|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Межклеточные контакты.
Соединения между клетками в составе тканей и органов многоклеточных животных организмов могут образовываться сложными специальными структурами, которые называют собственно, межклеточными контактами. Эти структурированные межклеточные контакты особенно выражены в покровных пограничных тканях, в эпителиях. Возможно, что первичное обособление пласта клеток, связанных друг с другом с помощью специальных структурированных межклеточных контактов, в филогенезе животных обеспечило образование и развитие тканей и органов. Благодаря электронной микроскопии накопилось множество данных об ультраструктуре этих соединительных образований. К сожалению, их биохимический состав и молекулярная структура еще недостаточно точно изучены. Изучая соединения клеток в эпителиальных пластах, можно Такое разнообразие контактов может встречаться при объединении однородных клеток. Например, в печени встречаются все
Схема строения межклеточных контактов. 1- простой контакт, 2- «замок», 3- плотный замыкающий контакт, 4 –промежуточный контакт, 5- десмосома, 6 – щелевидный контакт
Схема строения межклеточных контактов гепатоцитов крысы: nc- простой контакт, з – «замок», д – десмосома, ск – соединительный комплекс, зс – зона слипания, плотный контакт; жк – желчный капилляр, щк – щелевидный контакт.
Простой контакт, встречающийся среди большинства прилежащих друг к другу клеток различного происхождения. Большая часть поверхности контактирующих клеток эпителия так же связана с помощью простого контакта. где плазматические мембраны соприкасающихся клеток разделены пространством 15 — 20 нм. Как уже говорилось, это пространство представляет собой надмембранные компоненты клеточных поверхностей. Ширина щели между мембранами клеток может быть и больше 20 нм, образуя расширения, полости, но не меньше 10 нм. Со стороны цитоплазмы к этой зоне плазматической мембраны не примыкают никакие специальные дополнительные структуры. Соединение типа «замка» представляет собой выпячивание плазматической мембраны одной клетки в инвагинат (впячивание) другой. На срезе такой тип соединения напоминает плотничий шов. Межмембранное пространство и цитоплазма в зоне «замков» имеют те же характеристики, что и в областях простого контакта. Плотный замыкающий контакт — это зона, где внешние слои двух плазматических мембран максимально сближены. Часто видна трехслойность мембраны в этом контакте: два внешних осмиофильных слоя обеих мембран сливаются в общий слой толщиной 2 — 3 нм. Слияние мембран происходит не по всей площади плотного контакта, а представляет собой ряд точечных слияний мембран; Со стороны цитоплазмы, в этой зоне часто встречаются многочисленные фибриллы около 8 нм в диаметре, располагающиеся параллельно поверхности плазмалеммы. Такого типа контакты были обнаружены между фибробластами в культуре ткани, между эмбриональным эпителием и клетками мезенхимы. Очень характерна эта структура для эпителиев, особенно железистых и кишечных. В последнем случае плотный контакт образует сплошную зону слияния плазматических мембран, опоясывающую клетку в апикальной (верхней, смотрящей в просвет кишечника) ее части. Таким образом, каждая клетка пласта как бы обведена лентой этого контакта. Такие структуры при специальных окрасках можно видеть и в световом микроскопе. Они получили у морфологов название замыкающих пластинок. Оказалось, что в данном случае роль замыкающего контакта заключается не только в механическом соединении клеток друг с другом. Эта область - контакта непроницаема для макромолекул и ионов и тем самым, она запирает, перегораживает межклеточные полости (и вместе с ними собственно внутреннюю среду организма) от внешней среды (в данном случае просвет кишечника) Замыкающий или плотный контакт встречается между всеми типами эпителия (эндотелий, мезотелий, эпендима) Промежуточный контакт (или зона слипания) В этом месте межмембранное расстояние несколько "расширено (до 25 — 30 нм) и в отличие от простого контакта заполнено плотным содержимым, вероятнее всего, белковой природы. Это межмембранное вещество
Функциональная роль десмосом заключается главным образом в механической связи между клетками. Богатство десмосомами клеток покровного эпителия дает ему возможность быть жесткой и одновременно эластичной тканью. Контакты промежуточного типа встречаются не только среди эпителиальных клеток. Сходные структуры обнаружены между клетками гладкой мускулатуры, между клетками мышц сердца У беспозвоночных животных помимо указанных типов соединений встречаются перегородчатые десмосомы. В этом случае межмембранное пространство заполнено плотными перегородками, идущими перпендикулярно мембранам. Эти перегородки (септы) могут иметь вид лент или пчелиных сот (сотовидная десмосома) Щелевидный контакт представляет собой область протяженностью 6,5—3 мкм, где плазматические мембраны разделены промежутком в 2—3 нм, что после осмирования придает всей этой структуре семислойный вид. Со стороны цитоплазмы никаких специальных примембранньтх структур не обнаруживается. Этот тип соединения встречается во всех типах тканей. Функциональная роль щелевидного контакта заключается, видимо, в передаче ионов и молекул от клетки к клетке. Например, в сердечной мышце передача потенциала действия от клетки к клетке происходит через этот тип контакта, где ионы могут свободно переходить по этим межклеточным соединениям. Поддержание такой ионной связи между клетками зависит от энергии, получаемой благодаря окислительному фосфорилированию. Синаптический контакт (синапсы) Этот тип контактов характерен для нервной ткани и встречается как между двумя нейронами так и между нейроном и каким-либо иным элементом - рецептором или эффектором (например, нервно-мышечное окончание). Синапсы — участки контактов двух клеток, специализированных для односторонней передачи возбуждения или торможения от одного элемента к другому.
