|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
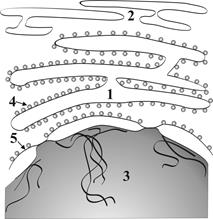
Г.2.1.2. Эндоплазматическая сеть - ЭПС● ЭПС представляет собой систему мембранных канальцев и цистерн, которые анастомозируя между собой образуют внутриклеточную сетеобразную структуру (рис. 5). ● По структурным особенностям различают два вида ЭПС: ● гранулярная (шероховатая) ЭПС с рибосомами на мембранах со стороны гиалоплазмы; ●агранулярная (гладкая) ЭПС без рибосом называется.
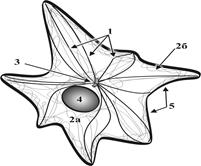
Рис. 5. Схема строения ЭПС: 1 – гранулярная ЭПС, 2 – агранулярная ЭПС, 3 – ядро клетки, 4 – фиксированные к мембране рибосомы, 5 – наружная ядерная мембрана. ►Основные функции гранулярной ЭПС связаны с синтезами белков «на экспорт», структурных белков клеточных мембран и ферментов лизосом. ►Основные функции гладкой ЭПС сопряжены с небелковыми синтезами (липиды, холестерин, гликоген и др.), накоплением и транспортом кальция, обезвреживанием ядовитых продуктов эндо- и экзогенного происхождения. ►По каналам ЭПС осуществляется поступление синтезированных веществ в комплекс Гольджи для их накопления. ►Усиление внутриклеточной синтетической активности клетки сопряжено с расширением цистерн и канальцев ЭПС и увеличением их количества Г.2.1.3. Комплекс Гольджи (рис. 6) представляет собой интеграцию полиморфных мембранных структур в околоядерной зоне клетки.
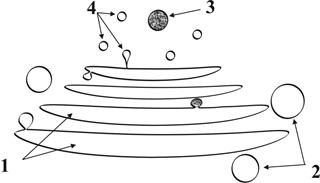
Рис. 6. Схема строения комплекса Гольджи: 1 – мембранные цистерны, 2 – мембранные вакуоли, 3 – секреторные гранулы, 4 – первичные лизосомы.
● В состав комплекса Гольджи входят следующие структуры: ● пакеты уплощенных мембранных цистерн ● большие и малые мембранные вакуоли ● секреторные гранулы (мембранные пузырьки с секретируемым содержимым) ● первичные лизосомы ► Комплекс Гольджи выполняет в клетке ряд важных функций: ► накопление и упаковка в гранулы (гранулообразование) синтезируемых на ЭПС веществ; ►выведение из клетки продуктов секреции; ►сборка новых биологических мембран для внутриклеточной регенерации (мембраногенез); ►образование лизосом. ● При функциональной активизации клетки в комплексе Гольджи происходит расширение цистерн, увеличение количества вакуолей и секреторных гранул. ● Комплекс Гольджи особенно хорошо развит в секреторных клетках. Г.2.1.4. Лизосомы (рис.7) ● Представляют собой мембранные пузырьки ● Ихдиаметр составляет 0,2 – 0,4 мкм ●Заполнены ферментами - катализаторами лизиса белков, жиров и углеводов. Эти ферменты синтезируются на шероховатой ЭПС и поступают в лизосомы через комплекс Гольджи ● Лизосомальная мембрана образуются в комплексе Гольджи.. Мембранная стенка лизосомы устойчива к действию собственных ферментов. ● Среди лизосом выделяют: первичные (мелкие, малоактивные), вторичные (крупные активные), аутолизосомы (обеспечивают процессы аутолиза – растворения и уничтожения собственных структур клетки), гетеролизосомы (обеспечивают процессы расщепления и растворения продуктов эндоцитоза – см.ниже) ● Количество лизосом в клетке крайне изменчиво. Число аутолизосом возрастает при усилении процессов, сопряженных с разрушениями клеточных структур (усиление процессов функционирования и внутриклеточной регенерации, повреждения клетки и др.). ● При старении клетки имеет место увеличение количества аутолизосом с пониженной ферментативной активностью. Это приводит к накоплению в клетке «недопереваренных» продуктов эндоцитоза и аутофагии, которые называются остаточными тельцами, т.е. происходит «замусоривание» клетки.
