|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Разнообразие гаметРассмотри мейоз клетки, имеющей три пары хромосом (2n = 6). В этом случае после двух мейотических делений образуются четыре клетки с гаплоидным набором хромосом (n = 3). Поскольку хромосомы каждой пары расходятся в дочерние клетки независимо от хромосом других пар, равновероятно образование восьми тиров гамет с различным сочетанием хромосом, присутствовавших в исходной материнской клетке.
8. Сперматогенез и о(в)огенез. Оплодотворение и его генетическая сущность.
Мужские половые клетки животных - сперматозоиды - образуются в семенниках. В их развитии различают несколько стадий.Первая из них - стадия размноженияпервичных половых клеток - сперматогониев, которые интенсивно делятся путем митоза. Затем эти клетки вступают в профазу Iмейоза и превращаются в сперматоциты, первого порядка. В результате прекращающегося синтеза РНК и, следовательно, белка профазные клетки увеличиваются в размерах. Это - стадия роста. Стадия, во время которой проходят одно за другим два мейотических деления, получила название стадии созревания. В результате первого деления созревания из одного сперматоцита первого порядка образуется два сперматоцита второго порядка. После второго деления из каждого сперматоцита второго порядка возникают две гаплоидные клетки - сперматиды. Таким образом, из одной исходной клетки, вступившей в мейоз, образуются четыре сперматиды. Они имеют гаплоидный набор хромосом, но еще не являются специализированными клетками, способными к движению и проникновению внутрь яйцеклетки. Превращение сперматид в сперматозоиды происходит во время следующей стадии - стадии формирования. Она продолжается несколько суток (у человека, например, почти четыре недели). В это время ядро уменьшается в размерах за счет плотной упаковки хромосом. Резко сокращается объем цитоплазмы, и формируются цитоплазматические структуры, характерные только для сперматозоидов. Рядом с ядром из аппарата Гольджи образуется акросома - пузырек, содержащий ферменты, способные растворить оболочку яйцеклетки в момент оплодотворения. С противоположной стороны ядра или головки спермия возникает длинный жгут, или хвост, придающий сперматозоиду подвижность. В основании хвоста образующие его фибриллы окружаются слоем митохондрий. Между хвостом и головкой располагается центриоль (рис. 75). Все эти преобразования обеспечивают дальнейшее поведение сперматозоида: его подвижность и проникновение в яйцеклетку. Плотная упаковка хромосом в головке сперматозоида и отсутствие цитоплазмы позволяют ему хорошо сохранять свою целостность вне организма и делают его очень устойчивым к неблагоприятным воздействиям внешней среды. Сперматогонии, лежащие непосредственно на базальной мембранеизвитых семенных канальцев, проходят несколько последовательных стадий митотического деления. Общее количество сперматогоний в яичке человека составляет около 1 млрд. Различают две основные категории сперматогоний: сперматогонии A и сперматогонии B. Некоторые сперматогонии А, которые делятся митотически, остаются стволовыми, т. е. сохраняют способность к делению и поддерживают свою популяцию. Остальные дифференцируются в сперматогонии В, которые делятся митотически, дифференцируются в сперматоциты первого порядка и вступают в мейоз. Сперматоциты первого порядка (первичные), соединены между собой межклеточными мостиками, которые остаются при митозесперматогонийблагодаря неполному разделению клеток. Последующие поколения клеток также остаются соединенными между собой, в результате чего образуется синцитий, клетки которого составляют клон. Клетки синцития делятся синхронно, лишь единичные клетки не делятся. В результате мейоза I образуются две дочерние клетки сперматоциты второго порядка, которые далее образуют сперматиды. Таким образом, в результате деления одной сперматогонии образуются четыре сперматиды, каждая из которых обладает гаплоидным набором хромосом.
Развитие и строение половых клеток самок
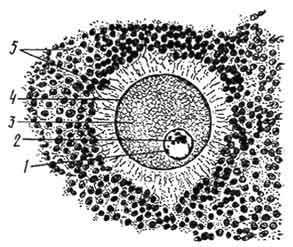
о сравнению со спермиями яйцеклетки, или овоциты, образуются в значительно меньшем количестве, поэтому на одну женскую половую клетку приходится множество мужских Размер яйцеклетки у одного и того же животного превышает размер спермия во много раз. Диаметр овоцитов может варьировать от 100 мкм до нескольких миллиметров, что обусловлено количеством желтка (включение цитоплазмы), необходимого питательного материала для развития зародыша. Яйцеклетки позвоночных овальной формы, неподвижны, состоят из ядра и цитоплазмы. Шаровидной формы и большого размера ядро построено, как и у соматической клетки, из хроматина, ядрышка, кариолимфы и кариолеммы. Ядро содержит гаплоидный (одинарный) набор хромосом. Яйцеклетки млекопитающих относят к гомогаметным, так как в их ядре имеется только Х-хромосома. Ядрышко крупное. Его размер и строение свидетельствуют об интенсивном синтезе рибосом и РНК.
1 - яйцо; 2 - ядрышко; 3 - цитоплазма; 4 - первичная оболочка; 5 - вторичная оболочка. Для цитоплазмы характерна относительно высокая степень базофилии, обладающей сродством с РНК. В ней содержатся свободные рибосомы, эндоплазматическая сеть, комплекс Гольджи, митохондрии, мультивезикулярные тельца, желток. В образовании последнего активное участие принимают эндоплазматическая сеть и комплекс Гольджи. Желток, являясь цитоплазматическим включением, имеет вид гранул, 00шаров и пластин; состоит из протеинов, углеводов, фосфолипидов. Овоциты обладают полярностью. Это объясняется неравномерным расположением желтка и других цитоплазматических структур, в связи с чем в овоците различают два полюса: апикальный и базальный. Периферический слой цитоплазмы яйцеклетки называют кортикальным слоем (cortex - кора). Он лишен полностью желтка, содержит множество митохондрий. Функции у этого слоя - перенос питательных веществ и участие в развитии зародыша на самых ранних стадиях эмбриогенеза. Яйцеклетки покрыты оболочками. Различают первичную, вторичную и третичную оболочки. Первичная оболочка - это плазмолемма, поэтому она присутствует всегда. Вторичная оболочка является производной фолликулярных клеток яичника. Она состоит из фолликулярных клеток и выполняет трофическую, защитную функции, препятствует полиспермии (оплодотворение яйцеклетки многими спермиями) и особенно хорошо развита у млекопитающих. Третичная оболочка овоцита формируется из материала, секретируемого клетками яйцевода. Она играет защитную, а также трофическую функции и развита у пресмыкающихся и птиц. Третичными оболочками яйцеклетки птиц являются белок, подскорлуповая и скорлуповая оболочки яйца. Таким образом, яйцеклетки в связи с выполняемыми ими функциями обладают рядом морфологических особенностей: наличием запаса питательного материала (желтка), кортикального слоя, специальных оболочек, полярностью, генетической однородностью (Х-хромосомой). Классификация яйцеклеток. Усложнение эмбриогенеза в ряду хордовых незначительно сказалось на строении половых клеток самцов, тогда как в яйцеклетках наблюдается постепенное нарастание содержания питательных веществ (желтка). По количеству желтка различают яйцеклетки с малым количеством желтка - олиголецитальные (oligos - мало, lecytos - желток), со средним количеством - мезолецитальные (mesos - средний) и с большим количеством желтка - полилецитальные (роly - много). По месту расположения желтка в цитоплазме различают яйцеклетки с равномерным распределением желтка - изолецитальные, или гомолецитальные (isos, homos - одинаковый, сходный), и с локализацией желтка у одного полюса - телолецитальные (telos - край, конец). Олиголецитальные и изолецитальные яйцеклетки у ланцетника и млекопитающих. Если у ланцетника это связано с наличием в их развитии личиночной стадии (личинка сама для себя добывает питательный материал), то у млекопитающих - в связи с внутриутробным развитием. Мезолецитальные и телолецитальные яйцеклетки - у амфибий, некоторых рыб, а полилецитальные и телолецитальные - у многих рыб, пресмыкающихся, птиц и яйцекладущих млекопитающих. Половое размножение организмов связано с их морфологической и физиологической половой дифференциацией (половой диморфизм) и половым процессом. Половой процесс характеризуется системой приспособительных механизмов: 1) образованием мужских и женских гамет. 