|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Кора головного мозку: зони- сенсорна,асоціативна,рухова.Пасивний транспорт
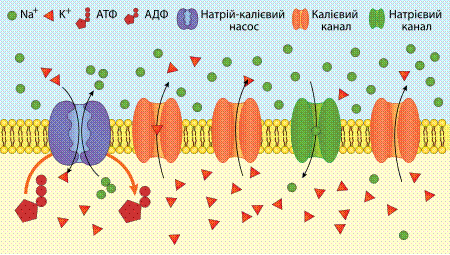
Пасивний транспорт відбувається мимовільно без затрати енергії шляхом дифузії, осмосу та полегшеної дифузії. Дифузія — це транспорт молекул та іонів через мембрану з ділянки з високою концентрацією до ділянки з низькою концентрацією, тобто речовини надходять за градієнтом концентрації. Дифузія може бути простою та полегшеною. Якщо речовини добре розчинні в ліпідах, то вони проникають до клітини шляхом простої дифузії. Напр. оксиген, потрібний клітині при диханні, та вуглекислий газ у розчині швидко дифундують крізь мембрани. Таким способом проникають до клітини також деякі фармацевтичні препарати, які є ліпідорозчинними. Вода також здатна проходити крізь мембранні пори, що утворені білками, і переносити молекули та іони речовин, які в ній розчинені. Дифузію води крізь напівпроникну мембрану називають осмосом. Вода переходить з ділянки з низькою концентрацією солей до ділянки, де їхня концентрація вища. Тиск на мембрану, що виникає при цьому, називають осмотичним. Усі живі клітини здатні регулювати осмотичний тиск, змінюючи концентрацію речовин поза клітиною та всередині клітини. Речовини, які не є розчинними у ліпідах, транспортуються через іонні канали, утворені в мембрані білками, чи за допомогою білків-переносників. Це полегшена дифузія, шляхом якої, напр. здійснюється надходження глюкози до еритроцитів. Серед систем пасивного транспорту важливу роль відіграють іонні канали, які забезпечують проникність мембрани для Na+, K+, Ca2+. Na+-канали активуються вератрадином, батрахотоксином, блокуються амілоридом, тріамтереном; K+-канали блокуються місцевими анестетиками (лідокаїном, дикаїном), деякими протисудомними (дифеніл, карбамазепін, вальпроати, фенобарбітал та ін.) і протиаритмічними засобами (аміодарон); Ca2+-канали чутливі до цілої низки хімічних речовин, зокрема верапамілу, дилтіазему, ніфедипіну та інших похідних дигідропіридинів. Трансмембранний обмін (антипорт) чи односпрямований транспорт (симпорт) іонів здійснюється спеціальними білками-переносниками. Система односпрямованого транспорту (котранспорту) представлена (Na++K++Cl–)-переносником, що є чутливим до дії діуретиків (фуросеміду, амілориду, туметаніду). Виключення або різка зміна властивостей переносників і каналів лежить в основі дії багатьох токсичних і фармацевтичних речовин. Деякі речовини — іонофори, до яких належать різні антибіотики (валіноміцин, амфотерицин В, нонактин, енніатини, аламетицин та ін.), синтетичні циклополіефіри самостійно здатні утворювати канали у ліпідному бішарі мембрани. Дія деяких ЛП заснована на зміні властивостей каналів і переносників, що дозволяє регулювати транспорт речовин у клітинах і організмі в цілому. Активний транспорт речовин крізь мембрану здійснюється проти градієнта їхньої концентрації із затратою енергії АТФ та за участю спеціальних мембранних білків — транспортних АТФаз, які також називаються іонними насосами. Найбільш поширеними в клітині тварин є Н+-АТФаза, Na+,K+-АТФаза і Са2+-АТФаза, що являють собою цілі мембранні комплекси із складною структурою. Функціональне значення біологічних насосів полягає у підтримці всередині клітини постійного іонного складу. Na+,K+-АТФаза сприяє виведенню Na+ з клітини та надходженню К+ до клітини за допомогою енергії АТФ і є прикладом антипортного транспорту. Із впливом на натрієвий насос пов’язаний механізм дії деяких фармацевтичних препаратів. Так, напр. серцеві глікозиди (дигоксин, уабаїн, строфантин К) пригнічують Na+,K+-АТФазу; деякі діуретики (тіазиди) інгібують активний транспорт Na+ та/або Cl– в епітелії канальців нирок; омепразол знижує кислотність шлункового соку, незворотно пригнічуючи протонний насос Н+-АТФазу парієтальних клітин шлунка. Са2+-АТФаза високочутлива до дії різноманітних тіолових отрут та ін. Крім вищенаведених видів активного транспорту, виділяють специфічні механізми переміщення речовин, пов’язані з порушенням цілісності мембрани, ендоцитоз та екзоцитоз. При ендоцитозі плазматична мембрана утворює вирости, які потім перетворюються на внутрішньоклітинні пухирці, що містять захоплений клітиною матеріал. Ці процеси відбуваються із витратою енергії АТФ. Розрізняють два види ендоцитозу: фагоцитоз і піноцитоз. Фагоцитоз (грец. phagos — пожирати, cytos — клітина) — це захоплення і поглинання клітиною великих часток (іноді цілих клітин та їхніх частин). Ендоцитоз рідини та розчинених в ній речовин називається піноцитозом (грец. pyno — пити, cytos — клітина). Шляхом ендоцитозу, напр. відбувається всмоктування жиру клітинами кишкового епітелію. Екзоцитоз — це процес виведення з клітини різноманітних речовин крізь мембрану, фактично зворотний ендоцитозу механізм. Шляхом екзоцитозу вивільнюються гормони, жирові краплини, а також медіатори в синапсах при збудженні.
Мембранний потенціал спокою[ред.] Матеріал з Вікіпедії — вільної енциклопедії.