► Функции лизосом связаны с процессами внутриклеточного и внеклеточного пищеварения: ►активизированные (вторичные) лизосомы участвуют в расщеплении и лизисе продуктов эндоцитоза; ►отдельная популяция лизосом, аутолизосомы, выделяя свои ферменты в гиалоплазму или сливаясь с измененными органеллами, инициируют процессы аутолиза (ферментативное растворение собственных структур клетки) и аутофагии. ► некоторые клетки (например, макрофаги) выделяют лизосомальные ферменты в межклеточное пространство для разрушения остатков погибших клеток и тканей собственного организма, а также внедрившихся микроорганизмов.
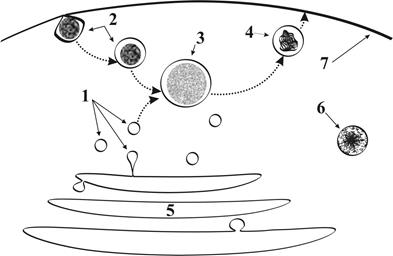
Рис. 7. Лизосомы и пероксисомы: 1 –лизосома, 2 – эндосома, 3 – пищеварительная вакуоль, 4 – остаточное тельце, 5 – комплекс Гольджи, 6 – пероксисома, 7 – цитолемма.
Г.2.1.5. Пероксисомы (рис.7) ● Представляют собоймембранные пузырьки. ● Ихдиаметр составляет 0,2 – 0,4 мкм. ● Заполнены ферментами метаболизма перекиси водорода. ● Отшнуровываются от расширенных участков канальцев гладкой ЭПС. ● Имеются во всех клетках, ноособенно многочисленны в клетках печени и почек, где активно протекают процессы дезинтоксикации (обезвреживание ядовитых продуктов метаболизма).
► Функции пероксисом связаны с процессами внутриклеточной дезинтоксикации: ► образование перекиси водорода – сильнейшего окислителя, который используется в целях дезинтоксикации (обезвреживания) конечных продуктов клеточного метаболизма. ►разрушение «избытков» перекиси водорода, которая обладает токсическим действием на клетку.
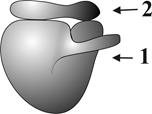
Г.2.1.6. Рибосомы (рис. 8) – немембранные органеллы. ● Функционирующие рибосомы состоят из двух связанных субъединиц (большой и малой), образованных рибонуклеопротеидами. ● Размер рибосом не превышает 25 нм.
Рис. 8. Схема рибосомы: 1 – большая субъединица, 2 – малая субъединица. ● Субъединицы рибосом образуются в ядрышке, а их сборка происходит в цитоплазме. ● Часть рибосом располагается в гиалоплазме - свободные рибосомы, другие рибосомы связываются с мембранами шероховатой ЭПС - связанные рибосомы. ● Некоторые рибосомы объединяются в комплексы – полисомы. ● Кроме цитоплазматических рибосом имеются митохондриальные рибосомы, которые кодируются митохондриальной ДНК. Часть рибосом находится на наружной мембране кариолеммы (ядерная оболочка см. ниже).
► Функции рибосом связаны с генетически запрограммированным внутриклеточным синтезом белка.
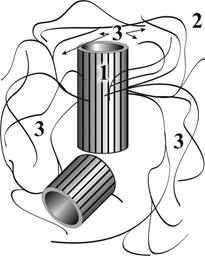
Г.2.1.7. Центросома - клеточный центр (рис. 9)
● Центросома - комплексный немембранный органоид, ● Центросомаявляется частью цитоскелета (см. ниже). ● Центросома состоит из двух центриолей (материнской и дочерней)и центросферы. ● Центросомаобладает структурной динамичностью, зависящей от состояния клетки. ●Характеристики центросомы неделящейся клетки: ▬расположена около ядра вблизи комплекса Гольджи; ▬ центриоли (материнская и дочерняя) составляют диплосому и представляют собой цилиндры (длина 0,3 мкм и диаметр 0,1 мкм), расположенные перпендикулярно друг к другу (рис.10); ▬ стенку каждого цилиндра составляют девять триплетов микротрубочек, построенных из тубулиновых белков (рис.10); ▬ к каждому триплету с наружной стороны присоединено сферическое белковое тельце – сателлит; ▬ от сателлитов материнской центриоли в гиалоплазму отходят микротрубочки, которые формируют центросферу.