2) их слиянием в процессе оплодотворения (сингамия), 3) объединением ядер (кариогамия), 4) синаупсисогомологивдных хромосом в мейозе и перекомбинацией наследственных факторов. Цикл полового размножения охватывает период от момента формирования половых клеток до их нового воспроизведения в следующем поколении. Оплодотворением принято называть побуждение яйца к развитию r результате кариогамии. Оплодотворение представляет собой процесс необратимый — оплодотворенное однажды яйцо не может быть оплодотворено вновь. Сингамия и кариогамия составляют сущность процесса оплодотворения. Однако у некоторых видов воспроизведение нового поколения осуществляется на основе только женской гаметы — яйцеклетки без оплодотворения (девственное размножение). В этом случае половое размножение также заканчивается созреванием гамет. Оба эти способа размножения могут чередоваться у одного и того же вида. В процессе оплодотворения осуществляются следующие важные генетические явления, необходимые для существования вида: 1) восстановление диплоидного набора хромосом, а в пределах диплоидного набора — парности гомологичных (материнских и отцовских) хромосом, разошедшихся в мейозе в процессе образования половых клеток у родительских организмов; 2) обеспечение материальной непрерывности между следующими друг за другом поколениями; 3) объединение в одном индивидууме наследственных свойств материнского и отцовского организмов. Для обеспечения оплодотворения необходимо одновременное созревание гамет материнского и отцовского организма. У перекрестноопыляющихся растений созревание мужских и женских половых клеток может не совпадать во времени, и это несоответствие служит приспособительным механизмом, препятствующим самоопылению. Возможно, что несоответствие во времени созревания половых клеток у разных полов одного вида является одним из путей возникновения перекрестного опыления.
9. Понятие о генотипе и фенотипе, гомо-гетерозиготности, аллельных генах и признаках. 10. Закон единообразия признаков у гибридов в F1 11. Наследование признаков при не – и полном, доминировании и кодоминировании
При полном доминировании фенотип гетерозиготы не отличается от фенотипа доминантной гомозиготы по данному аллелю. Видимо, в чистом виде полное доминирование встречается крайне редко или не встречается вовсе. Например, люди, гетерозиготные по гену гемофилии А(сцепленный с Х-хромосомой рецессивный аллель), имеют половинное количество нормального фактора свертывания по сравнению с гомозиготными по нормальному аллелю людьми, и активность фактора свертывания VIII у них в среднем вдвое ниже, чем у здоровых людей. При неполном доминировании гетерозиготы имеют фенотип, промежуточный между фенотипами доминантной и рецессивной гомозиготы. Например, при скрещивании чистых линий львиного зева и многих других видов цветковых растений с пурпурными и белыми цветками особи первого поколения имеют розовые цветки. При скрещивании чистых линий андалузских кур чёрной и белой окраски в первом поколении рождаются куры серой окраски. На молекулярном уровне самым простым объяснением неполного доминирования может быть как раз двукратное снижение активности фермента или другого белка (если доминантный аллель дает функциональный белок, а рецессивный — дефектный). Некоторые противоположные признаки находятся не в отношении полного доминирования (когда один всегда подавляет другой у гетерозиготных особей), а в отношении неполного доминирования. Например, при скрещивании чистых линий львиного зева с пурпурными и белыми цветками особи первого поколения имеют розовые цветки. При скрещивании чистых линий андалузских кур чёрной и белой окраски в первом поколении рождаются куры серой окраски. При неполном доминировании гетерозиготы имеют признаки, промежуточные между признаками рецессивной и доминантной гомозигот. При кодоминировании, в отличие от неполного доминирования, у гетерозигот признаки проявляются одновременно (смешанно). Типичный пример кодоминирования — наследование групп крови системы АВ0 у человека, где А и В — доминантные гены, а 0 — рецессивный. По этой системе генотип 00 определяет первую группу крови, АА и А0 — вторую, ВВ и В0 — третью, а АВ будет определять четвётртую группу крови. Т.о. всё потомство людей с генотипами АА (вторая группа) и ВВ (третья группа) будет иметь генотип АВ (четвертая группа). Их фенотип не является промежуточным между фенотипами родителей, так как на поверхности эритроцитов присутствуют оба агглютиногена (А и В). Явления кодоминирования и неполного доминирования признаков слегка видоизменяет первый закон Менделя: «Гибриды первого поколения от скрещивания чистых линий особей с противоположными признаками всегда одинаковы по этому признаку: проявляют доминирующий признак, если признаки находятся в отношении доминирования, или смешанный (промежуточный) признак, если они находятся в отношении кодоминирования (неполного доминирования)». Кодоминирование в генных аллелях В случае кодоминирования у гетерозигот полностью проявляются оба аллеля. Наиболее яркий пример кодоминирования - наследование групп крови АВО у человека. Группы крови О (I), А (II), В (III), АВ (IV) детерминируются геном / (для обозначения генов принят курсивный шрифт). Известны три основных аллеля этого гена, два из которых IA и IB доминантные, а один I0 — рецессивный. При гомозиготности IАIА эритроциты имеют только поверхностный антиген А (группа крови А). При гомозиготности IвIв эритроциты несут другой антиген - В (группа крови В). В случае гомозиготности I0I0 эритроциты лишены обоих антигенов (группа крови О). У гетерозигот IАI0 или IBI0 в соответствии с имеющимся антигеном группа крови А или В. У гетерозигот IAIB эритроциты несут оба антигена А и В (гpyппa крови АВ). Как правило, на уровне синтеза полипептидов аллели гетерозигот кодоминантны. Так у гетерозигот по серповидноклеточной анемии HbA/HbS, гемоглобины HbS составляют 35-42%, НbА 60-65%. Полная клиническая картина заболевания (тромбозы мелких сосудов, инфаркты внутренних органов, гемолитическая анемия, гиперплазия костного мозга, нарушения мозгового кровообращения; смерть в возрасте 3-10 лет, иногда позже) проявляется только в случае гомозиготности по HbS. У гомозигот мутантный гемоглобин Hb состоит из двух нормальных цепей и двух мутантных бета-цепей, в 6-м положении которых глутаминовая кислота заменена валином. Заболевание наследуется по аутосомно-рецессивному типу, наличие у гетерозигот до 65% нормального гемоглобина позволяет большинству из них чувствовать себя здоровыми. Однако в холодное время года, при повышенной нагрузке, а также при полетах на самолете у них появляются боли в суставах, сердце, брюшной полости, в области селезенки, что свидетельствует о неполной доминантности нормального аллеля НbА в экстремальных условиях у носителей мутантного аллеля HbS. Таким образом, кодоминантность у гетерозигот HbA/HbS проявляется только на уровне синтеза полипептидных цепей гемоглобина, а заболевание с полной клинической картиной наследуется по аутосомно-рецессивному типу.