Участь натрій-калієвої АТФази, калієвих та натрієвих каналів у формуванні мембранного потенціалу спокою Мембра́нний потенціа́л споко́ю (МПС) — це різниця потенціалів між зовнішньою та внутрішньою сторонамимембрани в умовах, коликлітина не збуджена.Цитоплазмаклітини заряджена негативно по відношенню до позаклітинної рідини через нерівномірний розподіл аніонів та катіонів по дві сторони мембрани. Різниця потенціалів (напруга) для різних клітин має значення від −50 до −200 мВ (мінус означає, що всередині клітина більш негативно заряджена ніж зовні). Мембранний потенціал спокою виникає на мембранах всіх клітин і збудливих (нервів, м'язів, секреторних клітин) і незбудливих. МПС необхідний для підтримання збудливості таких клітин, як м'язові та нервовоі. Також він впливає на транспорт всіх заряджених частинок у будь-якому типі клітин: він сприяє пасивному транспорту аніонів із клітини та катіонів у клітину. Утворення та підтримання мембранного потенціалу забезпечують різні типи іонних насосів(зокрема натрій-калієвий насос або натрій-калієва АТФаза) та іонних каналів (калієві, натрієві, хлорні іонні канали).
Реєстрація потенціалу спокою [ред.]
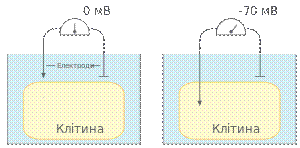
Реєстрація мембранного потенціалу спокою. На рисунку зліва мікроелектрод не введений у клітину і різниця потенціалів рівна нулю. При введенні електродав клітину реєструється напруга у приблизно −70 мВ Для реєстрації потенціалу спокою використовують спеціальну мікроелектродну техніку. Мікроелектрод — це тоненька скляна трубочка, із витягнутим кінцем, діаметром менше 1 мкм, заповнена розчином електроліту(найчастіше хлориду калію). Рефернтнимелектродом слугує срібна хлорована пластинка, розміщена у позаклітинному просторі, обидва електроди під'єднані доосцилографа. Спочатку обидва електроди занходяться у позаклітинному просторі і різниця потенціалів між ними відсутня, якщо ввести реєструючий мікроелектрод через мембрану у клітину, то осцилограф покаже стрибкоподібне зміщення потенціалу приблизно до −80 мВ. Цей зсув потенціалу називають мембранним потенціалом спокою.[1]. Формування потенціалу спокою [ред.]
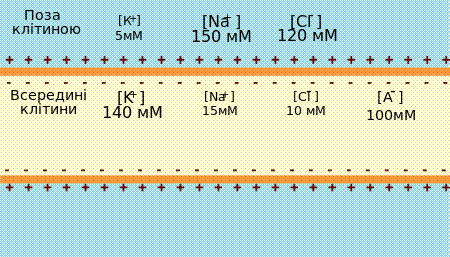
Концентрації іонів K+, Na+, Cl- та органічних аніонів всередині та зовні клітини До виникнення мембранного потенціалу спокою призводять два фактори: по-перше,концентраціїрізних іонів відрізняються зовні та всереднині клітини, по-другемембрана є напівпроникною: одні іони можуть через неї проникати, інші — ні. Обидва ці явища залежать від наявності у мембрані спеціальних білків: концентраційні градієнти створюють іонні насоси, а проникність мембрани для іонів забезпечують іонні канали. Найважливішу роль у формуванні мембранного потенціалу відіграють іони калію, натрію тахлору. Концентрації цих іонів відрізняюються по дві сторони мембрани. Для нейрона ссавцівконцентрація K+ становить 140 мМ всередині клітини і тільки 5мМ ззовні, градієнт концентрації Na+ майже протилежний — 150 мМ зовні та 15 мМ всередині. Такий розподіл іонів підтримуєтьсянатрій-калієвим насосом у плазматичній мембрані — білком що використовує енергію АТФ для закачування K+ у клітину і викачування Na+ із неї. Також існує концентраційний градієнт і для інших іонів, наприклад, хлорид аніону Cl-.[2][3] Концентраційні градієнти катіонів калію та натрію — це хімічна форма потенційної енергії. У перетворенні цієї енергії в електричну беруть участь іонні канали — пори, що формуються скупченнями спеціальних трансмембранних білків. Коли іони дифундують крізь канал, вони переносять одиницю електричного заряду. Будь-який сумарний рух позитивних або негативних іонів через мембрану буде створювати напругу, або різницю потенціалів по дві сторони мембрани. Іонні канали, що беруть участь в утовренні МПС мають вибіркову проникність, тобто дають можливість проникати тільки певному типу іонів. В мембрані нейрона у стані спокою відкритікалієві канали (ті, що в основному пропускають тільки калій), більшість натрієвих каналів — закриті. Дифузія іонів K+ через калієві канали є вирішальною для створення мембранного потенціалу. Через те, що концентрація K+ значно вища всередині клітини, хімічний градієнт сприяє відтоку цих катіонів з клітини, тому в цитоплазмі починають переважати аніони, які не можуть проходити через калієві канали. Відтік іонів калію із клітини обмежений самим мембранним потенціалом, оскільки при певному його рівні накопичення негативних зарядів у цитоплазмі буде обмежувати рух катіонів поза межі клітини. Таким чином, головним факором у виникненні МПС є розподіл іонів калію під дією електричного та хімічного потенціалів [3]. Рівноважний потенціал [ред.]
