ΖΖ; б) ΖΖ; б)  ZW x ZW x  ΧΖ; в) ΧΖ; в)  XW x XW x  ΧΖ? ΧΖ?
|
4. Природа генетического
материала
Связь между менделевскими генами и хромосомами клетки была твердо доказана Бриджесом в 1916 году (гл. 3). Ему удалось установить, что все гены имеют некоторые общие свойства. Во-первых, они способны создавать собственные копии (самореплицироваться) во время удвоения хромосом в период, предшествующий мейозу. Во-вторых, в результате мутаций гены могут переходить в различные аллельные формы, что также предполагает способность к саморепликации. Редкость мутаций указывает на то, что гены представляют собой очень стабильные структуры, способные к точной дупликации. В-третьих, гены различными способами оказывают влияние на фенотип. Проявление альтернативных признаков, как впервые заметил Мендель (длинный или короткий стебель, гладкие или морщинистые семена и т. п.), служит основным критерием идентификации генов при наблюдениях над расщеплением аллелей в потомстве различающихся по данному признаку родителей. Устойчивая передача признаков из поколения в поколение, нарушаемая лишь мутациями, ставит перед нами вопросы: как определяются такие признаки? Что представляет собой образующее ген вещество, способное к саморепликации, мутациям и фенотипическому проявлению?
Ответы на такие фундаментальные вопросы неизбежно очень сложны, когда вопросы формулируются применительно к росту, развитию и размножению высших организмов, например человека, или даже менее сложных организмов, таких как мухи или растения гороха. Чем проще устроен организм, тем больше он подходит для решения подобных вопросов. Первые сведения о физических и химических основах наследственности были получены при работе с микроорганизмами.
4. Природа генетического материала 89
Бактерии, ранее изучавшиеся лишь постольку, поскольку они являются возбудителями болезней человека и домашних животных, оказались удобным объектом для исследования наследственности и природы генетического материала. Вирусы, устроенные проще, чем бактерии, оказались и более удобным объектом. Вирусы способны размножаться лишь внутри живых клеток. Бактериофаги проникают в бактериальные клетки; другие вирусы поражают клетки растений и животных, и многие из них патогенны.
Еще в 1922 г. генетик Мёллер отметил два важных общих свойства бактериофагов и генов: и те и другие способны к размножению, создавая точные копии самих себя; и те и другие в результате мутаций могут принимать новые формы. Мёллер писал1):
«С другой стороны, если тельца Д'Эрелля (бактериофаги) действительно представляют собой гены, в основных чертах сходные с генами хромосом, то это дает нам возможность подступиться к проблеме гена с совершенно новых позиций... Было бы слишком опрометчиво назвать эти тельца генами, однако мы должны признать, что какие-либо отличия между генами и ими нам не известны. Следовательно, мы не можем категорически отвергать возможность того, что когда-нибудь мы научимся растирать гены в ступке, а затем снова собирать их в пробирке. Должны ли генетики превратиться в бактериологов, химиков и физиков, одновременно оставаясь при этом зоологами и ботаниками? Хотелось бы надеяться».
В 1922 г. генетики считали, что менделевские гены могут находиться лишь в ядрах клеток. Бактериофаги явно много мельче бактериальных клеток. Не могут ли они представлять собой «высвободившиеся» гены, способные к размножению, лишь попадая в бактериальную клетку? Эта гипотеза стимулировала множество исследований бактериофагов и других вирусов, исследований, частично подтвердивших идею Мёллера.
В этой главе мы рассмотрим, как изучение бактерий и вирусов привело к пониманию химической природы генетического материала и как были определены его физическая структура и свойства. В последующих главах мы проанализируем, какой вклад внесло изучение микроорганизмов в наши представления об организации и механизмах проявления генетического материала.
Дата добавления: 2015-12-16 | Просмотры: 713 | Нарушение авторских прав
|