12. Возвратное и анализирующее скрещивание
Все вышесказанное относится к анализу наследования признаков, когда гибриды F1 скрещиваются между собой. Однако для генетического анализа может быть использован и другой тип скрещивания — скрещивание гибрида с одной из родительских форм, несущей данную пару аллелей в гомозиготном состоянии. Такое скрещивание гибрида первого поколения с формой, несущей данную пару аллелей (доминантных или рецессивных) в гомозиготном состоянии, называют возвратным скрещиванием, или беккроссом (обозначается Fb); его можно записать следующим образом: Аа х АА или Аа х аа. Эти два скрещивания имеют неодинаковую ценность для генетического анализа и практики селекции. Очевидно, возвратное скрещивание гибрида F1(Aa) с родительской формой, гомозиготной по доминантной аллели (АА), даст однотипное по внешнему виду потомство. Все гаметы родительской формы будут нести доминантную аллель А, а у гибрида образуются гаметы двух сортов — с аллелями А и а. Поэтому в результате случайного сочетания этих гамет при оплодотворении в потомстве от такого скрещивания имеет место расщепление по генотипу в отношении 2Аа: 2АА, или 1: 1, в то время как расщепления по фенотипу не произойдет (1:0). Значительно больший интерес для генетического анализа представляет скрещивание гибрида F1(Aa) с формой, гомозиготной по рецессивному гену, т. е. с формой аа. Поскольку все гаметы этой формы несут рецессивную аллель, характер расщепления в потомстве по фенотипу будет соответствовать качеству гамет гибридного организма. В результате расщепление по гену окраски цветков окажется в отношении 1Аа: 1аа. По характеру расщепления в потомстве от такого скрещивания АаХаа можно проанализировать наследственную структуру гибрида по данному гену. Отсюда и скрещивание гибридного организма с исходной формой, гомозиготной по рецессивному гену, получило название анализирующего скрещивания. Совпадение полученных данных с теоретически ожидаемыми в анализирующем скрещивании служит одним из прямых доказательств равного образования двух сортов гамет. Данным скрещиванием для указанной цели и воспользовался Мендель. С помощью этого метода можно проверить гетерозиготность организма по изучаемой паре генов из любого поколения F1, F2, F3, F4 и т. д. Как будет видно из дальнейшего, подобное скрещивание является очень важным приемом генетического анализа сложных гибридов.
13. Расщепление признаков в F2 при дигибридном и полигибридном скрещивании. Закон независимого расщепления признаков и его объяснение.
Дигибридное скрещивание, Т. е скрещивание родительских форм, различающихся по двум парам признаков. Исходными формами для скрещивания взяты горох с желтыми и гладкими семенами, с другой — горох с зелеными и морщинистыми. При таком скрещивании мы имеем дело с разными парами аллельных генов. Одна такая пара включает гены окраски семян; вторая — гены формы семян. Если для скрещивания взяты гомозиготные формы, то все потомство в первом поколении гибридов будет обладать желтыми гладкими семенами — проявится правило единообразия. В первой паре генов доминантной окажется желтая окраска, рецессивной — зеленая. Во второй паре генов гладкая форма семян доминирует над морщинистой. При скрещивании между собой гибридов первого поколения в их потомстве произойдет расщепление. По фенотипу получатся четыре группы особей в различных численных отношениях: на 9 особей с желтыми гладкими семенами будут приходиться 3 с желтыми морщинистыми, 3 с зелеными гладкими и 1 с зелеными морщинистыми. В кратком виде это расщепление можно представить формулой: 9:3:3:1. Количественные отношения между числом различных фенотипов и генотипов в F2 При дигибридном скрещивании справедливы для аллелей с полным доминированием. При промежуточном характере наследования число фенотипически различных форм будет больше. Если по обоим признакам доминирование неполное, то количество фенотипически различных групп равняется числу генотипически различных групп. Второй закон Менделя. Его называют законом независимого расщепления: расщепление по каждой паре признаков идет независимо от других пар признаков. Три - и полигибридное скрещивание. Мендель проверял закон независимого комбинирования на различных комбинациях пар признаков. Он подтвердил также этот закон, поставив опыт по скрещиванию растений, отличавшихся сразу по трем признакам. Такое скрещивание называется Тригибридным. Например, скрещивание между двумя растениями гороха с разными признаками (желтый, гладкий, морщинистый, цветы пурпурные). Гибриды F1 будут тройными гетерозиготами или тригибридами. Вследствие доминантности семена у таких растений будут гладкими и желтыми, а цветы-пурпурными. Если все гены передаются независимо, то в тригибридном растении образуется восемь типов гамет, причем все с равной вероятностью. В общем случае каждый новый ген увеличивает число типов различных гамет вдвое, а число генетических классов втрое. Особь, гетерозиготная по П Парам генов, может произвести 2n типов гамет и 3n различных генотипов. Число внешне различающихся классов равно числу различных типов гамет при наличии доминирования и числу различных генотипов в отсутствие доминирования.
