|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Вышеуказанные показатели характеризуют не столько саму функцию дыхания, сколько потенциальную способность к выполнению этой функции.Измерение легочных объемов и емкостей обычно производят методами спирометрии, пневмотахометрии с интеграцией показателей и бодиплетизмографии Cпироме́трия — метод исследования функции внешнего дыхания, включающий в себя измерение объёмных и скоростных показателей дыхания. Прибор, с помощью которого осуществляется спирометрическое исследование, называется спирометром. Спирографи́я (лат. spiro дышать + греч. graphō писать, изображать)метод исследования функции легких путем графической регистрации во времени изменений их объема при дыхании. Спирография предназначена для оценки состояния легких путем измерения объема и скорости выдыхаемого воздуха. Данная процедура является безболезненной и не связана с введением в организм пациента каких-либо медицинских инструментов, занимает всего несколько минут и может проводиться в амбулаторных условиях. Пневмотахометрия — определение пиковых скоростей воздушного потока, достигаемых в режиме «дыхательного толчка». Получаемые показатели неточно называют мощностью выдоха и вдоха (Мвыд и Мвд). По своей сущности метод близок к спирограмме форсированного выдоха и в большей мере, чем ОФВ1 отражает сопротивление воздушному потоку, возникающее на уровне крупных бронхов. 12. Давление в плевральной полости. Изменения при спокойном и глубоком дыхании. При вдохе объем грудной полости возрастает. А так как плевральная изолирована от атмосферы, то давление в ней понижается. Легкие расширяются, давление в альвеолах становится ниже атмосферного. Воздух через трахею и бронхи поступает в альвеолы. Во время выдоха объем грудной клетки уменьшается. Давление в плевральной щели возрастает, воздух выходит из альвеол. Движения или экскурсии легких объясняются колебаниями отрицательного межплеврального давления. После спокойного выдоха оно ниже атмосферного на 4-6 мм.рт.ст. На высоте спокойного вдоха на 8-9 мм.рт.ст. После форсированного выдоха оно ниже на 1-3 мм.рт.ст., а форсированного вдоха на 10-15 мм. рт. ст. Наличие отрицательного межплеврального давления объясняется эластической тягой легких. Это сила, с которой легкие стремятся сжаться к корням, противодействуя атмосферному давлению. Она обусловлена упругостью легочной ткани, которая содержит много эластических волокон. Кроме того, эластическую тягу увеличивает поверхностное натяжение альвеол. 13. Транспульмональное давление. Пневмоторакс. Разница между альвеолярным и внутриплевральным давлением называют транспульмоналъным давлением. Она представляет собой разницу между давлением внутри альвеол и давлением на внешней поверхности легких. Транспульмональное давление является мерой эластических сил в легких, стремящихся к уменьшению объема легких в любой фазе дыхания. Это давление называют давлением коллапса. Пневмото́ракс — скопление воздуха или газов в плевральной полости. Он может возникнуть спонтанно у людей без хронических заболеваний лёгких, а также у лиц с заболеваниями лёгких и искусственный пневмоторакс. 14. Эластическая тяга лёгких, формула вычисления. Изменение в онтогенезе. Эластическая тяга легких – сила, которая стремится уменьшить объём лёгких. Эластическое сопротивление лёгких: En=изменение Ртранспульм./изменение Vлёгких 15. Растяжимость лёгких, формула вычисления. Изменения в онтогенезе. С = V/Δ P где С — растяжимость (англ. — compliance), dV — изменение легочного объема (мл), а dР — изменение внутриплеврального давления (см водн. ст.). 16. Рефлекторное изменения дыхания при раздувании лёгких. Дыхательный центр получает афферентные импульсы не только от хеморецепторов, но и от прессорецепторов сосудистых рефлексогенных зон, а также от механорецепторов легких, дыхательных путей и дыхательных мышц. Все эти импульсы вызывают рефлекторные изменения дыхания. Особенно важное значение имеют импульсы, поступающие к дыхательному центру по блуждающим нервам от рецепторов легких. От них в большой степени зависит глубина вдоха и выдоха. Рефлекс Геринга — Брейера. Раздувание легких рефлекторно тормозит вдох и вызывает выдох. Перерезка блуждающих нервов устраняет рефлекс. Нервные окончания, расположенные в бронхиальных мышцах, играют роль рецепторов растяжения легких. Их относят к медленно адаптирующимся рецепторам растяжения легких, которые иннервируются миелинизированными волокнами блуждающего нерва. Рефлекс Геринга — Брейера контролирует глубину и частоту дыхания. У человека он имеет физиологическое значение при дыхательных объемах свыше 1 л (например, при физической нагрузке). У бодрствующего взрослого человека кратковременная двусторонняя блокада блуждающих нервов с помощью местной анестезии не влияет ни на глубину, ни на частоту дыхания. У новорожденных рефлекс Геринга — Брейера четко проявляется только в первые 3—4 дня после рождения.