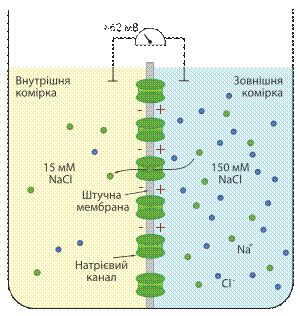
Встановлення рівноважного кальцієвого потенціалу
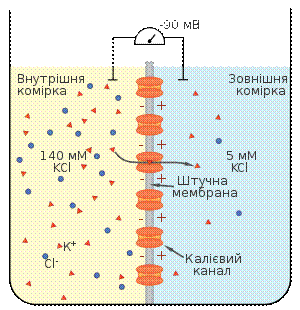
Встановлення рівноважного натрієвого потенціалу Для того, щоб визначити вплив руху певного іону через напівпроникну мембрану на формування мембранного потенціалу, будують модельні системи. Така модельна система складається із посудини поділеної на дві комірки шутчною напівпроникною мембраною, в яку вбудовані іонні канали. У кожну комірку можна занурити електрод і поміряти різницю потенціалів. Розглянемо випадок, коли штучна мебмрана проникна тільки для калію. По дві сторони мембрани модельної системи свторюють градієнт концентрації аналогічний до такого у нейроні: у комірку, що відповідає цитоплазмі (внутрішня комірка), поміщають 140 мМ розчинхлориду калію (KCl), у комірку, що відповідає міжклітинній рідині (зовнішня комірка) — 5 мМ розчин KCl. Іони калію будуть дифундувати через мембрану у зовнішню комірку по градієнту концентрації. Та, оскільки аніони Cl-проникати через мембрану не можуть у внутрішній комірці виникатиме надлишок негативного заряду, який перешкоджатиме подальому відтоку катіонів. Коли такі модельні нейрони досягнуть станурівноваги, дія хімічного та електричного потенціалу буде збалансована, жодної сумарної дифузії К+ не буде спостерігатись. Значення мембранного потенціалу, що виинкає за таких умов, називається рівноважним потенціалом для певного іона (Еіон). Рівноважний потенціал для калію становить приблизно −90 мВ. Аналогічний дослід можна провести і длянатрію, встановивши між комірками мембрану проникну тільки для цього катіона, і помістивши у зовнішню комірку розчин хлориду натрію із концентрацією 150 мМ, а у внутрішню — 15 мМ. Натрій буде рухатись у внутрішню комірку, рівоноважний потенціал для нього становитиме приблизно +62 мВ. Кількість іонів, що повинна дифундувати для генерації електричного потенціалу дуже невелика (приблизно 10−12 моль К+ на 1 см 2 мембрани), цей факт має два важливі наслідки. По-преше, це означає, що концентрації іонів, які можуть проникати через мембрану, залишаються стабільними зовні і всередині клітини, навіть після того як їхній рух забезпечив утоврення електричного потенціалу. По-друге, мізерні потоки іонів через мембрану, потірбні для встановлення потенціалу, не порушують електронейтральності цитоплазми та позаклітинної рідини в цілому, розподіл зарядів відбувається тільки в ділянці, що безпосередньо прилягає до плазматичної мембрани.[4] Рівняння Нернста [ред.] Рівноважний потенціал для певного іона, наприклад для калію, можна обрахувати за рівнянням Нернста, що має наступний вигляд:
де R — універсальна газова стала, Т — абслоютна температура (по шкалі Кельвіна), z — заряд іона, F — число Фарадея, [K+]o, [K+]i — концентрація калію зовні та всередині клітини відповідно. Оскільки описані процеси відбуваються за температури тіла — 310° К, а десятковимилогарифмами в обчисленні користуватись легше ніж натуральними, це рівняння перетворюють наступним чином:
Підставляючи концентрації К+ у рівняння Нернста отримуємо рівноважний потенціал для калію, що становить −90 мВ. Оскільки за нульовий потенціал приймається зовнішня сторона мембрани, то знак мінус означає, що за умов рівноважного калієвого потенціалу внутрішня сторна мембрани порівняно більш електронегативна. Аналогічні обрахунки можна провести і для рівноважного натієвого потенціалу, він становить +62 мВ. Рівняння Голдмана [ред.] Хоча рівноважний потенціал для іонів калію становить −90 мВ, МПС нейрона дещо менш негативний. Ця різниця відображає незначне але постійне проходження іонів Na+ черезмембрану у стані спокою. Оскільки концентраційний градієнт для натрію протилежний до такого для калію, Na+ рухається всередину клітини і зсуває сумарний заряд на внутрішній стороні мембрани у позитивну сторону. Насправді МПС нейрона становить від −60 до −80 мВ. Це значення значно ближче до ЕK ніж до ЕNa, тому що у стані спокою в нейроні відкрито багато калієвих каналів і дуже мало натрієвих. Також на встанвлення МПС впливає рух іонів хлору. У 1943 році Девід Голдаман запропонував вдосконалити рівняння Нернста так, щоб воно відображало вплив різних іонів на мембарнний потенціал, в цьому рівнянні враховується відносна проникність мембрани для кожного типу іонів:
де R — універсальна газова стала, Т — абслоютна температура (по шкалі Кельвіна), z — заряд іона, F — число Фарадея, [іон]o, [іон]i — концентрації іонів зовні та всередині клітин, Р — відносна проникність мембрани для відповідного іона. Значення заряду в даному рівнянні не записується, але воно враховане — для хлору зовнішня і внутрішня концентрація поміняні місцями, так як його заряд −1. Значення мембранного потенціалу спокою для різних тканин [ред.] · Посмуговані м'язи −95 мВ; · Непосмуговані м'язи −50 мВ; · Астроглія від −80 до −90 мВ; · Нейрони −70 мВ. Роль натрій-калієвого насосу у формуванні МПС [ред.] Мембранний потенціал спокою може існувати тільки за умови нерівномірного розподілу іонів, що забезпечується функціонуванням натрій-калієвого насосу. Окрім того, цей білок робить також має електрогенні властовості — він переносить 3 катіони натрію в обмін на 2 іони калію, що рухаються всередину клітини. Таким чином, Na+-K+-АТФаза знижує МПС на 5-10 мВ. Пригнічення діяльності цього білка призводить до незначного (на 5-10 мВ) миттєвого підвищення мембранного потенціалу, після чого він ще деякий час існуватиме на досить стабільному рівні, поки будуть зберігатись градієнти концентрацій Na+ та K+. Згодом ці градієнти почнуть зменшуватись, внаслідок проникнсоті мембрани до іонів, і через кілька десятків хвилин електричний потенціал на мембрані зникне [2].