Закон расщепления (второй закон Менделя) — при скрещивании двух гетерозиготных потомков первого поколения между собой во втором поколении наблюдается расщепление в определенном числовом отношении: по фенотипу 3:1, по генотипу 1:2:1. Скрещиванием организмов двух чистых линий, различающихся по проявлениям одного изучаемого признака, за которые отвечают аллели одного гена, называется моногибридное скрещивание. Явление, при котором скрещивание гетерозиготных особей приводит к образованию потомства, часть которого несёт доминантный признак, а часть — рецессивный, называется расщеплением. Следовательно, расщепление — это распределение доминантных и рецессивных признаков среди потомства в определённом числовом соотношении. Рецессивный признак у гибридов первого поколения не исчезает, а только подавляется и проявляется во втором гибридном поколении. Объяснение[править | править вики-текст] Закон чистоты гамет: в каждую гамету попадает только один аллель из пары аллелей данного гена родительской особи. В норме гамета всегда чиста от второго гена аллельной пары. Этот факт, который во времена Менделя не мог быть твердо установлен, называют также гипотезой чистоты гамет. В дальнейшем эта гипотеза была подтверждена цитологическими наблюдениями. Из всех закономерностей наследования, установленных Менделем, данный «Закон» носит наиболее общий характер (выполняется при наиболее широком круге условий). Гипотеза чистоты гамет. Мендель предположил, что при образовании гибридов наследственные факторы не смешиваются, а сохраняются в неизменном виде. У гибрида присутствуют оба фактора — доминантный и рецессивный, но проявление признака определяет доминантныйнаследственный фактор, рецессивный же подавляется. Связь между поколениями при половом размножении осуществляется через половые клетки — гаметы. Следовательно, необходимо допустить, что каждая гамета несет только один фактор из пары. Тогда при оплодотворениислияние двух гамет, каждая из которых несет рецессивный наследственный фактор, будет приводить к образованию организма с рецессивным признаком, проявляющимся фенотипически. Слияние же гамет, каждая из которых несет доминантный фактор, или же двух гамет, одна из которых содержит доминантный, а другая рецессивный фактор, будет приводить к развитию организма с доминантным признаком. Таким образом, появление во втором поколении рецессивного признака одного из родителей может быть только при двух условиях: 1) если у гибридов наследственные факторы сохраняются в неизменном виде; 2) если половые клетки содержат только один наследственный фактор из аллельной пары. Расщепление потомства при скрещивании гетерозиготных особей Мендель объяснил тем, что гаметы генетически чисты, то есть несут только один ген из аллельной пары. Гипотезу (теперь ее называют законом) чистоты гамет можно сформулировать следующим образом: при образовании половых клеток в каждую гамету попадает только один аллель из пары аллелей данного гена. Известно, что в каждой клетке организма в большинстве случаев имеется совершенно одинаковый диплоидный набор хромосом. Две гомологичные хромосомы обычно содержат каждая по одному аллелю данного гена. Генетически «чистые» гаметы образуются следующим образом:
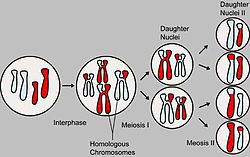
Основные этапы мейоза На схеме показан мейоз клетки с диплоидным набором 2n=4 (две пары гомологичных хромосом). Отцовские и материнские хромосомы обозначены разным цветом. В процессе образования гамет у гибрида гомологичные хромосомы во время I мейотического деления попадают в разные клетки. При слиянии мужских и женских гамет получается зигота с диплоидным набором хромосом. При этом половину хромосом зигота получает от отцовского организма, половину — от материнского. По данной паре хромосом (и данной паре аллелей) образуются два сорта гамет. При оплодотворении гаметы, несущие одинаковые или разные аллели, случайно встречаются друг с другом. В силу статистической вероятности при достаточно большом количестве гамет в потомстве 25 % генотипов будут гомозиготными доминантными, 50 % — гетерозиготными, 25 % — гомозиготными рецессивными, то есть устанавливается отношение 1АА:2Аа:1аа (расщепление по генотипу 1:2:1). Соответственно по фенотипу потомство второго поколения при моногибридном скрещивании распределяется в отношении 3:1 (3/4 особей с доминантным признаком, 1/4 особей с рецессивным). Таким образом, при моногибридном скрещивании цитологическая основа расщепления признаков — расхождение гомологичных хромосом и образование гаплоидных половых клеток в мейозе.
14. Наследование признаков при комплементарном взаимодействии генов
Закон независимого наследования генов еще раз демонстрирует дискретный характер генетического материала. Это проявляется в независимом комбинировании аллелей разных генов и в их независимом действии - фенотипическом выражении. Однако в ряде случаев идентификация фенов сопряжена с некоторыми трудностями. Обратимся к примеру. Комплементарность У популярного генетического объекта плодовой мушки Drosophila melanogaster имеется большое число форм, наследственно различающихся по окраске глаз. У мух так называемого дикого типа или типа, распространенного в природе, глаза темно-красные. Существуют формы с ярко-красными глазами. Этот признак рецессивен по отношению к дикому типу. Он наследуется по моногибридной схеме при скрещивании нормальных мух и мух с ярко-красными глазами. Соответствующий ген обозначается: st (scarlet) - рецессив; st+ - доминант. Существуют также мухи с коричневыми глазами. Это тоже рецессивный признак, наследующийся по моногибридной схеме при скрещивании диких мух и мух с коричневыми глазами. Соответствующий ген обозначается bw (brown) - рецессив; bw+ - доминант1. Если скрестить мух с ярко-красными глазами и мух с коричневыми глазами, то получаются следующие результаты. В Fi все мухи имеют темно-красные глаза (дикий тип), а при скрещивании гибридов первого поколения в F2 появляются четыре класса расщепления: мухи с темно-красными, ярко-красными, коричневыми и белыми глазами в соотношении 9:3:3:1. Для объяснения этого результата обратимся к логике генетического анализа. Результаты, получившиеся в F1, показывают, что существует некоторый тип взаимодействия. Можно предположить, что это взаимодействие аллелей одного гена при моногибридном скрещивании. Однако в F2 появляются четыре класса в соотношении, характерном для дигибридного скрещивания при полном доминировании по обоим признакам, и среди них формируется самый малочисленный класс - мухи с белыми глазами. Если справедливо предположение о том, что это скрещивание дигибридное, то генотипы родительских форм записывают: P ♀ stst bw+ bw+ x ♂ st+st+ bwbw где знак «+» соответствует нормальным (доминантным) аллелям генов: bw и st. Тогда самки и самцы образуют по одному типугамет - соответственно st bw+ и st+bw, а генотип гибридов F1будет: st+st bw+bw. Такие дигетерозиготные мухи должны образовать четыре типа гамет, которые во всевозможных сочетаниях при скрещивании между собой гибридов F1 дадут в F2 следующее расщепление по генотипу. С помощью фенотипических радикалов можно написать следующее расщепление по фенотипу: 9 st+ - bw+ - - с темно-красными глазами 3 st+ - bw bw - с коричневыми глазами 3 st st bw+ - - с ярко-красными глазами 1 st st bw bw - с белыми глазами Учитывая, что аллели st и bw рецессивны, можно объяснить появление первых трех фенотипических классов при расщеплении. При наличии нормальных аллелей st+ и bw+ мухи должны принадлежать к дикому типу по окраске глаз (9 с темно-красными глазами). При гомозиготности только по рецессивной аллели bw bw мухи должны быть с коричневыми глазами (3), так же как при гомозиготности только по st st мухи должны быть с ярко-красными глазами (3). Наконец, остается последний класс - двойной гомозиготный рецессив (1 st st bw bw), который соответствует мухам с белыми глазами. Все эти выводы можно проверить, исследуя далее расщепление при анализирующих скрещиваниях и скрещиваниях между особями F1. Таким образом, предположение о дигибридном расщеплении в рассмотренном скрещивании подтверждается, а новообразование - белоглазые мухи в F2 - результат взаимодействия рецессивных аллелей st и bw. В рассмотренной схеме также наблюдалось взаимодействие генов в F2, в результате которого дрозофилы имели нормальный цвет глаз. Такой тип взаимодействия носит название комплементарности или комплементарного (взаимно дополнительного) действия, когда доминантные аллели обоих генов обусловили нормальный (или дикий) фенотип (под комплементарностью обычно подразумевают именно этот тип взаимодействия генов). В F2, рецессивные аллели тех же генов обусловили появление белоглазых мух. Был приведен пример того, что носит название формально-генетического анализа, при котором полностью абстрагируются от механизмов действия исследуемых генов. Если анализ проведен верно и формальные отношения генов и аллелей выявлены правильно, то последующее выяснение физиологических механизмов, лежащих в основе генных взаимодействий, только подтверждает и конкретизирует выводы. Биохимический механизм взаимодействия аллелей генов st и bw исследован достаточно подробно. Известно, что у дрозофилы окраска глаз обусловлена синтезом двух пигментов - красного и бурого. Рецессивная аллель bw в гомозиготе прерывает синтез красного пигмента, поэтому глаза содержат только бурый пигмент. Рецессивная аллель st в гомозиготе блокирует синтез бурого пигмента, вследствие чего в глазах мух содержится только красный пигмент. Когда в дигетерозиготе оказываются нормальные аллели обоих генов, синтезируются оба пигмента. Результат - комплементарное взаимодействие нормальных аллелей, наблюдаемое в F1. Если в F2 в гомозиготе оказываются и bwbw, и stst, то не синтезируются ни красный, ни бурый пигменты, и глаза оказываются белыми, неокрашенными. Можно рассмотреть и другой тип комплементарного взаимодействия генов у дрозофилы, если идти от метаболического эффекта генов к схеме их взаимодействия. Вновь обратимся к биосинтезу пигментов глаза у дорозофилы. Известно, что кроме рецессивных аллелей st синтез бурого пигмента блокируют и рецессивные аллели гена puгрle (pr). Фенотип гомозигот stst и ргрт - ярко-красные глаза. При скрещивании таких мух в F1 глаза нормальные - темно-красные, поскольку работают оба гена - комплементарно взаимодействуют их доминантные аллели. В F2 наблюдается следующее соотношение фенотипов: 9 с темно-красными и 7 с ярко-красными глазами. Это объясняется тем, что выход в гомозиготу любой из двух рецессивных аллелей достаточен для блокирования синтеза пигмента, тем более, когда и рг, и st находятся в гомозиготе. Это тоже пример комплементарного взаимодействия, но без новообразования в F2. По типу комплементарное™ взаимодействуют гены, контролирующие разные этапы одного и того же метаболического пути. Однако для многих морфологических признаков неизвестен биохимический механизм их реализации, поэтому приходится ограничиваться констатацией формально-генетической схемы их наследования. Так, по типу комплементарности взаимодействуют гены, определяющие форму гребня кур, форму плода у тыквы и др.