17. Рефлекторное изменения дыхания при спадении лёгких ниже ФОЭ.
18.Альвеолярный воздух, его состав, методика определения. Причины и значение относительного постоянства состава. кислород в альвеолярном воздухе - 14,2-14,6%, углекислота 5,5-5,7%, азот около 80%. Парциальное давление кислорода и углекислого газа в А. в. имеет большое значение, т. к. определяет диффузионный обмен газов. Из альвеол кислород диффундирует в кровь, а из крови углекислый газ — в альвеолы. Снижение содержания кислорода в А. в. рефлекторно вызывает спазм легочных артериол и гипертензию малого круга кровообращения. Состав А. в., и прежде всего содержание кислорода, в различных отделах легких несколько отличается, особенно при легочной патологии. Объем А. в, увеличивается при эмфиземе, снижается при ателектазах и отеке легкого. Объем всего воздуха, содержащегося в альвеолах и дыхательных путях, можно измерить методом разведения индикаторного газа (гелий, азот, радиоактивный ксенон и др.). Среднее парциальное напряжение углекислого газа в А. в. (РАСО2) всегда близко к напряжению углекислого газа артериальной крови (РАСО2) за исключением случаев тяжелой легочной патологии. Напряжение кислорода в А. в. (РАО2) можно подсчитать по уравнению альвеолярного воздуха: РАО2 = РIО2 - РАСО2 ⋅ 1,2 где РIО2 — напряжение кислорода во вдыхаемом воздухе (обычно 150 мм рт. ст.); РАСО2 — напряжение СО2 в А. в. (его измеряют в конечных порциях выдыхаемого воздуха капнографом или приравнивают к РАСО2, обычно измеряемому с помощью прибора Аструпа в порциях крови, набранной из артерии или из пальца); 1,2 — фактор поправки при обычной величине дыхательного коэффициента, равной 0,8. Определение напряжения газов А. в. важно для оценки газообмена в легких.
19. Вентиляционно-перфузионное отношение. Особенности вентиляции и перфузии в различных отделах лёгкого. В норме величина вентиляционно-перфузионного отношения (ВПО) равна 0,8-1,0. У здоровых людей этот показатель отражает адекватность минутного объёма альвеолярной вентиляции (V) минутному объёму кровотока (Q) в лёгких, то есть V/Q = 0,8-1,0. У человека в вертикальном положении тела величина легочного кровотока на единицу объема ткани легкого линейно убывает в направлении снизу вверх, и меньше всего снабжаются кровью верхушки легких. Соответственно в положении тела человека на спине кровоток в нижних (дорсальных) отделах легких становится выше, чем в верхних (вентральных). Это обусловлено тем, что артериальная кровь, поступающая в легкие из правого желудочка, проходит по сосудам легких из областей низкого внутриплеврального давления в области тонкостенных капилляров, которые окружены альвеолами, содержащими воздух под давлением, близким к атмосферному. Поэтому в зависимости от соотношения давления в альвеолах (РА), мелких артериях (Ра) и мелких легочных венах (Pv) легкие разделены на функциональные зоны Веста В верхушках легких (зона 1) могут возникнуть области с давлением в легочных капиллярах (особенно в фазу диастолы) ниже альвеолярного (Ра > Ра > Pv)- Капилляры в таких зонах могут спадаться, и кровоток через них становится невозможным. Такие участки легких вентилируются, но не участвуют в газообмене и формируют альвеолярное мертвое пространство. В средних отделах легких (зона 2) под действием гравитации давление в альвеолах, как правило, превышает венозное (Ра > РА > Pv). Поэтому величину кровотока в зоне 2 по Весту определяет разность между артериальным и альвеолярным давлениями. В зоне 2 практически не возникает альвеолярное мертвое пространство. В нижних отделах легких (зона 3) давление в легочных венах выше альвеолярного (Ра > Pv > Ра) и величина кровотока, как и в обычных сосудах, определяется разницей между артериальным и венозным давлениями.