Екзоцитоз— це система, яка забезпечує переміщення іонів натрію і калію через плазмолему. Перенесення іонів здійснює спеціальний фермент-переносник Na+-K+-АТФ-аза, який використовує енергію клітини і випомповує іони натрію через плазмолему назовні, одночасно він захоплює іони калію ззовні і звільнює їх усередині клітини. Таким чином підтримується різниця концентрації натрію і калію: концентрація натрію вища у тканинній рідині (зовні плазмолеми), а концентрація калію — в цитоплазмі. При гідролізі однієї молекули АТФ з клітини випомповуються 3 іони Na+ і вводяться в неї 2 іони K+. Водночас білок- переносник іону транспортує глюкозу в клітину. Натрієво-калієва помпа забезпечує підтримку постійного об’єму клітини (шляхом регулювання осмотичного тиску), а також мембранного потенціалу. Транспорт у мембранному упакуванні характерний тим, що речовини, які переносяться через плазмолему, зазвичай оточені мембраною. (Так, транспорт речовин у клітину називають ендоцитозом, а з клітини — екзоцитозом. Крапля, що виділяється, може бути оточена мембраною (наприклад, ліпосома) або ні. При ендоцитозі (від грецьк. endo — всередину і cytos — клітина) матеріал, який знаходиться в позаклітинному просторі, охоплени складками плазмолеми, потрапляє в клітину у вигляді ендоцитозного міхурця, або ендосоми. Потім вміст ендосоми піддається процесингу (внутрішньоклітинній обробці). Різновидами ендоцитозу є фагоцитоз (від грецьк. phagein — поїдати і cytos — клітина) — захоплення і поглинання великих (до 1 мкм) і щільних частинок і піноцитоз (від грецьк. pinein — пити і cytos — клітина) — втягування оточених плазмолемою рідин і розчинних речовин. Рецепторно-опосередкований ендоцитоз значно ефективніший, оскільки він опосередкований рецепторами, які зв’язуються з молекулами фагоцитованого об’єкта — лігандами (від латин. ligare — зв’язувати),.. Після поглинання речовини комплекс рецептор-ліганд розпадається, і рецептори можуть знов повертатися в плазмолему. Прикладом такого рецепторно-опосередкованого ендоцитозу може служити фагоцитоз лейкоцитом мікроба. На плазмолемі лейкоцита є рецептори до іму- ноглобулінів (антитіла), і якщо поверхня мікробів вкрита антитілами- Екзоцитоз— це система, яка забезпечує переміщення іонів натрію і калію через плазмолему. Перенесення іонів здійснює спеціальний фермент-переносник №+-К+-АТФ-аза, який використовує енергію клітини і випомповує іони натрію через плазмолему назовні, одночасно він захоплює іони калію ззовні і звільнює їх усередині клітини. Таким чином підтримується різниця концентрації натрію і калію: концентрація натрію вища у тканинній рідині (зовні плазмолеми), а концентрація калію — в цитоплазмі. При гідролізі однієї молекули АТФ з клітини випомповуються 3 іони і вводяться в неї 2 іони К+. Водночас білок- переносник іону транспортує глюкозу в клітину. Натрієво-калієва помпа забезпечує підтримку постійного об’єму клітини (шляхом регулювання осмотичного тиску), а також мембранного потенціалу. Транспорт у мембранному упакуванні характерний тим, що речовини, які переносяться через плазмолему, зазвичай оточені мембраною. (Так, транспорт речовин у клітину називають ендоцитозом, а з клітини — екзоцитозом. Крапля, що виділяється, може бути оточена мембраною (наприклад, ліпосома) або ні. При ендоцитозі (від грецьк. епсіо — всередину і суІ05 — клітина) матеріал, який знаходиться в позаклітинному просторі, охоплени складками плазмолеми, потрапляє в клітину у вигляді ендоцитозного міхурця, або ендосоми. Потім вміст ендосоми піддається процесингу (внутрішньоклітинній обробці). Різновидами ендоцитозу є фагоцитоз (від грецьк. рНадеіп — поїдати і суй>5 — клітина) — захоплення і поглинання великих (до 1 мкм) і щільних частинок і піноцитоз (від грецьк. ріпеіп — пити і суіоз — клітина) — втягування оточених плазмолемою рідин і розчинних речовин. Рецепторно-опосередкований ендоцитоз значно ефективніший, оскільки він опосередкований рецепторами, які зв’язуються з молекулами фагоцитованого об’єкта — лігандами (від латин. Іідаге — зв’язувати),.. Після поглинання речовини комплекс рецептор-ліганд розпадається, і рецептори можуть знов повертатися в плазмолему. Прикладом такого рецепторно-опосередкованого ендоцитозу може служити фагоцитоз лейкоцитом мікроба. На плазмолемі лейкоцита є рецептори до іму- ноглобулінів (антил’ла), і якщо поверхня мікробів вкрита антитілами-опсонінами (від грецьк. opson — приправа), швидкість фагоцитозу різко зростає. Особливого посилення набуває ендоцитм в облямованих міхурцях, де в місцях їх формування нагромаджуються рецептори і особливі білки — клатрини, які зв’язують ліганди. Коли міхурець втягується в цитоплазму, його вміст дуже швидко піддається г.роце- сингу — протеолізу. Якщо клатринова оболонка не втрачається, вміст міхурця залишається незмінним. Екзоцитоз (від грецьк. exo — назовні і cytos — клітина) — процес, при якому мембранні екзоцитозні міхурці, що сформувалися в цитоплазмі, наближаються до плазмолеми і зливаються з нею своєю мембраною, яка вбудовується в плазмолему. При цьому вміст міхурця виділяється в позаклітинний простір. Трансцитоз (від латин. trans — через і cytos — клітина) — процес, при якому на одній поверхні клітини формується ендоцитозний міхурець, який переноситься на протилежну поверхню клітини як ек- зоцитозний і виділяє свій вміст в позаклітинний простір. Трансцитоз характерний для клітин з периферичною тонкою плазмолемою (ендо- теліоцити), які вистеляють дрібні кровоносні судини. У цих клітинах міхурці, зливаючись, можуть утворювати.тимчасові трансцелюлярні канали, через які транспортуються водорозчинні речовини. У процесах ендоцитозу і екзоцитозу важливу роль відіграють особливі фузогенні (від латин. fusio — злиття) мембранні білки, які концентруються в ділянках інвагінації (впинання) плазмолеми. При ендоцитозі вони сприяють змиканню складок плазмолеми, а під час екзоцитозу забезпечують вбудовування мембран міхурця в плазмолему Збалансованість процесів ендоцитозу і екзоцитозу досягається таким чином, що при ендоцитозі міхурці формуютьсятплазмолеми і її поверхня повинна зменшуватися, тоді як при екзоцитозі мембрани міхурців вбудовуються в плазмолему і збільшують її площу. Однак активні процеси екзо- і ендоцитозу не впливають суттєво на площу поверхні плазмолеми у зв’язку з урівноваженістю цих процесів. Постійний кругообіг мембран в клітині отримав назву «мембранного конвеєра».