Типы синапсов: 1- пресинаптическая мембрана (мембрана отростка нервной клетки); 2 – постсинаптическая мембрана; 3 – синаптическая щель; 4 – синаптические пузырьки; 5 - митохондрии
В принципе подобного рода функциональная нагрузка, передача импульса может осуществляться и другими типами контактов (например, щелевидным контактом в сердечной мышце) однако в синаптической связи достигается высокая эффективность и подвижность реализации импульса. Синапсы образуются на отростках нервных клеток – это терминальные участки дендритов и аксонов. Межнейронные синапсы обычно имеют вид грушевидных расширений, бляшек на конце отростка нервной клетки. Такое терминальное расширение отростка одной из нервных клеток может контактировать и образовывать синаптическую связь как с телом другой нервной клетки, так и с ее отростками. Периферические отростки нервных клеток (аксоны) образуют специфические контакты с клетками-эффекторами или клетками-рецепторами. Следовательно, синапс — это структура, образующаяся между участками двух клеток, так же как и десмосома Мембраны этих клеток разделены межклеточным пространством синаптической щелью шириной около 20 — 30 нм Часто в просвете этой щели виден тонковолокнистый, перпендикулярно расположенный по отношению к мембранам материал. Мембрана в области синаптического контакта одной клетки называется пресинаптической, другой, воспринимающей импульс, — постсинаптической. В электронном микроскопе обе мембраны выглядят плотными, толстыми. Около пресинаптической мембраны выявляется огромное количество мелких вакуолей, синаптических пузырьков, заполненных медиаторами. Синаптические пузырьки в момент прохождения нервного импульса выбрасывают свое содержимое в синаптическую щель. Постсинаптическая мембрана часто выглядит толще обычных мембран из-за скопления около нее со стороны цитоплазмы множества тонких фибрилл. Синаптические нервные окончания удается выделить при фракционировании клеточных компонентов нервной ткани. При этом оказывается, что структура синапса очень устойчива: после разрушения клеток участки контактов отростков двух соседних клеток отрываются, но не разъединяются. Тем самым можно считать, что синапсы помимо функции передачи нервного возбуждения обеспечивают жесткое соединение поверхностей двух взаимодействующих клеток. Плазмодесмы. Этот тип межклеточных связей встречается у растений. Плазмодесмы представляют собой тонкие трубчатые цитоплазматические каналы, соединяющие две соседние клетки. Диаметр этих каналов обычно составляет 40—50 нм. Ограничивающая эти каналы мембрана непосредственно переходит в плазматические мембраны соседствующих клеток. Плазмодесмы проходят сквозь клеточную стенку, разделяющую клетку. Таким образом, у некоторых растительных клеток плазмодесмы соединяют гиалоплазму соседних клеток, поэтому формально здесь нет полного разграничения, отделения тела одной клетки от другой, это скорее представляет из себя синцитий' объединение многих клеточных территорий с помощью цитоплазматических мостиков. Внутрь плазмодесм могут проникать мембранные трубчатые элементы, соединяющие цистерны эндоплазматического ретикулума сосед них клеток. Образуются плазмодесмы во время деления клетки, когда строится первичная клеточная оболочка. У только что разделившихся клеток число плазмодёсм может быть очень велико (до 1000 на клетку), при старении клеток их число падает за счет разрывов при увеличении толщины клеточной стенки. Функциональная роль плазмодёсм очень велика, с их помощью обеспечивается межклеточная циркуляция растворов, содержащих питательные вещества, ионы и другие соединения. По плазмодесмам могут перемещаться липидные капли. Через плазмодесмы происходит заражение клеток растительными вирусами. Дата добавления: 2015-12-15 | Просмотры: 1109 | Нарушение авторских прав |