20. Газообмен в лёгких. Диффузионная способность лёгких. Транспорт О2 осуществляется в физически растворенном и химически связанном виде.. Согласно закону Фика, газообмен О2 между альвеолярным воздухом и кровью происходит благодаря наличию концентрационного градиента О2 между этими средами. В альвеолах легких парциальное давление О2 составляет 13,3 кПа, или 100 мм рт.ст., а в притекающей к легким венозной крови парциальное напряжение О2составляет примерно 5,3 кПа, или 40 мм рт.ст. Давление газов в воде или в тканях организма обозначают термином «напряжение газов» и обозначают символами Ро2, Рсo2. Градиент О2 на альвеолярно-капиллярной мембране, равный в среднем 60 мм рт.ст., является одним из важнейших, но не единственным, согласно закону Фика, факторов начальной стадии диффузии этого газа из альвеол в кровь. Транспорт О2 начинается в капиллярах легких после его химического связывания с гемоглобином. Гемоглобин (Нb) способен избирательно связывать О2 и образовывать оксигемоглобин (НbО2) в зоне высокой концентрации О2 в легких и освобождать молекулярный О2 в области пониженного содержания О2 в тканях. При этом свойства гемоглобина не изменяются и он может выполнять свою функцию на протяжении длительного времени. Метаболические факторы являются основными регуляторами связывания О2 с гемоглобином в капиллярах легких, когда уровень O2, рН и СО2 в крови повышает сродство гемоглобина к О2 по ходу легочных капилляров. В условиях тканей организма эти же факторы метаболизма понижают сродство гемоглобина к О2 и способствуют переходу оксигемоглобина в его восстановленную форму — дезоксигемоглобин. В результате О2 по концентрационному градиенту поступает из крови тканевых капилляров в ткани организма. Оксид углерода (II) — СО, способен соединяться с атомом железа гемоглобина, изменяя его свойства и реакцию с О2. Очень высокое сродство СО к Нb (в 200 раз выше, чем у О2) блокируют один или более атомов железа в молекуле гема, изменяя сродство Нb к О2. Переход О2 из альвеолярного воздуха в кровь легочных микрососудов, а СО2 в обратном направлении осуществляется путем диффузии по градиенту концентрации газов в указанных средах. Показателей, характеризующим диффузию газов, является величина, обратная сопротивлению диффузии, называемая диффузионной способностью легких (ДL). Этот параметр показывает количество мл. газа, проходящее через легочную мембрану за 1 мин. при трансмембранной разности парциальных давлений газа в 1 мм.рт. ст. У здорового человека в покое ДL О2 легких составляет 15-20 мл О2/мин./мм.рт.ст. ДL CО2 в 20 раз выше, поэтому ограничения диффузии СО2 в легких практически не существует. 21. Содержание О2 и СО2 в венозной и артериальной крови. В состоянии растворения в артериальной крови содержится 0,25 мл О2, 2,69 мл СО2 и 1,04 мл N. В венозной крови содержится 12% кислорода, 55—57% углекислого газа и около 1% азота. артериальная кровь здорового человека содержит 18-20% кислорода, 50-52% углекислоты и около 1% азота. 22.Кислородная ёмкость крови. Кислородная ёмкость крови — количество кислорода, которое может быть связано кровью при её полном насыщении в 100мл; выражается в объёмных процентах (% об.); зависит от концентрации в крови гемоглобина. Определение Кислородной ёмкости крови важно для характеристики дыхательной функции крови. Кислородная ёмкость крови человека — около 18—20 % об. 23. Значение физически растворённых О2 и СО2 в крови. В артериальной крови содержание физически растворенного кислорода составляет 0,003 мл кислорода на 1 мл крови, а содержание углекислого газа в артериальной крови - 0,026 мл на 1 мл крови. Хотя содержание в крови кислорода и углекислого газа в физически растворенном состоянии относительно невелико, это состояние играет огромную роль в жизнедеятельности организма. Для того, чтобы связаться с теми или иными веществами, дыхательные газы сначала должны быть доставлены к ним в физически растворенном виде. Таким образом, при диффузии в ткани или кровь каждая молекула кислорода или углекислого газа определенное время пребывает в состоянии физического растворения.
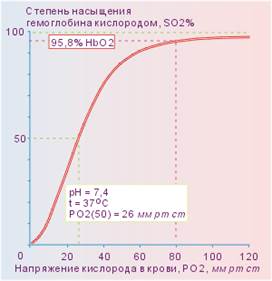
24. Кривая диссоциации оксигемоглобина. Значение крутой и отлогой её частей.
Из графика видно, что при высоких значениях напряжения кислорода в крови (правая треть графика) угол наклона минимален. На этом участке графика, соответствующем напряжению кислорода в артериальной крови, при значительных изменениях напряжения кислорода (~60 ÷ 100 мм рт ст) степень насыщения гемоглобина кислородом велика (>90%) и мало изменяется (~90 ÷ 97%). кислородом значительно уменьшается. При этом кислород немедленно используется в метаболизме тканей.