15. Наследование признаков при эпистазе. Гены эпи- и гопистатичные, ингибиторы и супрессоры.
Вернемся к анализу взаимодействия генов рr и st у дрозофилы. Соотношение фенотипических классов в F2 можно представить себе и как следствие того, что рецессивная аллель рr в гомозиготе препятствует проявлению доминантной аллели st+. Точно так же рецессивная аллель st в гомозиготе препятствует проявлению доминантной аллели рr+. Действительно, то, что известно о генетическом контроле синтеза бурого глазного пигмента у дрозофилы, вполне соответствует предложенному здесь объяснению. Такой тип взаимодействия носит название эпистатического, или эпистаза, и условно изображается: рr > st+ и st > pr+. В данном случае рецессивная аллель рr эпистатична по отношению к доминантной аллели st+, a st эпистатична по отношению к рr+. Данный случай взаимодействия генов называют также двойным рецессивным эпистазом. По изменению числа и соотношения классов дигибридного расщепления в F2 рассматривают несколько типов эпистатических взаимодействий: простой рецессивный эпистаз (а > В; а> Ь или b > А; Ь > а), который выражается в расщеплении 9:3:4; простой доминаданый эпистаз (А> В; А> b или В > А; B > а) с расщеплением 12:3:1 и т.д. Один ген, подавляющий действие другого, называют эпистатическим геном, ингибитором или супрессором. Подавляемый ген носит название гипостатического. Как уже показано, констатация того или иного типа взаимодействия генов в дигибридном скрещивании условна. Тем не менее, при кажущемся нарушении закона независимого наследования (появлении неожиданных классов в расщеплении или уменьшении числа классов), связанного с взаимодействием двух генов, всегда можно свести наблюдаемые соотношения в F2 к классическому 9:3:3:1. При этом важно понять, какие классы объединились, и тогда интерпретировать тип взаимодействия. Необходимо также отметить, что само словосочетание «взаимдействие генов» условно. В действительности взаимодействуют продукты генов, а не сами гены, так что правильнее было бы говорить о взаимодействии фенов, а не о взаимодействии генов. Отсюда понятно, что судить о том, с каким скрещиванием имеет дело экспериментатор: моногибридным, дигибридным или полигибридным - можно только на основании результатов полного гибридологического анализа 16. Наследование количественных признаков. Полимерия.
Наряду с комплементарным и эпистатическим принято также рассматривать взаимодействие генов по типу полимерии. В этом случае разные гены как бы дублируют действие друг друга, и одной доминантной аллели любого из взаимодействующих генов достаточно для проявления изучаемой фенотипической характеристики. Так, при скрещивании растений пастушьей сумки с треугольными плодами (стручками) и с овальными плодами в F1 образуются растения с плодами треугольной формы. При их самоопылении в F2 наблюдается расщепление на растения с треугольными и овальными стручками в соотношении 15:1. Это объясняется тем, что существуют два гена, действующих эднозначно. В этих случаях их обозначают одинаково (А1 и А2). Тогда все генотипы: А1 - А2 -, А1 - а2а2, а2а2А2 - будут иметь одинаковую фенотипическую характеристику - треугольные стручки, и только растения а1а1а2а2 будут отличаться - образовывать овальные стручки. Это случай так называемый некумулятивной полимерии. Однозначные, или полимерные, гены могут действовать и по типу кумулятивной полимерии. Так, шведский генетик Г. Нильсон-Эле в 1908 г. описал серию однозначно действующих генов, которые определяют окраску эндосперма зерен пшеницы. При этом интенсивность окраски зерен оказалась пропорциональной числу доминантных аллелей разных генов в тригибридном скрещивании. Наиболее окрашенными были зерна А1А1А2А2А3А3, а зерна а1а1а2а2а3а3 не имели пигмента. Между этими крайними типами при расщеплении в F2 наблюдались промежуточные варианты в соотношении 1:6:15:20:15:6:1. По типу кумулятивной полимерии наследуются многие количественные признаки, например цвет кожи у человека; молочность, яйценоскость, масса и другие признаки сельскохозяйственных животных; длина колоса у злаков, содержание сахара в корнеплодах сахарной свеклы и др. Изучением наследования таких признаков занимается специальный раздел генетики - генетика количественных признаков, которая важна прежде всего для селекции и разработки проблем микроэволюции.
17. Особенности расщепления F2 при сцеплении двух или нескольких генов
Дальнейшие исследования генетиков показали, что законы Менделя о независимом наследовании признаков при дигибридном скрещивании применимы лишь тогда, когда разные гены располагаются в разных парах гомологичных хромосом. В том случае, если два гена находятся в одной паре гомологичных хромосом, расщепление в потомстве гибридов будет другим. У любого организма генов значительно больше, чем хромосом. Например, у человека имеется около миллиона генов, а хромосом всего 23 пары. Следовательно, в одной хромосоме размещается в среднем несколько тысяч генов. Гены, расположенные в одной хромосоме, называют сцепленными. Все гены этой хромосомы образуют группу сцепления, которая при мейозе обычно попадает в одну гамету. Значит, гены, входящие в одну группу сцепления, не подчиняются закону независимого наследования, а при дигибридном скрещивании вместо ожидаемого расщепления по фенотипу в соотношении 9:3:3:1 дают соотношение 3:1, как при моногибридном скрещивании. Закономерности сцепленного наследования были установлены американским биологом Томасом Морганом (1866-1945). В качестве объекта он использовал плодовую муху дрозофилу. У дрозофилы окраску тела и длину крыльев определяют следующие пары аллелей: А - серое тело, а - черное тело, В - длинные крылья, b - зачаточные крылья. Гены, отвечающие за окраску тела и длину крыльев, находятся в одной паре гомологичных хромосом и наследуются сцепленно.