Таламус Матеріал з Вікіпедії — вільної енциклопедії.
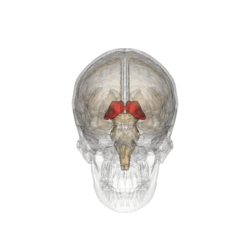
Таламус в проекції черепа
Таламус або талямус (лат. thalamus dorsalis, відгрец. θάλαμος — «кімната») — парна симетрична область головного мозку хребетних тварин, головна частина проміжного мозку. Таламус складається із системи мієлінізованих ламел, що розділяють ядра таламуса — кластери тіл нейронів. Часто в складі таламуса виділяють аллоталамус (10% об'єму) і ізоталамус. Таламус перш за все відповідає за перерозподіл інформації, що поступає від органів чуття, за виключенням нюху, до кори головного мозку. Коли інформація потрапляє на ядра таламуса, там відбувається її первинна обробка. Вважається, що таламус активно залучений у процес пам'яті. Пошкождення таламуса може привести доантероградної амнезії або викликати тремор — мимовільне тремтіння кінцівок у стані спокою. Таламус утворює бічні сторони шлуночка. Він майже повністю сформований із сірої речовини. У ньому налічують майже 40 ядер.
Гіпоталамус Матеріал з Вікіпедії — вільної енциклопедії.
Гіпотала́мус (гіп… + грец. thalamos — кімната) — відділ проміжного мозку, що виконує функцію мозкового контролю над внутрішніми органами. Є вищим центром регуляції вегетативних функцій організму, місцем взаємодії нервової та ендокринної систем. Гіпоталамус розглядається як центр, що інтегрує діяльність вісцеральної системи, іннервує органи й тканини, а також відповідає за сталість внутрішнього середовища. Його головною функцією є підтримка гомеостазу в організмі. Гіпоталамус повинен відповідати на багато різноманітних сигналів, які мають внутрішнє або зовнішнє походження. Тому він пов'язаний з багатьма відділами ЦНС, включаючи ретикулярну формацію стовбуру головного мозку та автономні зони, лімбічну долю переднього мозку(зокрема, мозочковий мигдалик, перегородка, нюхова цибулина, кора головного мозку). Гіпоталамус чутливий до таких факторів як: · Світло: тривалість дня та фотоперіод для регуляції циркадного та сезонного ритмів; · Нюхові стимули, включно з феромонами; · Стероїди, включаючи годанальні стероїди та кортикостероїди; · Нервові імпульси, що надходять, зокрема від серця, шлунку та від репродуктивних органів; · Автономне подразнення; · Стимули, які надходять з кров'ю, включаючи лептин, ґрелін, ангіотензин, інсулін, слизові гормони, цитокіни, плазмову концентрацію глюкози та осмотичну концентрацію розчину тощо; · Стрес; · Мікроорганізми, що призводять до підвищення температури тіла. В свою чергу гіпоталамус впливає на · проекції нервів та · гормони ендокринної системи. Діяльність гіпоталамуса регулює гіпофіз. Гіпоталамус і гіпофіз утворюють єдину гіпоталамо-гіпофізарну систему — типовий приклад тісного взаємозв'язку нервового і гуморальногоспособів регуляції функцій організму.
Базальні ядра, їх функції, симптоми ураження. Базальні ядра знаходяться в глибині кінцевого мозку. До них відносяться: - хвостате ядро; - шкаралупа (разом з попереднім утворює смугасте тіло);- бліда куля. Як єдине ціле з базальними ядрами функціонують чорна субстанція тасубталамічне ядро. Ці ядра об’єднані між собою двосторонніми зв’язками, отримуютьінформацію від кори (асоціативних та рухових зон) та мозочка. Післявідповідної обробки інформація від базальних ядер передається: - через моторні (передні) ядра таламуса до рухової кори; - через стовбурові рухові ядра (червоне ядро, вестибулярне ядро,ретикулярна формація) до мотонейронів спинного мозку (до м’язів. В діяльності базальних ядер важливу роль відіграє рух збудження, якиймає назву циклу шкаралупи та циклу хвостатого ядра. 1. Цикл шкаралупи – забезпечує участь базальних ядер в реалізаціїпрограм складних набутих рухових реакцій. Цикл починається з передачіінформації від премоторної зони кори (ПМК) до шкаралупи (далі до блідоїкулі (БК) (через чорну субстанцію (ЧС) та субталамічне ядро (СТЯ)інформація, перероблена в базальних ядрах передається в першу моторнузону кори (МК) через моторні ядра таламуса. Цикл хвостатого ядра – забезпечує участь базальних ядер в формуванніпрограм складних рухових реакцій. При здійсненні цього циклу інформаціявід асоціативних зон кори (АЗК) про те, яким повинен бути результатрухової реакції, направляється в хвостате ядро (ХЯ) (далі в шкаралупу(Ш) і бліду кулю (БК) і далі через моторні ядра таламуса в рухову кору.Завдяки цьому рухова кора отримує готову рухову програму, яка підлягаєнаступній реалізації. Необхідно пам’ятати, що у створенні програм рухових актів окрімбазальних ядер важливу роль відіграє кора мозочка та його зубчасті ядра. Симптоми ураження базальних ядер: При ураженні смугастого тіла: гіпотонія м’язів; гіперкінези, в тому числі у спокої; При ураженні блідої кулі та чорної субстанції: гіпокінезія, рухи втрачають індивідуальне забарвлення за рахунокзникнення міміки та пантоміміки; воскова ригідність – людина може довго утримувати ненормальну (незручну)позу, положення кінцівок; тремор спокою.