25. Факторы, определяющие сродство Hb к О2. 1. увеличение температуры = снижение сродства 2. Влияние рН (эффект Бора): снижение рН снижение сродства. увеличивается РСО2 = снижается рН +собств.эффект на гемоглобин. 3. Накопление углекислоты (взаимодействует с глобиновой частью Нb - снижение его сродства к кислороду) 26. Факторы, влияющие на ход кривой диссоциации оксигемоглобина: t, pH, PСО2 1. Температура. увеличение температуры =>кривая – вправо, снижеается угол наклона (снижение сродства); 2. Влияние рН (эффект Бора): снижение рН =>уменьшение угла наклона (снижение сродства). увеличивается РСО2 = снижается рН +собств.эффект на гемоглобин. 3. чем выше РСО2, тем ниже рН. Увеличение напряжения СО2 в крови сопровождается снижением сродства гемоглобина к кислороду и уменьшением угла наклона кривой диссоциации НbО2. 27. Расчёт коэффициента утилизации кислорода. -процентное отношение доли кислорода, используемой тканями (разности концентраций кислорода вартериальной и венозной крови), к общему содержанию его в артериальной крови. 28. Перенос СО2 кровью. Образование бикарбонатов и карбогемоглобина. В венозной крови содержится около 580 мл / л С02. С02 образуется в тканях при окислительных процессах. В большинстве тканей Рсо2 составляет 50-60 мм рт. ст. (6,7-8 кПа). В крови, поступающей в артериальное конец капилляров, РаCO2 составляет около 40 мм рт. ст. (5,3 кПа). Наличие градиента заставляет С02 диффундировать из тканевой жидкости до капилляров. Чем активнее в тканях осуществляются процессы окисления, тем больше создается СОТ и тем больше Ртк.со2. Кровь, проходящая через легкие, отдает далеко не весь С02. Большая часть его остается в артериальной крови, поскольку соединения, которые образуются на основе С02, участвуют в поддержании кислотно-основного равновесия крови - одного из параметров гомеостаза. Направление реакции определяется непрерывным выходом С02 с эритроцита в плазму, а из плазмы в альвеолы. В легких в связи с постоянным его выделением происходит реакция диссоциации карбогемоглобин: 29. Значение карбоангидразы. С02, который проникает в кровь, сначала подвергается гидратации с образованием угольной кислоты: С02 + Н20 Н2СОз. 30. Газообмен между кровью и тканями Перенос O2 и СO2 между кровью системных капилляров и клетками тканей осуществляется путем простой диффузии, т. е. так же, как между кровью легочных капилляров и альвеолярным воздухом. Скорость переноса газа через слой ткани прямо пропорциональна площади слоя и разнице парциального давления газа по обе его стороны и обратно пропорциональна толщине слоя. При газообмене между тканями и кровью толщина диффузионного барьера менее 0,5 мкм, однако в мышцах в состоянии покоя расстояние между открытыми капиллярами составляет около 50 мкм. При работе, когда потребление кислорода мышцами увеличивается, открываются добавочные капилляры, что уменьшает диффузионное расстояние и увеличивает диффузионную поверхность. Поскольку СO2 диффундирует в тканях примерно в 20 раз быстрее, чем O2, удаление углекислого газа происходит гораздо легче, чем снабжение кислородом. 31. Гипоксия и гиперкапния. Отличие гипоксии от асфиксии. гиперкапния – увеличение содержания углекислоты. Гипоксия - это недостаток кислорода в тканях или кислородное голодание тканей. Асфиксия (беспульсие) - остропротекающее удушье вследствие альвеолярной гиповентиляции с гипоксемией и гиперкапнией (утопление, удушение, отек легких). 32. Дыхательный центр продолговатого мозга Дыхательный центр представляет собой парное образование, состоящее из центра вдоха (инспираторного) и центра выдоха (экспираторного). Каждый центр регулирует дыхание одноименной стороны: при разрушении дыхательного центра с одной стороны наступает прекращение дыхательных движений с этой стороны. Нейроны верхнего отдела моста, регулирующие акт дыхания, были названы пневмотаксическим центром. Дыхательный центр, расположенный в продолговатом мозге, посылает импульсы к мотонейронам спинного мозга, иннервирующим дыхательные мышцы. Диафрагма иннервируется аксонами мотонейронов, расположенных на уровне III—IV шейных сегментов спинного мозга. Мотонейроны, отростки которых образуют межреберные нервы, иннервирующие межреберные мышцы, расположены в передних рогах (III—XII) грудных сегментов спинного мозга. 33. Автоматизм дыхательного центра, отличия от автоматизма проводящей системы сердца. Автоматия дыхательного центра поддерживается и изменяется под влиянием импульсов от дыхательных мышц, сосудистых рефлексогенных зон, различных интеро- и экстерорецепторов, а также под влиянием многих гуморальных факторов (рН крови, содержание углекислого газа и кислорода в крови и др). 34. Пневмотаксический и апнеистический центр. В структурах моста тоже различают два дыхательных центра. Один из них — пневмотаксический — способствует смене вдоха на выдох (за счет переключения возбуждения из центра вдоха на центр выдоха); второй центр осуществляет тоническое влияние на дыхательный центр продолговатого мозга. 