Рис. 10. Схема строения материнской центриоли: 1 – триплеты тубулиновых белков, 2 - сателлит, 3 – микротрубочки.
●Характеристики центросомы делящейся митозом клетки: ▬ при подготовке клетки к митотическому делению происходит матричное удвоение (дубликация) и расхождение центриолей по полюсам клетки; ▬ на каждом полюсе клетки формируется своя диплосомная центриоль, которая участвуют в образовании центросферы и микротрубочек веретена деления; ▬ микротрубочки прикрепляются к хромосомам и обеспечивают перемещение хромосом по полюсам, а также их распределение между дочерними клетками; ▬ после завершения митоза центриоль каждой дочерней клетки приобретает характеристики интерфазной (см. выше).
► Функции центросом: ► индуцирование полимеризации тубулиновых белков и сборку микротрубочек; ► комплексирование (создание) компонентов цитоскелета ► внутриклеточное перемещением хромосом при митозе.
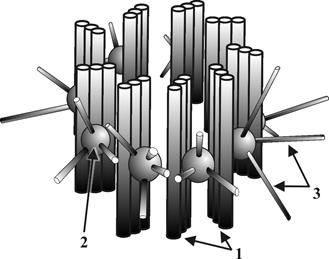
Г.2.1.8. Цитоскелет – внутриклеточный трехмерный немембранный структурный комплекс (рис. 11).
Рис. 11. Схема строения цитоскелета: 1 – микротубулы цитоскелета, 2 – микрофиламенты цитоскелета, 2б – микрофиламенты кортекса цитоскелета, 3 клеточный центр, 4 – ядро, 5 – цитолемма.
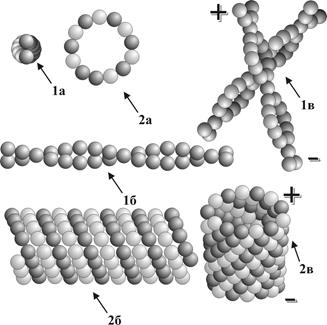
● Цитоскелет включает в себя собственно цитоскелет, а также тубулярно – фибриллярные элементы кортекса, центросомы, микроресничек и микроворсинок. ● Основными структурными элементами цитоскелета являются микротрубочки (микротубулы), микрофиламенты (рис. 12 ) и промежуточные филаменты. ●Характеристики микротрубочек: ▬ представляют собой полые неветвящиеся цилиндры диаметром около 20 нм; ▬ стенки цилиндров построены из молекул тубулиновых белков; ▬ первичный синтез тубулиновых белков осуществляется на свободных рибосомах, а пространственная сборка на центросомах и базальных тельцах; ▬ структура микротрубочек обладает выраженной пространственной динамичностью за счет постоянно текущих на их противоположных полюсах процессов полимеризации и деполимеризации тубулинов. ●Характеристики микрофиламентов: ▬ представляют собой нитчатые двухцепочечные структуры диаметром 5 нм, которые собираются в микропучки и образуют в цитоплазме сетеобразные структуры различной степени сложности; ▬ построены из молекул сократительных белков (преимущественной из актина), первичный синтез которых осуществляется на свободных рибосомах; ▬ способны к активному АТФ-обеспечиваемому и кальций-зависимому сокращению; ▬ на полюсах микрофиламентов протекают процессы деполимеризации белков под действием лизосомальных ферментов и полимеризации при участии цитоплазматических актин-связывающих белков.
●Характеристики промежуточных филаментов: ▬ Являются дополнительным структурным элементом цитоскелета. ▬ Это относительно короткие ветвящиеся нитчатые образования диаметром 10 нм. ▬ Они построены из опорно-каркасных белков (кератина, виментина, десмина). ▬ Преимущественно развиты в клетках тканей, испытывающих механические нагрузки.
►Цитоскелет осуществляет в клетке локомоторную функцию. Она заключается: ►в создании опорного внутриклеточного каркаса; ►в организации межклеточных контактов; ► в поддержании и изменении формы клетки; ► в обеспечении внутриклеточных транспортов и структурных перемещений; ► в обеспечении передвижений свободно существующих клеток в пространстве; ► в участии в делении клетки.