Кора головного мозку: зони- сенсорна,асоціативна,рухова. Кора головного мозку: зона сенсорна Сенсорні зони – це функціональні зони кори головного мозку, які через висхідні нервові шляхи отримують сенсорну інформацію від більшості рецепторів тіла. Вони займають окремі ділянки кори, пов’язані з певними видами відчуттів. Розміри цих зон корелюють з числом рецепторів у відповідній сенсорній системі. - Первинні сенсорні зони і первинні моторні зони (проекційні зони); - Вторинні сенсорні зони і вторинні моторні зони (асоціативні одномодальних зони); - Третинні зони (асоціативні разномодальних зони); Первинні сенсорні і моторні зони займають менше 10% поверхні кори головного мозку і забезпечують найбільш прості сенсорні і рухові функції. Кора головного мозку: зони асоціативні (функціональні) Асоціативні зони – це функціональні зони кори головного мозку. Вони пов’язують знову що надходить сенсорну інформацію з полученой раніше і зберігається в блоках пам’яті, а також порівнюють між собою інформацію, що отримується від різних рецепторів. Сенсорні сигнали інтерпретуються, осмислюються і при необхідності використовуються для визначення найбільш підходящих відповідних реакцій, які вибираються в асоціативної зоні і передаються в пов’язану з нею рухову зону. Таким чином, асоціативні зони беруть участь у процесах запам’ятовування, навчання і мислення, і результати їх діяльності складають те, що зазвичай називають інтелектом. Окремі великі асоціативні області розташовані в корі поруч з відповідними сенсорними зонами. Наприклад, зорова асоціативна зона розташована в потиличній зоні безпосередньо попереду сенсорної зорової зони і здійснює описані вище асоціативні функції, пов’язані із зоровими відчуттями. Деякі асоціативні зони виконують лише обмежену спеціалізовану функцію і пов’язані з іншим асоціативними центрами, здатними піддавати інформацію подальшій обробці. Наприклад, звукова асоціативна зона аналізує звуки, розділяючи їх на категорії, а потім передає сигнали в більш спеціалізовані зони, такі як мовна асоціативна зона, де сприймається сенс почутих слів. Ці зони відносяться до асоціативної корі і беруть участь в організації когнітивних функцій і складних форм поведінки. - Вторинні сенсорні зони і вторинні моторні зони (асоціативні одномодальних зони); - Третинні зони (асоціативні разномодальних зони); - Паралімбіческіе зони і - Лимбические зони. Кора головного мозку: зона рухова Рухові зони – це функціональні зони кори головного мозку, що посилають рухові імпульси до довільних м’язам по спадним шляхах, які починаються в білій речовині великих півкуль. Багато рухові імпульси йдуть прямо в спинний мозок через два великих пірамідних тракту (кортікоспінальних тракту), що проходять в стовбурі мозку. Решта рухові імпульси передаються по екстрапірамідних шляхів, тут же йдуть рухові імпульси від базальних гангліїв і мозочка. У довгастому мозку всі шляхи перехрещуються, так що імпульси, які йдуть від кори лівої півкулі, іннервують праву половину тіла і навпаки. Тіла нейронів, що у освіті пірамідних трактів, лежать в рухових зонах кори, а їх аксони утворюють синапси безпосередньо з мотонейронами спинного мозку в тому його сегменті, де ці нейрони виходять на периферію. У головному мозку немає ніяких проміжних синапсів, тому імпульси і наступні відповіді на них по шляху не затримуються і не видозмінюються. Головним екстрапірамідних трактом є ретікулоспінальний тракт, перемикаючий імпульси від ретикулярної формації, яка лежить в стовбурі мозку між таламуса і довгастим мозком. З різних відділів головного мозку, що контролюють рухову активність, імпульси надходять у певні ділянки ретикулярної формації, де вони модифікуються під впливом імпульсів, що йдуть від кори, і стають або збудливими, або гальмівними. Наприклад, імпульси від мозочка і премоторной зони кори, керуючої координованими рухами, надходять в ту область ретикулярної формації, яка знаходиться в довгастому мозку і посилає імпульси, що стимулюють гальмівні мотонейрони. Останні придушують активацію певних м’язів, що дає можливість здійснювати складні координовані рухи тіла. Інші комбінації рухових імпульсів, навпаки, стимулюють збуджуючі нейрони, і загальний вплив ретикулярної формації на рухову активність виявляється збудливим. Більшість волокон сенсорних нейронів на своєму шляху через таламус до кори віддає колатералі (бічні гілки) у ретикулярну формацію, беручи участь в утворенні ретикулярної активує системи, яка тонізує кору і бере участь в пробудженні організму від сну. Недостатня активність цієї системи або її руйнування приводить відповідно до глибокого сну або коми. Як вважають, багато речовини, що викликають загальний наркоз, надають свою дію, тимчасово блокуючи синаптичну передачу в цій системі. Передбачається також, що ретикулярна активує система відповідальна за виникнення і підтримання спонукань до дії і концентрації уваги.