35. Основные скопления дыхательных нейронов продолговатого мозга. Их связи и значение. Выделяют дорзальную группу нейронов, активирующих мышцы вдоха, — инспираторный отдел и вентральную группу нейронов, контролирующих преимущественно выдох, — экспираторный отдел. В каждом из этих отделов имеются различные по свойствам нейроны. Среди нейронов инспираторного отдела выделяют: 1) ранние инспираторные — их активность повышается за 0,1-0,2 с до начала сокращения инспираторных мышц и длится в течение вдоха; 2) полные инспираторные — активны во время вдоха; 3) поздние инспираторные — активность повышается в середине вдоха и заканчивается в начале выдоха; 4) нейроны промежуточного типа. Часть нейронов инспираторного отдела обладает способностью самопроизвольно ритмически возбуждаться. Описаны аналогичные по свойствам нейроны в экспираторном отделе дыхательного центра. Взаимодействие между этими нейронными пулами обеспечивает формирование частоты и глубины дыхания. Нейроны инспираторного отдела получают информацию о напряжении газов в артериальной крови, рН крови от хеморецепторов сосудов и о рН ликвора от центральных хеморецепторов, расположенных на вентральной поверхности продолговатого мозга. К дыхательному центру поступают также нервные импульсы от рецепторов, контролирующих растяжение легких и состояние дыхательных и других мышц, от терморецепторов, болевых и сенсорных рецепторов. Сигналы, поступающие к нейронам дорзальной части дыхательного центра, модулируют их собственную ритмическуюактивность и оказывают влияние на формирование ими потоков эфферентных нервных импульсов, передающихся в спинной мозг и далее к диафрагме и наружным межреберным мышцам. Эфферентные нервные импульсы от инспираторных нейронов передаются по нисходящим волокнам в составе вентрального и передней части бокового канатика белого вещества спинного мозга к а-мотонейронам, формирующим диафрагмальные и межреберные нервы. Все волокна, следующие к мотонейронам, иннервирующим мышцы выдоха, являются перекрещенными, а из волокон, следующих к моторным нейронам, иннервирующим инспираторные мышцы, перекрещены 90%. Моторные нейроны, активированные потоком нервных импульсов инспираторных нейронов дыхательного центра, посылают эфферентные импульсы к нервно-мышечным синапсам мышц вдоха, обеспечивающих увеличение объема грудной клетки. Вслед за грудной клеткой увеличивается объем легких и происходит вдох.
36. Рецепторы лёгких. Их значение в регуляции дыхания. Легочные рецепторы растяжения Основной ответ на возбуждение легочных рецепторов растяжения — уменьшение частоты дыхания в результате увеличения времени выдоха. Эта реакция называется инфляционным (т. е. возникающим в ответ на раздувание) рефлексом Геринга — Брейера. Его можно выявить в экспериментальных условиях, регистрируя у кролика активность участка диафрагмы и не препятствуя при этом деятельности другой дыхательной мускулатуры. Ирритантные рецепторы J-рецепторы 37. Симпатические и парасимпатические влияния на просвет бронхов § Симпатическая иннервация обеспечивает: расширение бронхов, уменьшение выделения слизи в бронхах; § Парасимпатическая иннервация действует следующим образом: сужение бронхов, увеличение слизи в бронхах;
38. Значение центральных и периферических хеморецепторов в регуляции дыхания. Рецепторы, чувствительные к углекислому газу, имеются в каротидных тельцах, аортальных тельцах дуги аорты, а также непосредственно в продолговатом мозге — центральные хеморецепторы. Последние расположены на вентральной поверхности продолговатого мозга в области между выходом подъязычного и блуждающего нервов. Рецепторы углекислого газа воспринимают также изменения концентрации ионов Н+. Рецепторы артериальных сосудов реагируют на изменения рС02 и рН плазмы крови, при этом поступление к инспираторным нейронам афферентных сигналов от них возрастает при увеличении рСО2, и (или) снижении рН плазмы артериальной крови. В ответ на поступление от них большего числа сигналов в дыхательный центр рефлекторно увеличивается вентиляция легких за счет углубления дыхания. Центральные хеморецепторы реагируют на изменения рН и рСО2, ликвора и межклеточной жидкости продолговатого мозга. Считают, что центральные хеморецепторы преимущественно реагируют на изменение концентрации протонов водорода (рН) в интерстициальной жидкости. При этом изменение рН достигается вследствие легкого проникновения углекислого газа из крови и ликвора через структуры гематоэнцефалического барьера в мозг, где в результате его взаимодействия с Н20 образуется углекислота, диссоциирующая с высвобождением прогонов водорода. Сигналы от центральных хеморецепторов также проводятся к инспираторным нейронам дыхательного центра. Некоторой чувствительностью к сдвигу рН интерстициальной жидкости обладают сами нейроны дыхательного центра. Снижение рН и накопление углекислого газа в ликворе сопровождается активацией инспираторных нейронов и увеличением вентиляции легких.