Рис. 12 Схема строения элементов микротрубочек и микрофиламентов: 1а – микрофиламенты, поперечный срез; 1б – микрофиламенты, вид сбоку; 1в – объемный вид микрофиламентов; 2а – микротрубочки, поперечный срез; 2б – микротрубочки, вид сбоку; 2в – объемный вид микротрубочек; (+) – полюс полимеризации; (-) – полюс деполимеризации. Г.2.2. Включения – непостоянные структурные компоненты цитоплазмы, образующиеся в процессе клеточного метаболизма. Их количество зависит от функционального состояния клетки. Среди включений различают несколько структурно-функциональных типов: ●трофические (капли липидов, белковые гранулы, глыбки гликогена); ● пигментные (гемоглобин, билирубин, меланин, липофусцин); ● секреторные (гранулы с синтезированными клеткой биологически активными веществами, подлежащими экзоцитозу с целью регуляции жизнедеятельности других клеток и тканей); ● экскреторные (продукты клеточного метаболизма, подлежащие выведению с целью нейтрализации или уничтожения).
Г.2.3. Гиалоплазма – коллоидный аморфный матрикс цитоплазмы, который создает специфическое микроокружение для клеточных структур, обеспечивает их жизнедеятельность и взаимодействие. ● Гиалоплазма имеет консистенцию жидкого геля ● В состав гиалоплазмы входит связанная и свободная вода, растворы минеральных солей, биополимеры белковой, липидной и углеводной природы. ● Она способна менять своё агрегатное состояние (становиться более жидкой или более вязкой) в зависимости от состояния жизнедеятельности клетки, а также проникновения в клетку чужеродных агентов. .
● В гиалоплазме обнаружена мелкопетлистая микротрабекулярная сеть, которая может распадаться и собираться вновь в зависимости от функционального состояния клетки, фазы митотического цикла, в ходе внутриклеточных восстановительных процессов или при дедифференцировке.
► Функции гиалоплазмы: ►создание постоянства внутриклеточной среды; ►обеспечение условий для внутриклеточных транспортов и перемещений; ►интеграция органелл в функциональные комплексы; ►отложение запасных продуктов в виде включений; ►обменные процессы с внутриядерным и межклеточным веществом, поддержание объемного постоянства клетки. Г.3. Ядро – является одной из основных структурных частей эукариотической клетки (рис. 13). ● Ядро содержит основной объем ДНК, которая является ключевым субстратом генетического аппарата. ● Как целостная структура ядро существует в клетке в период интерфазы митотического цикла. ● В клетке может быть одно или несколько ядер
►Основные функции ядра связаны с процессами хранения, воспроизведения, передачи и реализации наследственной информации.
Ядро состоит из структурных (кариолемма, кариоскелет, хроматин, ядрышко,) и неструктурного (кариоплазма ) компонентов.
Г.3.1. Кариолемма – ядерная оболочка (рис.13), отделяющая кариоплазму от цитоплазмы и обеспечивающая обмен между ними.
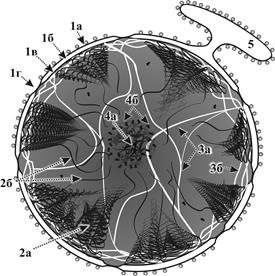
Рис. 13. Схема строения ядра: 1а – наружная мембрана кариолеммы, 1б – перинуклеарное пространство, 1в – внутренняя мембрана кариолеммы, 1г – ядерная пора, 2а – гетерохроматин, 2б – эухроматин, 3а – кариоскелет, 3б – ламина, 4а – фибриллярный компонент ядрышка, 4б – гранулярный компонент ядрышка, 5 – гранулярная ЭПС.
● Кариолеммаобразована двумя биомембранами (наружной и внутренней), разделенными перинуклеарным пространством. ● В областях локальных слияний этих мембран имеются ядерные поры, соединяющие цитоплазму с содержимым ядра. Ядерные поры обеспечивают поступление молекул РНК и субъединиц рибосом из ядра в цитоплазму. В обратном направлении через них происходит активный транспорт синтезированных белков. ● На наружной мембране кариолеммы имеются рибосомы. ● К внутренней мембране со стороны кариоплазмы плотно прикрепляется ядерная пластинка (ламина). Она построенная из опорных белковых филаментов, соединенных с кариоскелетом (см. ниже). Имеет важное значение в поддержании формы ядра, в создании пространственной организации ядерных пор и хроматина
Г.3.2. Хроматин (рис.13)– это структурный предшественник хромосом в интерфазном ядре. ● Хроматин состоит из комплекса ДНК и хромосомных белков, которые регулируют степень спирализации хроматина. ● Хроматин может присутствовать в двух структурных формах: ▬ гетерохроматин (спирализованный или конденсированный) ▬ эухроматин (деспирализованный или деконденсированный) ● Эти формы способны переходить одна в другую. Их объемное соотношение в интерфазном ядре постоянно изменяется.