Відділ нервової системи, Що регулює основну частину вісцеральних функцій організму, називають автономної (вегетативної) нервової системою. Ця система регулює артеріальний тиск, рухову активність шлунково-кишкового тракту, спорожнення сечового міхура,потовиділення, температуру тіла і багато інших функцій, причому деякі з них регулює автономна нервова система практично повністю, а інші - лише частково. Однією з найбільш дивовижних особливостей автономної нервової системи є її здатність змінювати вісцеральні функції дуже швидко та інтенсивно, наприклад протягом 3-5 сек вдвічі змінити частоту серцевих скорочень, протягом 10-15 сек подвоїти артеріальний тиск або вінших надзвичайних ситуаціях знизити його за 10-15 сек до рівня, що може викликати непритомність. Потовиділення може початися протягом декількох секунд, а сечовий міхур мимоволі випорожниться також швидко. Автономна нервова система активується переважно центрами, локалізованими в спинному мозку, стовбурі мозку і гіпоталамусі. Крім того, частини кори великого мозку, особливо лімбічної кори, можуть посилати сигнали до нижчерозташованимицентрам і таким чином впливати на вегетативну регуляцію. Автономна нервова система часто діє за допомогою вісцеральних рефлексів. Це означає, що підсвідомі сенсорні сигнали від внутрішніх органів надходять у вегетативні ганглії, ствол мозку або гіпоталамус і потім повертаються назад, безпосередньо до внутрішніх органів, регулюючи їх активність шляхом підсвідомих рефлекторних реакцій. Еферентнісигнали до різних органів тіла передаються через два основні відділу автономної нервової системи - симпатичний і парасимпатичний, структурно-функціональні особливості яких обговорюються далі. Слід розуміти, що імеено через дані сигнальні системи здійснюється вплив при масажі тіла. Сучасні курси масажу засновані на розвантаженні нервової та м'язової системи через аферентні і еферентні нервові волокна. Слід розуміти, що вплив масажу через м'язову і суглобову нервову систему дозволяє значно поліпшити трофіку тканин за рахунок регулювання взаємодії різних відділів автономної нервової системи. На малюнку представлена основна організація периферичного відділу симпатичної нервової системи. На малюнку особливо виділені: (1) одна з двох ланцюжків паравертебральних симпатичних гангліїв, пов'язаних снінлльнимі нервами зі спинним мозком (з боку хребетного стовпа), (2) два превертебральних ганглія (черевний і подчревной), (3) нерви, що йдуть від гангліїв до різним внутрішнім органам. Симпатичні нервові волокна починаються разом зі спінальних нервами в спинному мозку між сегментами Т1 і L2 і проходять спочатку до симпатичної ланцюжку, а потім до тканин і органів, що стимулюється симпатичними нервами. Відмінність між симпатичними і скелетними руховими нервами полягає в наступному. Кожен симпатичний шлях від спинного мозку до стимулируемой тканини складається з двох нейронів - прегангліонарних і постгангліонарних, тоді як шлях рухових волокон до скелетної м'яза має лише один нейрон. Тіло клітини кожного прегангліонарних нейронів лежить в бічному розі спинного мозку, його волокно проходить, через передній корінець спинного мозку до відповідного спинальний нерв. Як тільки спинальний нерв покидає спинномозковій канал, прегангліонарних симпатичні волокна відокремлюються і проходять через білу гілку в один з гангліїв симпатичної ланцюжка. Потім можливий один із трьох варіантів ходу волокон: (1) волокно може формувати синапс з постгангліонарних симпатичним нейроном в тому ганглії, в який воно ввійшло, (2) волокно може проходити вгору і вниз і формувати синапс в одному з інших гангліїв ланцюжка, (3) волокно може проходити різні відстані по ланцюжку і потім через один з симпатичних нервів вийти з ланцюжка назовні, утворюючи в результаті синапси в периферичному симпатическом ганглії. Постгангліонарних симпатичних нейрон лежить або в одному з гангліїв симпатичної ланцюжка, або в одному з периферичних симпатичних гангліїв. Від кожного з цих джерел постгангліонарні волокна потім відправляються до місця призначення в різних органах. Симпатичні нервові волокна в нервах, Іннервують скелетні м'язи. Деякі з постгангліонарних волокон виходять з симпатичної ланцюжка і повертаються в спинальні нерви через сірі гілки на всіх рівнях спинного мозку. Всі ці симпатичні волокна є дуже тонкими волокнами типу С і поширюються до всіх частин тіла по шляху скелетних нервів. Вони регулюють кровоносні судини, потові залози і м'язи, що піднімають волосся. Симпатичні волокна становлять в середньому приблизно 8% загального числа волокон скелетного нерва, що підтверджує їх величезне значення.
Симпатична нервова система. Симпатичний відділ автономної нервової системи включає в себепаравертебральні ганглії, що лежать у вигляді парних ланцюжків (стовбурів) вздовж всього хребта, а також непарні превертебральні ганглії і, крім цього,симпатичні нерви. У бокових рогах спинного мозку на рівні грудних і поперекових сегментів розміщений симпатичний центр Якобсона. Активність його нейронів регулюється через синаптичні контакти структурами стовбура головного мозку. Аксони нейронів симпатичного центра Якобсона виходять з спинного мозку в складі вентральних (передніх) корінців і утворюють білі з'єднувальні гілки з гангліями симпатичних стовбурів. Від цих стовбурів відходять постгангліонарні аксони і йдуть до органів голови, грудної, черевної та тазової порожнини в складі соматичних нервів, або в складі самостійних нервів. Це сірі з'єднувальні гілки. Аксони нейронів симпатичного центра Якобсона, що пройшли безпереривання через паравертебральні ганглії прямують до непарнихпревертебральних гангліїв, наприклад, серцевого, легеневого, черевного абосонячного сплетіння, верхнього брижевого, нижнього брижевого.Прегангліонарні аксони дають коллатералі також і до сусідніх гангліїв. Таким чином, прегангліонарний аксон, що виходить на рівні якого-небудь сегмента спинного мозку, іннервує декілька пара- і превертебральних гангліїв, чим забезпечується множинна (мультиплікантна) центральна регуляція різних вісцеральних функцій. Парасимпатична нервова система. Парасимпатичний відділ автономної нервової системи включає в себе ганглії, розміщені поблизу ефекторних органів або в них, а також парасимпатичні нерви. Тіла прегангліонарних парасимпатичних нейронів лежать в стовбурі мозку та в сакральному відділі спинного мозку. Аксони прегангліонарних нейронів йдуть допостгангліонарних нейронів, розміщених в гангліях. Прегангліонарні парасимпатичні волокна, що іннервуютьвнутрішньоочні м'язи, залози голови та ін. покидають стовбур мозку в складітрьох пар черепномозкових нервів - III (окоруховий), VII (лицевий), IX (язико-глотковий). До органів грудної і черевної порожнини прегангліонарні парасимпатичні волокна йдуть у складі X пари (блукаючий нерв). До органів порожнини тазу парасимпатичні волокна підходять у складі тазових нервів. На відміну від симпатичної нервової системи парасимпатична нервова система не інервує м'язи судин, за винятком статевих органів і можливо мозку.
Гормон[ред.] Матеріал з Вікіпедії — вільної енциклопедії. Зміни шаблонів/файлів цієї версії очікують на перевірку. Стабільна версія була перевірена 23 березня 2013.