39. Влияние перерезок ствола и спинного мозга на разных уровнях на характер дыхания 1. При нанесении перерезки выше моста характер дыхания не изменяется. Отделение продолговатого мозга от спинного приводит к полной остановке дыхания. Таким образом, за чередование вдоха и выдоха отвечает "центр", расположенный в нижних отделах ствола мозга. Этот отдел продолжает ритмично посылать импульсы к дыхательным мышцам даже после того, как перекрываются все идущие к нему афферентные каналы. 2. Если нанести перерезку на границе между верхней и средней третью моста и одновременно пересечь оба блуждающих нерва, то дыхание остановится в фазе вдоха, лишь иногда прерываемой экспираторными движениями. Такой тип дыхания называется апнейзис. Очевидно, что после подобной перерезки устраняются тормозные влияния верхних отделов моста на нейроны, отвечающие за вдох. 3. После перерезки ствола мозга ниже моста дыхательные движения сохраняются, однако их ритм становится неправильным. Иногда возникает дыхание типа гаспинга, при котором длительный выдох периодически прерывается короткими вдохами.
40. Влияние перерезок блуждающих и симпатических нервов на характер дыхания После перерезки или фармакологической блокады определённых вегетативных нервов иннервируемые органы лишаются их тонического влияния и такая утрата сразу же обнаруживается. После такого вмешательства деятельность рабочих органов изменяется под влиянием сохранившего с ним связь отдела вегетативной нервной системы. Дыхательный центр вызывает сокращения дыхательной мускулатуры и после перерезки блуждающих нервов, т. е. после выключения рефлекторной саморегуляции дыхания. При перерезке обоих блуждающих нервов дыхание сохраняется, но оно становится более глубоким и редким. Углубление дыхания после перерезки блуждающих нервов происходит благодаря тому, что прерываются афферентные волокна, вызывающие торможение дыхательного центра. Раздражение центрального конца перерезанного блуждающего нерва во время вдоха вызывает угнетение вдыхательных движений и задержку дыхания на выдохе.
41. Особенности регуляции дыхания во время сна и бодрствования Дыхание во время сна контролируется менее строго, чем во время бодрствования; в то же время сон оказывает сильнейшее воздействие на параметры дыхания, в первую очередь на чувствительность хеморецепторов к APco2 и на ритм дыхания. ■ В течение фазы «медленного» сна ритм дыхания в целом становится более регулярным, чем в состоянии бодрствования, но чувствительность хеморецепторов к APco2 снижается, как и эфферентные влияния на дыхательные мышцы и мышцы глотки. ■ Во время фазы «быстрого» сна происходит дальнейшее снижение чувствительности к APco2, но ритм дыхания становится нерегулярным (вплоть до отсутствия всякого ритма). ■ Заметной чертой фазы быстрого сна является общее снижение мышечного тонуса. Мышцы гортани, глотки и языка участвуют в расслаблении, которое может обусловить обструкцию верхних дыхательных путей. Частым последствием сужения дыхательных путей во время сна является храп. 42. Регуляция дыхания при мышечной работе Интенсивность дыхания тесно связана с интенсивностью окислительных процессов: глубина и частота дыхательных движений уменьшаются в покое и увеличиваются при работе, притом тем сильнее, чем напряженнее работа. Мышечная работа всегда сопровождается увеличением легочной вентиляции, что совершенно необходимо для удовлетворения возникающей при работе потребности в кислороде. При интенсивной работе легочная вентиляция может достигать 120 л/мин вместо 5-8 л/мин в покое. Одна из закономерностей корковой регуляции дыхания в процессе тренировки человека к новому для него виду мышечной деятельности выражается в том, что при этом устанавливается характер дыхания, адэкватный данному виду мышечной деятельности. Следовые явления после работы сохраняются и фиксируются в коре головного мозга как определенная стереотипная комбинация, ведающая координацией функций, в том числе и приспособлением дыхания к тому именно виду мышечной деятельности, которая выполняется повторно. 