 Плазматическая мембрана, как уже говорилось, принимает активное участие в межклеточных контактах, связанных с конъюгацией одноклеточных организмов. У многоклеточных организмов за счет межклеточных взаимодействий образуются сложные клеточные ансамбли, поддержание которых может осуществляться разными путями. В зародышевых, эмбриональных тканях, особенно на ранних стадиях развития, клетки остаются в связи друг с другом за счет способности их поверхностей слипаться. Это свойство адгезии (соединения, контактирования) клеток может определяться свойствами их поверхности, которые специфически взаимодействуют друг с другом. Механизм этих связей еще недостаточно изучен, но вероятнее всего, что он обеспечивается взаимодействием между липопротеидами и гликокаликсом плазматических мембран. При таком межклеточном взаимодействии эмбриональных клеток между плазматическими мембранами всегда остается щель шириной около 20 нм, заполненная гликокаликсом. Обработка ткани ферментами, нарушающими целостность гликокаликса (муказы, действующие гидролитически на муцины, мукополисахариды) или повреждающими плазматическую мембрану (протеазы), приводит к обособлению клеток друг от друга, к их диссоциации. Однако если удалить фактор диссоциации, то клетки могут снова собираться, реагрегировать. Так можно диссоциировать клетки разных по окраске губок, оранжевых и желтых. Оказалось, что в смеси этих клеток образуются два типа агрегатов: состоящие только из желтых и только из оранжевых клеток. При этом смешанные клеточные суспензии самоорганизуются, восстанавливая исходную многоклеточную структуру. Сходные результаты были получены с суспензиями разделенных клеток эмбрионов амфибий; в этом случае происходит избирательное пространственное обособление клеток эктодермы от энтодермы и от мезенхимы. Более того, если для реагрегации используются ткани поздних стадий развития зародышей, то в пробирке самостоятельно собираются различные клеточные ансамбли, обладающие тканевой и органной специфичностью, образуются эпителиальные агрегаты, сходные с почечными канальцами и т. д.
Плазматическая мембрана, как уже говорилось, принимает активное участие в межклеточных контактах, связанных с конъюгацией одноклеточных организмов. У многоклеточных организмов за счет межклеточных взаимодействий образуются сложные клеточные ансамбли, поддержание которых может осуществляться разными путями. В зародышевых, эмбриональных тканях, особенно на ранних стадиях развития, клетки остаются в связи друг с другом за счет способности их поверхностей слипаться. Это свойство адгезии (соединения, контактирования) клеток может определяться свойствами их поверхности, которые специфически взаимодействуют друг с другом. Механизм этих связей еще недостаточно изучен, но вероятнее всего, что он обеспечивается взаимодействием между липопротеидами и гликокаликсом плазматических мембран. При таком межклеточном взаимодействии эмбриональных клеток между плазматическими мембранами всегда остается щель шириной около 20 нм, заполненная гликокаликсом. Обработка ткани ферментами, нарушающими целостность гликокаликса (муказы, действующие гидролитически на муцины, мукополисахариды) или повреждающими плазматическую мембрану (протеазы), приводит к обособлению клеток друг от друга, к их диссоциации. Однако если удалить фактор диссоциации, то клетки могут снова собираться, реагрегировать. Так можно