● Гетерохроматин является плотно упакованным транскрипционно неактивным хроматином. Он выявляется в световом микроскопе в виде базофильных глыбок преимущественно на периферии ядра или вокруг ядрышек. Этот хроматин специализирован на хранении генетической информации. Его количество максимально увеличивается к началу митоза. ● Эухроматин практически невидим в световом микроскопе. С него происходит считывание (транскрипция) генетической информации для последующей реализации в цитоплазме в виде активизации синтетических процессов. Поэтому эухроматин называется «функциональным». Его максимальное количество приходится на пресинтетический период интерфазы.
Во время клеточного деления (митоз или мейоз) хроматин полностью спирализуется и образует палочковидные, хорошо окрашивающиеся структуры – хромосомы.
● Во всех соматических клетках генетически женского организма одна из половых Х-хромосом характеризуется стойкой конденсацией (спирализацией) в интерфазе - это Х-половой хроматин. Он обнаруживается в ядре с помощью светового микроскопа при окрашивании клеток щелочными красителями и называется тельцем Бара. Микроскопическое выявление телец Барра имеет значение в судебно-медицинской практике для определения генетического пола.
Г.3.3. Ядрышко (рис.13) – базофильная непостоянная структура интерфазного ядра. ● Располагается в центре ядра или несколько эксцентрично. ● Количество и размеры ядрышек зависят от уровня метаболизма и функциональной активности клетки. ● Ядрышко не имеет собственной оболочки. ● Оно образовано специализированными участками некоторых хромосом, которые называются ядрышковыми организаторами.
► Функции ядрышка: ► Синтез рибосомальной РНК ► Формирование субъединиц рибосом. Последние выходят через ядерные поры в цитоплазму, попарно соединяются и образуют рибосомы. Г.3.4. Кариоскелет (рис.13 ) – трехмерная сетевидная структура, заполняющая весь внутренний объем ядра. ● Состоит из опорных фибриллярных белков, которые образуют тонкопетлистую сеть. ● Крепится к ядерной пластинке (ламине)
►Функции кариоскелета: ► поддержание и изменение формы ядра; ► пространственное распределение хроматина и его спирализация; ► передвижение субъединиц рибосом; ► регуляция ширины перинуклеарного пространства, ► регуляция величины и количества ядерных пор. Г.3.5. Кариоплазма (ядерный сок) – внутриядерная коллоидная аморфная субстанция. ► Функции кариоплазмы: ►поддержание постоянства внутриядерной среды; ►обеспечение условий для внутриядерных транспортов и перемещений,обменные процессы с цитоплазмой ► создание микроокружения для структурных компонентов ядра.
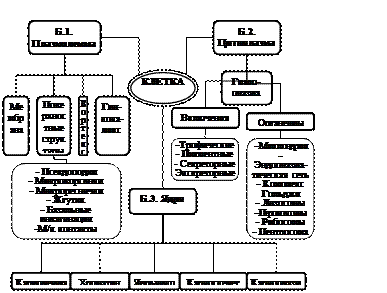
Г.4. Общий план строения клетки
Д. ЦИТОФИЗИОЛОГИЯ ● Это раздел цитологии, изучающий нормальную жизнедеятельность клетки ● Клетка является комплексной жизнеспособной биосистемой, которая обладает запрограммированной надежностью и резервностью. ● Все структуры живой клетки находятся в состоянии постоянной пространственной мобильности и морфофункциональных взаимосвязей в рамках генетически обусловленного гомеостаза. ● Жизнедеятельность клетки – это беспрерывная череда адаптационно-компенсаторных реакций, смен режимов рабочей активности и относительного покоя, процессов восстановления, самообновления, воспроизведения и старения, которые обеспечиваются интеграцией всех клеточных компонентов в единое морфофункциональное целое. Дата добавления: 2016-06-05 | Просмотры: 806 | Нарушение авторских прав |