Гормон (грец. Ορμόνη) — це біологічно-активна хімічна речовина, що виділяється ендокринними залозами безпосередньо у кров і впливає на певні органи і тканини-мішені або на організм в цілому[1]. Гормони є гуморальними (ті що переносяться з кров'ю) регуляторами певних процесів у певних органах і системах. Існують й інші визначення, згідно з якими трактування поняття гормон ширше: «сигнальні хімічні речовини, що виробляються клітинами тіла і впливають на клітини інших частин тіла» [ Джерело? ]. Це визначення є кращим, оскільки воно охоплює багато речовин, які традиційно зараховують до гормонів: гормони тварин, які позбавлені кровоносної системи наприклад, екдизони круглих червів, гормони хребетних, які виробляються не в ендокринних залозах (простагландини,еритропоетини тощо), а також гормони рослин.
Історія [ред.] Гормони були відкриті у 1902 році Старлінгом і Бейлісом. Призначення [ред.] Використовуються в організмі для підтримки його гомеостазу, а також для регуляції багатьох функцій (росту, розвитку, обміну речовин, реакції на зміни умов середовища). Рецептори [ред.] Всі гормони реалізують свою дію на організм або на окремі органи і системи за допомогою спеціальних рецепторів цих гормонів. Рецептори гормонів діляться на 3 основні класи: · рецептори, пов'язані з іонними каналами в клітині (іонотропні рецептори) · рецептори, що є ферментами, або пов'язані з білками-передавачами сигналу з ферментативною функцією (метаботропні рецептори, наприклад GPCR) · рецептори ретиноївої кислоти, стероїдних і тиреоїдних гормонів, які зв'язуються з ДНК і регулюють роботу генів. Для всіх рецепторів характерний феномен саморегуляції чутливості за допомогою механізмузворотному зв'язку — при низькому рівні певного гормону автоматично компенсаторно зростає кількість рецепторів у тканинах та їх чутливість до цього гормону — процес, що називають сенсибілізацією (а також ап-регуляцією (посилення, від англ. up-regulation), або сенситізацією (англ. sensitization)) рецепторів. І навпаки, при високому рівні певного гормону відбувається автоматичне компенсаторне зниження кількості рецепторів в тканинах та їх чутливості до цього гормону — процес, що називається десенсибілізацією (а також даун-регуляцією (від англ. down-regulation — зниження), або десенситизацією (англ. desensitization)) рецепторів. Збільшення або зменшення вироблення гормонів, а також зниження, або збільшення чутливості гормональних рецепторів і порушення гормонального транспорту призводить до ендокринних захворювань. Механізми дії [ред.] Коли гормон, що знаходиться в крові, досягає клітини-мішені, він вступає у взаємодію із специфічними рецепторами; рецептори «прочитують послання» організму, і в клітині починають відбуватися певні зміни. Кожному конкретному гормону відповідають виключно «свої» рецептори, що знаходяться в конкретних органах і тканинах, — тільки при взаємодії гормону з ними утворюється гормон-рецепторний комплекс. Механізми дії гормонів можуть бути різними. Одну з груп складають гормони, які з'єднуються з рецепторами, що знаходяться усередині клітин, — як правило, у цитоплазмі. До них належать гормони з ліпофільними властивостями — наприклад, стероїдні гормони (статеві гормони, глюко- і мінералокортикоїди), а також гормони щитовидної залози. Будучи жиророзчинними, ці гормони легко проникають через клітинну мембрану і починають взаємодіяти з рецепторами у цитоплазмі, або ядрі. Вони слабо розчинні у воді, при транспортуванні по крові зв'язуються з білками-носіями. Вважається, що в цій групі гормонів гормон-рецепторний комплекс виконує роль своєрідного внутрішньоклітинного реле — утворившись в клітині, він починає взаємодіяти з хроматином, який знаходиться в клітинних ядрах і складається з ДНК і білка, і тим самим прискорює або сповільнює роботу тих, або інших генів. Вибірково впливаючи на конкретний ген, гормон змінює концентрацію відповідною РНК і білка, і разом з тим коректує процеси метаболізму. Біологічний результат дії кожного гормону вельми специфічний. Хоча у клітині-мішені гормони змінюють зазвичай менше 1% білків і РНК, цього виявляється цілком достатньо для отримання відповідного фізіологічного ефекту. Більшість інших гормонів характеризуються трьома особливостями: · вони розчиняються у воді; · не зв'язуються з білками носіями; · починають гормональний процес, як тільки з'єднуються з рецептором, який може знаходитися в ядрі клітки, її цитоплазмі, або розташовуватися на поверхні плазматичної мембрани. У механізмі дії гормон-рецепторного комплексу таких гормонів обов'язково беруть участь посередники, які індукують відповідь клітини. Найважливіші з таких посередників — цАМФ(циклічний аденозинмонофосфат), інозитолтрифосфат, іони кальцію. Так, в середовищі, позбавленому іонів кальцію, або в клітках з недостатньою їх кількістю, дія багатьох гормонів послаблюється; при застосуванні речовин, що збільшують внутрішньоклітинну концентрацію кальцію, виникають ефекти, ідентичні до дії деяких гормонів. Участь іонів кальцію, як посередника забезпечує вплив на клітини таких гормонів, як вазопресині катехоламіни. Проте є гормони, внутрішньоклітинного посередника яких дотепер не виявлено. З найвідоміших таких гормонів можна назвати інсулін, у якого на роль посередника пропонували цАМФ і цГМФ, а також іони кальцію і навіть перекис водню, але переконливих доказів на користь якої-небудь однієї речовини немає. Багато дослідників вважають, що у такому разі посередниками можуть виступати хімічні з'єднання, структура яких повністю відрізняється від структури вже відомих науці посередників. Виконавши своє завдання, гормони або розщеплюються в клітинах-мішенях, або в крові, або транспортуються до печінки, де розщеплюються, або, нарешті, видаляються з організму в основному з сечею (наприклад, адреналін).
Дата добавления: 2016-03-26 | Просмотры: 879 | Нарушение авторских прав |


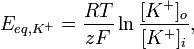 ,
,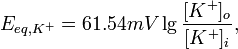
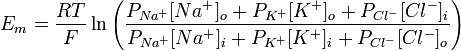
 Неперевірена версія
Неперевірена версія