При скрещивании дрозофилы с серым телом и длинными крыльями с дрозофилой, имеющей черное тело и зачаточные крылья, все гибриды первого поколения имели серую окраску тела и длинные крылья. При дальнейшем скрещивании между собой гибридных мух первого поколения в F2 не произошло ожидаемого расщепления по фенотипу 9:3:3:1. Вместо этого в F2 были получены мухи с родительскими фенотипами в соотношении примерно 3:1. Появление в F2 двух фенотипов вместо четырех позволило сделать вывод, что гены окраски тела и длины крыльев дрозофил находятся в одной хромосоме. Так был установлен закон Т.Моргана: гены, расположенные в одной хромосоме, наследуются совместно - сцепленно, то есть наследуются преимущественно вместе. Однако при дигибридном скрещивании при сцепленном наследовании признаков не всегда появляются особи только двух фенотипов. Иногда появляются особи еще двух фенотипов с перекомбинацией (новым сочетанием) родительских признаков: серое тело - зачаточные крылья, черное тело - длинные крылья. (Особей с такими фенотипами немного - около 8,5% каждого типа.) Почему же нарушается сцепление генов и появляются особи с новыми фенотипами? Было установлено, что сцепление генов может быть полным и неполным. Полное сцепление наблюдается в том случае, если скрещиваются серый самец с длинными крыльями и самка с черным телом и зачаточными крыльями. Расщепление по фенотипу в этом случае будет 1:1, то есть наблюдается полное сцепление генов в одной хромосоме. При скрещивании серой длиннокрылой самки с самцом, имеющим черное тело и зачаточные крылья, расщепление по фенотипу будет примерно 41,5:41,5:8,5:8,5, что характеризует неполное сцепление. Причина нарушения сцепления заключается в том, что в ходе мейоза происходит кроссинговер и гомологичные хромосомы обмениваются своими участками. В результате гены, расположенные в одной из гомологичных хромосом, оказываются в другой хромосоме. Возникают новые сочетания признаков. У самцов дрозофил в мейозе кроссинговер не происходит, поэтому при скрещивании серого длиннокрылого самца дрозофилы с рецессивной самкой с черным телом и зачаточными крыльями сцепление будет полным. Неполное сцепление наблюдается в том случае, если самка гетерозиготна, а самец гомозиготен. В данном примере кроссинговер происходит примерно у 17% самок. Таким образом, если не происходит перекреста хромосом и обмена генами, то наблюдается полное сцепление генов. При наличии кроссинговера сцепление генов бывает неполным. Благодаря перекресту хромосом возникают новые сочетания генов и признаков. Чем дальше друг от друга расположены гены в хромосоме, тем больше вероятность перекреста между ними и обмена участками хромосом. Количество разных типов гамет будет зависеть от частоты кроссинговера или расстояния между анализируемыми генами. Расстояние между генами исчисляется в морганидах: единице расстояния между генами, находящимися в одной хромосоме, соответствует 1% кроссинговера. Такая зависимость между расстояниями и частотой кроссинговера прослеживается только до 50 морганид. Сравните результаты скрещивание дрозофил:
Результатом исследований Т.Х.Моргана стало создание им хромосомной теории наследственности: 1. Гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов, причем набор генов каждой из негомологичных хромосом уникален; 2. Каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены; 3. Гены расположены в хромосомах в определенной линейной последовательности; 4. Гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов; 5. Сцепление генов может нарушаться в процессе кроссинговера; это приводит к образованию рекомбинантных хромосом; 6. Частота кроссинговера является функцией расстояния между генами: чем больше расстояние, тем больше величина кроссинговера (прямая зависимость); 7. Каждый вид имеет характерный только для него набор хромосом - кариотип.
18. Наследование пола и понятие о половых хромасомах
Хромосомная теория показала, в чем состоит внутренний механизм детерминации пола и почему в природе в большинстве случаев рождается половина особей мужского и половина особей женского пола.. Развитие признаков, ограниченных полом, обусловлено генами, расположенными в аутосомах обоих полов, но проявляются они только у особей одного пола (яйценоскость у кур, молочность у коров).. Развитие признаков, контролируемых полом, обусловлено генами, расположенными также в аутосомах обоих полов, но степень и частота проявления их (экспрессивность и пенетрантность) разная у особей разного пола. Это особенно заметно проявляется у гетерозигот, у которых происходит сдвиг доминантности. Изменение доминантности гена обусловлено влиянием половых гормонов (облысение и нормальный рост волос у человека). Пол у большинства животных и растений определяется генетически в момент оплодотворения. При исследовании кариотипов многих животных было установлено, что у женского организма каждая хромосома имеет парную (идентичную по размерам, морфологии и содержанию генов), а у мужских организмов имеются две непарные хромосомы, которые резко отличаются по величине, морфологии и заключенной в них генетической информации. При дальнейшем исследовании было показано, что эти непарные хромосомы и определяют пол организма. Их назвали половыми, или гетерохромосомами, в отличие от остальных - аутосом. Большую из непарных хромосом, одинаковую у мужского и женского организмов, назвали Х-хромосомой, а меньшую, имеющуюся только у мужских организмов, — У-хромосомой. Хромосомная теория пола К. Корренса (1907). Суть ее заключается в том, что пол будущего потомка определяется сочетанием половых хромосом в момент оплодотворения. Пол, имеющий одинаковые половые хромосомы, называют гомогаметным, так как он дает один тип гамет, а имеющий тип гамет, а имеющий разные — гетерогаметным, так как он образует два типа гамет. У человека, всех млекопитающих, мухи дрозофилы гомогаметный пол женский, а гетерогаметный – мужской. Мужской и женский пол различаются наличием особой пары половых хромосом. Пара половых хромосом гомогаметного пола состоит из двух одинаковых хромосом в виде ХХ. В некоторых крайних случаях гетерогаметный пол совершенно теряет второй компонент пары половых хромосом в виде Y-хромосомы. В этих случаях половые хромосомы представлены одной хромосомой (тип ХО). Гетерогаметным полом у различных форм бывает разный пол. У человека, дрозофилы и других форм гетерогаметен мужской пол, который обладает XY-хромесомами.
19. Наследование признаков, сцеплённых с полом, ограниченных и зависимых от пола.