43. Определение обмена энергии методом Дугласа и Холдена открытый способ непрямой калориметрии - способ Дугласа — Холдейна, при котором в течение 10—15 мин собирают выдыхаемый воздух в мешок из воздухонепроницаемой ткани (мешок Дугласа), укрепляемый на спине обследуемого. Он дышит через загубник, взятый в рот, или резиновую маску, надетую на лицо. В загубнике и маске имеются клапаны, устроенные так, что обследуемый свободно вдыхает атмосферный воздух, а выдыхает воздух в мешок Дугласа. Когда мешок наполнен, измеряют объем выдохнутого воздуха, в котором определяют количество О2 и СО2. Объем выдохнутого воздуха определяется с помощью газовых часов. Из мешка берут пробу воздуха и определяют в ней содержание кислорода и углекислого газа; вдыхаемый воздух содержит определенное их количество. Отсюда по разности в процентах вычисляют количество потребленного кислорода, выделенного углекислого газа и дыхательный коэффициент. Затем находят соответствующий его величине тепловой эквивалент кислорода, который умножают на количество литров потребленного кислорода. При этом получают величину обмена за тот промежуток времени, в течение которого производилось определение газообмена. Затем переводят эту величину на сутки. 44. Дыхательный коэффициент. Значения при окислении в организме Б, Ж. У. Дыхательным коэффициентом (ДК) называется отношение объема выделенного СО2 к объему поглощенного О2. Дыхательный коэффициент различен при окислении белков, жиров и углеводов. С6Н12О6 + 6 О2 = 6 СО2 + 6 Н2О При окислении глюкозы число молекул образовавшегося СО2 равно числу молекул затраченного (поглощенного) О2. Следовательно, дыхательный коэффициент (отношение СО2/О2) при окислении глюкозы и других углеводов равен 1. При окислении жиров и белков дыхательный коэффициент будет ниже единицы. При окислении жиров дыхательный коэффициент равен 0,7. Аналогичный расчет можно сделать и для белка; при его окислении в организме дыхательный коэффициент равен 0,8. При смешанной пище у человека дыхательный коэффициент обычно равен 0,85—089. Определенному дыхательному коэффициенту соответствует определенный калорический эквивалент кислорода.
45. Калорический эквивалент кислорода. Калорический эквивалент кислорода – показывает сколько тепла выделилось, если организм потребил 1 л О2. Если мы знаем, сколько О2 использовал организм и знаем, какие в-ва (Б, Ж или У) окислились, то мы можем посчитать энергетические затраты. (т. е. сколько тепла, освобождается при полном окислении какого-либо вещества до СО2 и Н2О на каждый литр поглощенного кислорода) Калорический эквивалент кислорода при окислении белков равен 4,8 ккал (20,1 кДж), жиров - 4,7 ккал (19,619 кДж), углеводов - 5,05 ккал (21,2 кДж). 46. Основной обмен энергии, его значение. Основной обмен - это минимальные для бодрствующего организма затраты энергии, определенные в строгих стандартных условиях: - в положении лежа, при полном мышечном и эмоциональном покое (т. к. мышечное и эмоциональное напряжение значительно повышают энерготраты); - натощак, через 12-16 часов после последнего приема пищи (чтобы исключить специфическое-динамическое действие пищи); - при внешней температуре комфорта - 18-20 градусов тепла (температура выше или ниже этих цифр может значительно изменить - увеличить или уменьшить - энерготраты); Величина основного обмена в среднем составляет 1 ккал в 1 ч на 1 кг массы тела. У мужчин в сутки основной обмен равен 1700 ккал, у женщин - на 10% ниже. Энергия в условиях основного обмена идет на то, чтобы клетки поддерживали необходимый для жизни минимальный уровень ОВ процессов, чтобы работали постоянно сердце, почки, печень, дыхательные мышцы, нужно поддерживать мышечный тонус. 47. Изменение величин основного обмена в онтогенезе. Основной обмен у детей интенсивнее, чем у взрослых, так как на единицу массы у них приходится относительно большая поверхность тела, и процессы диссимиляции, а не ассимиляции являются преобладающими. Энергетические затраты на рост тем больше, чем моложе ребенок. Так что расход энергии, связанный с ростом, в возрасте 3 месяцев составляет 36%, в возрасте 6 мес. - 26%, 9 мес. - 21 % общей энергетической ценности пищи. У человека среднего возраста основной обмен составляет 4187 Дж на 1 кг массы в 1 ч. В среднем это 7-7,6 МДж в сутки. При этом для каждого человека величина основного обмена относительно постоянная. 48. Факторы, от которых зависит величина основного обмена энергии. Возраст, пол, рост, масса тела, наличие заболеваний (при некоторых эндокринных заболеваниях)