диссоциировать клетки разных по окраске губок, оранжевых и желтых. Оказалось, что в смеси этих клеток образуются два типа агрегатов: состоящие только из желтых и только из оранжевых клеток. При этом смешанные клеточные суспензии самоорганизуются, восстанавливая исходную многоклеточную структуру. Сходные результаты были получены с суспензиями разделенных клеток эмбрионов амфибий; в этом случае происходит избирательное пространственное обособление клеток эктодермы от энтодермы и от мезенхимы. Более того, если для реагрегации используются ткани поздних стадий развития зародышей, то в пробирке самостоятельно собираются различные клеточные ансамбли, обладающие тканевой и органной специфичностью, образуются эпителиальные агрегаты, сходные с почечными канальцами и т. д.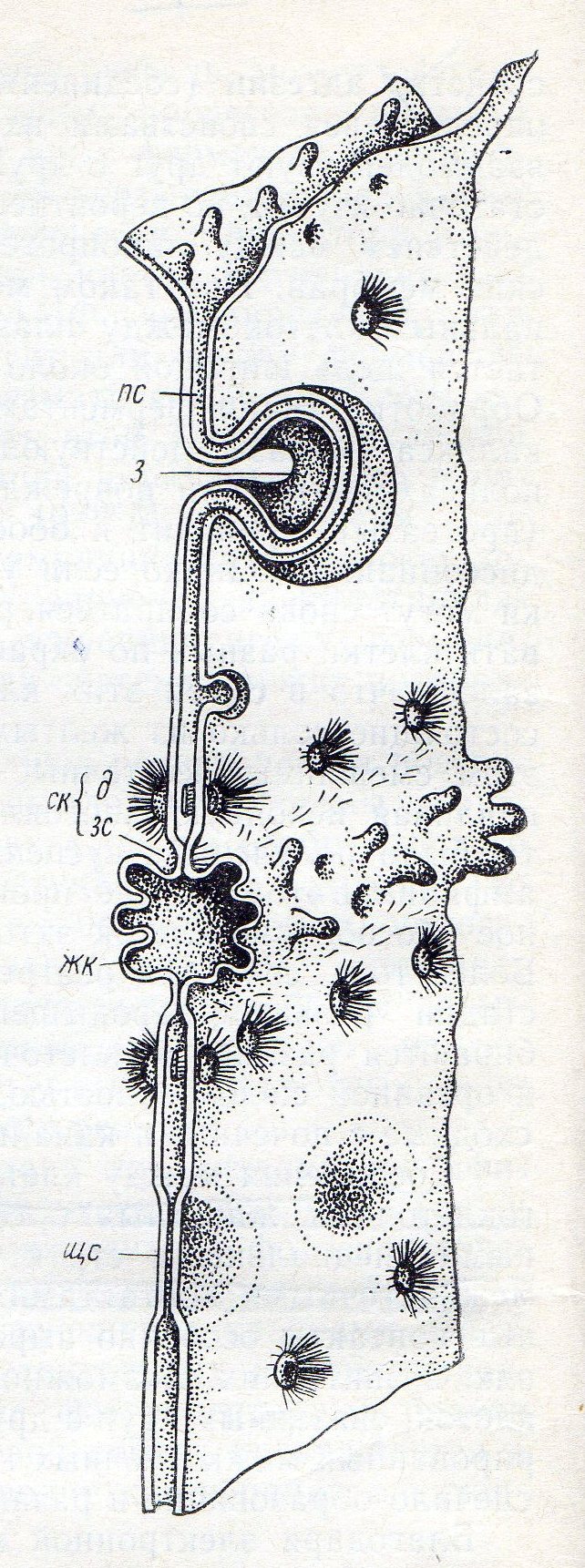
 разрушается протеиназами и исчезает после удаления кальция. Со стороны цитоплазмы в этом месте видно скопление тонких микрофибрилл 4—7 нм толщиной, располагающихся в виде сети на глубину до 0,3—0,5 мкм, что создает высокую электронную плотность всей структуры, которая сразу же бросается в глаза при изучении таких контактов в электронном микроскопе. Существует несколько типов этого контакта. Один из них, зона слипания, образует поясок, или ленту, вокруг клетки. Часто такой поясок идет сразу же за зоной плотного контакта. Часто встречается, особенно в покровном эпителии, так называемая десмосома. Последняя представляет собой небольшую площадку диаметром до 0,5 мкм, где между мембранами располагается область с высокой электронной плотностью, иногда имеющая слоистый вид. К плазматической мембране в зоне десмосомы со стороны" цитоплазмы прилегает участок электронноплотного вещества, так что внутренний слой мембраны кажется утолщенным. Под утолщением находится область тонких фибрилл, которые могут быть погружены в относительно плотный матрикс. Эти фибриллы (в случае покровного эпителия тонофибриллы) часто образуют петли и возвращаются в цитоплазму. В целом области десмосомы видны в электронном микроскопе как темные пятна, симметрично расположенные на плазматических мембранах соседних клеток. Десмосомы удалось выделить в виде отдельной фракции из покровного эпителия.
разрушается протеиназами и исчезает после удаления кальция. Со стороны цитоплазмы в этом месте видно скопление тонких микрофибрилл 4—7 нм толщиной, располагающихся в виде сети на глубину до 0,3—0,5 мкм, что создает высокую электронную плотность всей структуры, которая сразу же бросается в глаза при изучении таких контактов в электронном микроскопе. Существует несколько типов этого контакта. Один из них, зона слипания, образует поясок, или ленту, вокруг клетки. Часто такой поясок идет сразу же за зоной плотного контакта. Часто встречается, особенно в покровном эпителии, так называемая десмосома. Последняя представляет собой небольшую площадку диаметром до 0,5 мкм, где между мембранами располагается область с высокой электронной плотностью, иногда имеющая слоистый вид. К плазматической мембране в зоне десмосомы со стороны" цитоплазмы прилегает участок электронноплотного вещества, так что внутренний слой мембраны кажется утолщенным. Под утолщением находится область тонких фибрилл, которые могут быть погружены в относительно плотный матрикс. Эти фибриллы (в случае покровного эпителия тонофибриллы) часто образуют петли и возвращаются в цитоплазму. В целом области десмосомы видны в электронном микроскопе как темные пятна, симметрично расположенные на плазматических мембранах соседних клеток. Десмосомы удалось выделить в виде отдельной фракции из покровного эпителия.