|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Анна Милтон(методика составления)
Благодаря кроссинговеру генетики получили возможность исследовать порядок расположения генов в хромосомах и строить генетические карты. Принцип построения генетических карт прост. В его основе лежит положение о том, что если кроссинговер между генами происходит редко, то сила сцепления бóльшая, если часто – сцепление слабое. По частоте кроссинговера судят о расстоянии между генами, однако определить порядок расположения генов в хромосоме можно только изучив взаимоотношения между тремя генами, например, А В С. Предположим, что частота кроссинговера между генами А и В равна некоторой величине М, а между генами В и С – N, то частота перекреста между А и С может быть равной либо величине М + N, либо M – N. Такая закономерность распространяется на все гены данной группы сцепления. Вполне очевидно, что объяснить эти факты можно, только признав линейное расположение генов внутри хромосомы. На основании таких фактов в настоящее время для многих организмов составлены генетические карты хромосом. Наиболее подробные карты известны для дрозофилы, душистого горошка, кукурузы, мыши, кролика, курицы, нейроспоры и др. Карту хромосом по всей ее длине строят постепенно, складывая данные о частоте перекреста между двумя последовательно расположенными наиболее близкими генами. В результате генетическая длина карты хромосом может доходить до 100 единиц и более. В поздней профазе мейоза (пахинеме) у гомологичных хромосом могут образовываться несколько хиазм. Это приводит к появлению одиночных или двойных, а также тройных и более сложных кросинговеров (рис. 40). Для обнаружения двойных кросинговеров необходимо наличие гетерозиготности по трем генам, локализованным в одной группе сцепления. Рассмотрим это для трех генов дрозофилы, локализованных в половой хромосоме, а именно: 1) цвет тела: серый (А) или желтый (а); 2) форма щетинок: прямые (В) или загнутые (в); 3) размер крыльев: нормальные (С) или маленькие (с). Для составления карты хромосом необходимо: 1. Скрестить гомозиготные особи с альтернативными признаками и получить F1; 2. Произвести анализирующее скрещивание; 3. Установить кроссоверные особи в процентах среди гибридов анализирующего скрещивания. Ход эксперимента выглядит так:
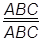 × ♂ × ♂ 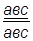
Самки, гетерозиготные по этим трем сцепленным генам, были скрещены с самцами, гомозиготными по рецессивным аллелям изучаемых генов.
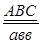 × ♂ × ♂
гаметы: ♀ АВС, авс – некроссоверные аВС, Авс – одиночный кроссинговер на участке А – В АВс, авС – одиночный кроссинговер на участке В – С АвС, аВс – двойной кроссинговер на участке А – С
Рис 40. Типы гамет, образующиеся у самки дрозофилы, гетерозиготной по трем парам генов, находящихся в одной хромосоме
В потомстве анализирующего скрещивания получилось следующее расщепление: Fан 7336 7334 1994 2072 1318 1360 332 350
Всего поучено 22096 мух (7336+7334+…+350), в том числе 14670 – некроссоверных (66,4%). Рассчитаем частоту перекреста и расстояние между генами: 1) А и В: 2) В и С: 3) А и С: Учитывая, что один процент перекреста условно принят за единицу расстояния между генами, можно заключить, что гены А и В отстоят друг от друга дальше, чем В и С. Расстояние между А и С составляет 30,5 кроссоверных единиц, что равно сумме двух расстояний: 18,4 к. е. + 12,1 к. е. = 30,5 к. е. Определим величину двойного кроссинговера (обмен средними участками):
Таким образом, из 22096 особей с перекрестом на первом участке (А-В) было 18,4%, на втором (В-С) – 12,1%, с двойным перекрестом – 3,1%. Фактически перекрест на участке А-В произошел не в 18,4% случаев, а в несколько большем числе, так как и при двойном кроссинговере был перекрест на участке А-В. Для получения общей величины перекреста на данном участке нужно сложить величины 18,4% + 3,1% = 21,5%. Таким же образом следует внести поправку в частоту перекреста и на участке В-С, так как он был не только при одиночном, но и при двойном кроссинговере; получим цифру 15,2% (12,1+3,1). Общее число перекрестов будет равно 36,7% (21,5+15,2). Частота единичных кроссоверов между А и С равна 30,5%, однако в это число входят и двойные. Поскольку каждый двойной кроссовер состоит из двух единичных между крайними генами, то для того, чтобы получить их суммарную частоту между А и С, частоту двойных 3,1 нужно удвоить и прибавить к частоте единичных (6,2 + 30,5 = 36,7).
Исследованиями установлено, что двойной перекрест осуществляется фактически в меньшем числе случаев, чем ожидается. Предполагаемый двойной перекрест можно рассчитать, если известен порядок расположения генов в хромосоме и частота кроссинговера между ними. Напомним, что вероятность совпадения двух отдельных событий равна произведению этих вероятностей для каждого из них. В нашем случае частота перекреста на участке между генами А-В равна 21,5, а между В-С – 15,2%, то вероятность возникновения двух перекрестов в одной тетраде (по одному в каждом из этих участков) составит:
Однако фактически двойную хиазму несли только 3,1% тетрад. Это объясняется явлением интерференции (подавления) двойного кроссинговера, когда кроссинговер в одном локусе препятствует его возникновению в другом. Степень интерференции зависит от расстояния между генами. По мере уменьшения расстояний между генами интерференция увеличивается, и наоборот. Степень интерференции хиазм можно выразить в виде отношения:
Это отношение называют коэффициентом совпадения или коинциденции. В нашем примере он равен 95%, следовательно, часть случаев (5%) двойного кроссинговера не была осуществлена вследствие интерференции. Анна Милтон Дата добавления: 2015-01-18 | Просмотры: 720 | Нарушение авторских прав |

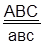 ;
; 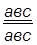 ;
; 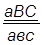 ;
; 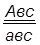 ;
; 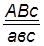 ;
; 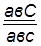 ;
; 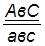 ;
; 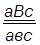 ;
;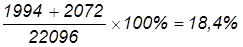 .
.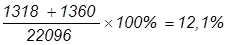 .
.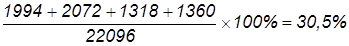 .
.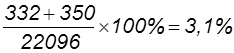 .
.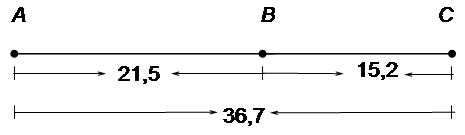 Таким образом, если учесть двойные кроссоверы, генетическая карта, построенная на основе их частот, становится линейной (21,5 + 15,2 = 6,2 + 30,5).
Таким образом, если учесть двойные кроссоверы, генетическая карта, построенная на основе их частот, становится линейной (21,5 + 15,2 = 6,2 + 30,5).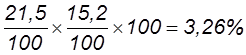 .
.
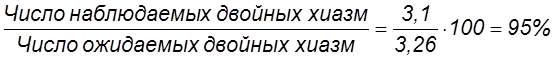 .
.