Морган и его сотрудники заметили, что наследование окраски глаз у дрозофилы зависит от пола родительских особей, несущих альтернативные аллели. Красная окраска глаз доминирует над белой. При скрещивании красноглазого самца с белоглазой самкой в F1, получали равное число красноглазых самок и белоглазых самцов. Однако при скрещивании белоглазого самца с красноглазой самкой в F1 были получены в равном числе красноглазые самцы и самки. При скрещивании этих мух F1, между собой были получены красноглазые самки, красноглазые и белоглазые самцы, но не было ни одной белоглазой самки. Тот факт, что у самцов частота проявления рецессивного признака была выше, чем у самок, наводил на мысль, что рецессивный аллель, определяющий белоглазость, находится в Х - хромосоме, а Y - хромосома лишена гена окраски глаз. Чтобы проверить эту гипотезу, Морган скрестил исходного белоглазого самца с красноглазой самкой из F1. В потомстве были получены красноглазые и белоглазые самцы и самки. Из этого Морган справедливо заключил, что только Х - хромосома несет ген окраски глаз. В Y - хромосоме соответствующего локуса вообще нет. Это явление известно под названием наследования, сцепленного с полом. Гены, находящиеся в половых хромосомах, называют сцепленными с полом. В Х-хромосоме имеется участок, для которого в Y-хромосоме нет гомолога. Поэтому у особей мужского пола признаки, определяемые генами этого участка, проявляются даже в том случае, если они рецессивны. Эта особая форма сцепления позволяет объяснить наследование признаков, сцепленных с полом. При локализации признаков как в аутосоме, так и в Х- b Y-хромосоме наблюдается полное сцепление с полом. У человека около 60 генов наследуются в связи с Х-хромосомой, в том числе гемофелия, дальтонизм (цветовая слепота), мускульная дистрофия, потемнение эмали зубов, одна из форм агаммглобулинемии и другие. Наследование таких признаков отклоняется от закономерностей, установленных Г.Менделем. Х-хромосома закономерно переходит от одного пола к другому, при этом дочь наследует Х-хромосому отца, а сын Х-хромосому матери. Наследование, при котором сыновья наследуют признак матери, а дочери - признак отца получило, название крисс-кросс (или крест-накрест). Известны нарушения цветового зрения, так называемая цветовая слепота. В основе появления этих дефектов зрения лежит действие ряда генов. Красно- зеленая слепота обычно называется дальтонизмом. Еще задолго до появления генетики в конце XVIII и в XIX в. было установлено, что цветовая слепота наследуется согласно вполне закономерным правилам. Так, если женщина, страдающая цветовой слепотой, выходит замуж за мужчину с нормальным зрением, то у их детей наблюдается очень своеобразная картина перекрестного наследования. Все дочери от такого брака получат признак отца, т.е. они имеют нормальное зрение, а все сыновья, получая признак матери, страдают цветовой слепотой (а-дальтонизм, сцепленный с Х-хромосомой) Р Ха Ха х Ха y Ха Ха,y F1 Ха Ха, Хаy В том же случае, когда наоборот, отец является дальтоником, а мать имеет нормальное зрение, все дети оказываются нормальными. В отдельных браках, где мать и отец обладают нормальным зрением, половина сыновей может оказаться пораженными цветовой слепотой. В основном наличие цветовой слепоты чаще встречается у мужчин. Э.Вильсон объяснил наследование этого признака, предположив, что он локализовал в Х-хромосоме и что у человека гетерогаметным (XY) является мужской пол. Становится вполне понятным, что в браке гомозиготной нормальной женщины (Ха Ха) с мужчиной дальтоником (Хаy) все дети рождаются нормальными. Однако при этом, все дочери становятся скрытыми носителями дальтонизма, что может проявиться в последующих поколениях. Другим примером наследования сцепленного с полом, может послужить рецессивныйполулетальный ген, вызывающий несвертываемость крови на воздухе - гемофилию. Это заболевание появляется почти исключительно только у мальчиков. При гемофилии нарушается образование фактора VIII, ускоряющего свертывание крови. ген, детерминирующий синтех фактора VIII, находится в участке Х-хромосомы, недоминантным нормальным и рецессивным мутантным. Возможны следующие генотипы и фенотипы: |Генотипы |Фенотипы | |Хн Хн |Нормальная женщина | |Хн Хn |Нормальная женщина (носитель) | |Хнy |Нормальный мужчина | |Хny |Мужчина гемофилик |
В гомозиготном состоянии у женщин ген гемофилии летален. Особей женского пола, гетерозиготных по любому из сцепленных с полом признаков, называют носителями соответствующего рецессивного гена. Они фенотипически нормальны, но половина их гамет несет рецессивный ген. Несмотря на наличие у отца нормального гена, сыновья матерей-носителей с вероятностью 50% будут страдать гемофилией. Один из наиболее хорошо документированных примеров наследования гемофилии мы находим в родословной потомков английской королевы Виктории. Предполагают, что ген гемофилии возник в результате мутации у самой королевы Виктории или у одного из ее родителей. Среди унаследовавших это врожденное заболевание - цесаревич Алексей, сын последнего русского царя Николая II. Мать цесаревича, царица Александра Федоровна (Алиса, рис.2), получила от своей бабушки королевы Виктории ген гемофилии и передала его в четвертом поколении бывшему наследнику царского престола. На рис.2 показано, как этот ген передавался ее потомкам. Один из сцепленных с полом рецессивных генов вызывает особый тип мышечной дистрофии (тип Дюмена). Эта дистрофия проявляется в раннем детстве и постепенно ведет к инвалидности и смерти ранее 20-летнего возраста. Потому мужчины с дистрофией Дюмена не имеют потомства, а женщины гетерозиготные по гену этого заболевания, вполне нормальны. Среди доминантных признаков, связанных с Х-хромосомой, можно указать на ген, который вызывает недостаточность органического фосфора в крови. В результате, при наличии этого гена, часто развивается рахит, устойчивый к лечению обычными дозами витамина А. В этом случае картина сцепленного с полом наследования заметно отличается от того хода передачи по поколениям, который был описан для рецессивных болезней. В браках девяти больных женщин со здоровыми мужчинами среди детей была половина больных девочек и половина мальчиков. Здесь, в соответствии с характером наследование доминантного гена, в Х-хромосомах произошло расщепление в отношении 1:1:1:1. Другим примером доминантного гена, локализованного в Х-хромосоме человека, может послужить ген, вызывающий дефект зубов, приводящий к потемнению эмали зубов. Так как гетерогаметный пол гемизиготен по сцепленным с полом генам, то эти гены всегда проявляются в их фенотипе, даже если они рецессивны. Большинство генов, имеющихся в Х-хромосоме, в Y-хромосоме отсутствует, однако определенную генетическую информацию она все-таки несет. Различают два типа такой информации: во-первых, содержащуюся в генах, присутствующих только в Y-хромосоме, и, во-вторых, в генах, присутствующих как в Y-, так и в Х-хромосоме (гемфрагический диатез). Y-хромосома передается от отца всем его сыновьям, и только им. Следовательно, для генов, содержащихся только в Y-хромосоме, характерно голандрическое наследование, т.е они передаются от отца к сыну и проявляются у мужского пола. У человека в Y-хромосоме содержатся по крайней мере три гена, один из которых необходим для дифференциации семенников, второй требуется для проявления антигена гистосовместимости, а третий оказывает влияние на размер зубов. Y-хромосома имеет немного признаков, среди которых есть патологические. Патологические признаки наследуются по параллельной схеме наследования (100%-ое проявление по мужской линии). К ним относят: 1) облысение; 2) гипертрихоз (оволосенение козелка ушной раковины в зрелом возрасте); 3) наличие перепонок на нижних конечностях; 4) ихтиоз (чешуйчатость и пятнистое утолщение кожи).
2.3. Наследование признаков, контролируемых полом.
Имеется ряд признаков, контролируемых генами, расположенными в аутосомах, однако для проявления этих признаков необходима определенная среда, создаваемая генами, находящимися в половых хромосомах (например, гены, определяющие мужские признаки, находятся в аутосомах, и их фенотипические эффекты маскируются наличием пары Х-хромосом, в присутствии одной Х-хромосомы мужские признаки проявляются. Такие признаки называются обусловленными или контролируемыми полом. Появление лысины - аутосомно-доминантный признак, но проявляется практически только у мужчин при наследовании, контролируемом полом, у женщин подавляются гены, детерминирующие рост бороды.
20. Практическое использование сцепленных с полом признаков у кур и тутового шелкопряда
Раннее распознавание пола используется для сортировки вылупившихся цыплят на петушков и курочек по окраске оперения, сцепленной с полом, а также для «сверхранней» сортировки по полу тутового шелкопряда. Под действием ионизирующего облучения у шелкопряда пересажена аутосома с доминантным геном, обусловливающим тёмную окраску яиц тутового шелкопряда, на половую W- хромосому. Сцепление хромосом стойко передаётся по наследству. Те яйца, в которые попадает W- хромосома с пересаженным доминантным геном, приобретают тёмный цвет и развиваются в самок, в то время как яйца мужского пола, не получив доминантного гена, остаются непигментированными. Фотоэлектрические автоматы с большой скоростью разделяют разноокрашенные яйца по полам. Выведенные таким способом (В. А. Струнников и Л. М. Гуламова) меченые по полу породы шелкопряда находят практическое применение в советском шелководстве.
21. Роль ДНК в сохранении, передаче и реализации наследственной информации
Генетическая информация - программа свойств организма, получаемая от предков и заложенная в наследственных структурах в виде генетического кода. Предполагается, что становление генетической информации шло по схеме: геохимические процессы - минералообразование - эволюционный катализ (автокатализ). Почти вся генетическая информация хранится в ядре клетки. Давайте рассмотрим, что она собой представляет и в каком виде она находится. Возможно, что первые примитивные гены представляли собой микрокристаллические кристаллы глины, причем каждый новый слой глины выстраивается в соответствии с особенностями строения предыдущего, как бы получая от него информацию о строении. Реализация генетической информации происходит в процессе синтеза белковых молекул с помощью трех РНК: информационной (иРНК), транспортной (тРНК) и рибосомальной (рРНК). Процесс передачи информации идет: - по каналу прямой связи: ДНК - РНК - белок; и - по каналу обратной связи: среда - белок - ДНК. Живые организмы способны получать, сохранять и передавать информацию. Причем живым организмам присуще стремление полученную информацию о себе и окружающем мире использовать максимально эффективно. Наследственная информация, заложенная в генах и необходимая живому организму для существования, развития и размножения передается от каждого индивида его потомкам. Эта информация определяет направление развития организма, и в процессе взаимодействия его с окружающей средой реакция на ее индивида может искажаться, обеспечивая тем самым эволюцию развития потомков. В процессе эволюции живого организма возникает и запоминается новая информация, в том числе для него возрастает ценность информации. В ходе реализации наследственной информации в определенных условиях внешней среды формируется фенотип организмов данного биологического вида. Генетическая информация определяет морфологическое строение, рост, развитие, обмен веществ, психический склад, предрасположенность к заболеваниям и генетические пороки организма. Многие ученые, справедливо подчеркивая роль информации в становлении и эволюции живого, отмечали это обстоятельство в качестве одного из главных критериев жизни. Так, В.И. Карагодин считает: "Живое есть такая форма существования информации и кодируемых ею структур, которая обеспечивает воспроизведение этой информации в подходящих условиях внешней среды". Связь информации с жизнью отмечает и А.А. Ляпунов: "Жизнь - это высокоупорядоченное состояние вещества, использующее для выработки сохраняющихся реакций информацию, кодируемую состояниями отдельных молекул". Известный наш астрофизик Н.С. Кардашев также подчеркивает информационную составляющую жизни: "Жизнь возникает благодаря возможности синтеза особого рода молекул, способных запоминать и использовать вначале самую простую информацию об окружающей среде и собственной структуре, которую они используют для самосохранения, для воспроизводства и, что для нас особенно важно, получения еще большего количества информации". На эту способность живых организмов сохранять и передавать информацию обращает внимание в своей книге "Физика бессмертия" эколог Ф. Типлер: "Я определяю жизнь как некую закодированную информацию, которая сохраняется естественным отбором". Более того, он считает, если это так, то система жизнь - информация является вечной, бесконечной и бессмертной. Раскрытие генетического кода и установление закономерностей молекулярной биологии показали необходимость соединения современной генетики и дарвиновской теории эволюции. Так родилась новая биологическая парадигма - синтетическая теория эволюции (СТЭ), которую можно рассматривать уже как неклассическую биологию. Основные идеи эволюции Дарвина с его триадой - наследственностью, изменчивостью, естественным отбором - в современном представлении эволюции живого мира дополняются представлениями не просто естественного отбора, а такого отбора, который детерминирован генетически. Началом разработки синтетической или общей эволюции можно считать работы С.С. Четверикова по популяционной генетике, в которых было показано, что отбору подвергаются не отдельные признаки и особи, а генотип всей популяции, но осуществляется он через фенотипические признаки отдельных особей. Это приводит к распространению полезных изменений во всей популяции. Таким образом, механизм эволюции реализуется как через случайные мутации на генетическом уровне, так и через наследование наиболее ценных признаков (ценности информации!), определяющих адаптацию мутационных признаков к окружающей среде, обеспечивая наиболее жизнеспособное потомство. Сезонные изменения климата, различных природные или техногенные катастрофы с одной стороны, приводят к изменению частоты повторяемости генов в популяциях и, как следствие, к снижению наследственной изменчивости. Этот процесс иногда называют дрейфом генов. А с другой - к изменениям концентрации различных мутаций и уменьшению разнообразия генотипов, содержащихся в популяции, что может привести к изменениям направленности и интенсивности действия отбора. Генетический код – это система записи генетической информации в молекуле нуклеиновой кислоты о строении молекулы полипептида, а именно, о количестве, последовательности расположения и типах аминокислот. В одном гене записана информация об одной полипептидной цепочке, т.е. о первичной структуре белка. Генетический код характеризуется триплетностью, т.е. три нуклеотида, расположенные последовательно в цепочке нуклеиновой кислоты (ДНК или РНК), образуют триплет или кодон (кодовое слово), который кодирует одну аминокислоту и ее местоположение в пептидной цепи. Кодоны различаются последовательностью и типами нуклеотидов (азотистых оснований). Существует 64 типа кодонов, что соответствует количеству возможных сочетаний из 4 (4 типа нуклеотидов, различающихся азотистыми основаниями) по 3 (43). 61 из них – информативные кодоны, они определяют (кодируют) аминокислоты. 3 кодона (в ДНК – АТТ, АТЦ, АЦТ, соответственно в иРНК – УАА, УАГ, УГА) называют стоп-кодонами, они обеспечивают окончание синтеза белковой цепочки. Кодон ТАЦ в ДНК или АУГ в иРНК (кодирует аминокислоту метионин) – стартовый, т.е. стоит первым в гене и с него начинается синтез пептида. Дата добавления: 2015-02-02 | Просмотры: 2167 | Нарушение авторских прав |