- соотношение между количеством энергии, поступающей с пищей и энергии, расходуемой организмом. В регуляции энергетического обмена участвуют центральные и периферические механизмы. Центральный контроль реализуется с участием эффекторных систем головного мозга (нейропептиды, моноамины, эндоканнабиноиды и др.). Периферическая регуляция обеспечивается сигналами, исходящими от гормонов, характеризующих объём жировых депо (лептин, инсулин), ряда других гормонов желудочно-кишечного тракта (глюкагоноподобный пептид-1, грелин и т.д.), нутриентов (глюкоза, свободные жирные кислоты и др.). На основании анализа поступившей информации формируются адекватные реакции, направленные на обеспечение долгосрочного контроля (поддержание массы тела) и краткосрочного контроля (инициация либо прекращение приема пищи) гомеостаза 50. Калорическая ценность питательных веществ. Калорийность пищи - содержание свободной энергии обеспечивается наличием в ней белков, жиров и углеводов. Зная состав пищевых продуктов и их усвояемость, можно вычислить энергетическую ценность принятой пищи, используя так называемые калорические коэффициенты питательных веществ. Калорическим, или тепловым, коэффициентом называют количество тепла, освобождаемое при сгорании 1 г. вещества. Калорические коэффициенты основных питательных веществ при окислении их в организме таковы: 1г. белка 17,17 кДж (4,1 ккал); 1г. жира 38,94 кДж (9,3 ккал); 1г. углеводов 17,17 кДж (4,1 ккал). Определение этих коэффициентов производят с помощью калориметрической бомбы Бертло. При определении калорийности различают величину «брутто» и «нетто». Калорийность «брутто» - общая калорийность принятой пищи, калорийность «нетто» вычисляется с поправкой на усвояемость. Не вся принятая пища усваивается, т.е. всасывается из пищеварительного тракта и используется в организме; часть пищи покидает кишечник в виде шлаков. 51. Специфическое динамическое действие пищи. После приема пищи интенсивность обмена веществ и энерготраты организма увеличиваются по сравнению с их уровнем в условиях основного обмена. Увеличение обмена веществ и энергии начинается через час, достигает максимума через 3 ч после приема пищи и сохраняется в течение нескольких часов. Влияние приема пищи, усиливающее обмен веществ и энергетические затраты, получило название специфического динамического действия пищи.
При белковой пище оно наиболее велико: обмен увеличивается в среднем на 30 %. При питании жирами и углеводами обмен увеличивается у человека на 14—15 %.
52. Центры терморегуляции, последствия их разрушения. Калорические коэффициенты Б, Ж, У. Центр терморегуляции находится в гипоталамусе. Передний отдел гипоталамуса воспринимает информацию от периферических и центральных терморецепторов. Центр теплопродукции расположен в ядрах заднего отдела гипоталамуса. В терморегуляторном центре гипоталамуса обнаружены различные по функциям группы нервных клеток: 1) термочувствительные нейроны преоптической области; 2) клетки, «задающие" уровень поддерживаемой в организме температуры тела («установочная точка» терморегуляции) в переднем гипоталамусе; 3) вставочные нейроны (интернейроны) гипоталамуса; 4) эффекторные нейроны, управляющие процессами теплопродукции и теплоотдачи, в заднем гипоталамусе. При разрушении центра терморегуляции в гипоталамусе гомойотермных животных превращается в пойкилотермные. Калорические коэффициенты основных питательных веществ при окислении их в организме таковы: 1г. белка 17,17 кДж (4,1 ккал); 1г. жира 38,94 кДж (9,3 ккал); 1г. углеводов 17,17 кДж (4,1 ккал). 53.Центры теплообразования и теплоотдачи, последствия их разрушения. Центр терморегуляции находится в гипоталамусе. Передний отдел гипоталамуса воспринимает информацию от периферических и центральных терморецепторов. Центр теплопродукции расположен в ядрах заднего отдела гипоталамуса. Разрушение этих отделов гипоталамуса или нарушение их нервных связей посредством перерезки на уровне среднего мозга в экспериментах на животных приводит к нарушению контроля за температурой тела у гомойотермных организмов 54. Термоиндифферентная зона. Изменение её величины в постнатальном онтогенезе. Термоиндифферентная зона - температура окружающей среды, при которой теплопродукция организма минимальна (~28 °С - для обнаженного человека, ~ 18-20 °С - для легко одетого). для новорожденных детей составляет 33 °С В связи с большой относительной поверхностью тела и более напряженной терморегуляцией границы термоиндифферентной зоны в раннем возрасте сдвигаются в сторону более высоких температур и составляют от 19 до 24 °С. 55. Центральные терморецепторы. Развитие центральных терморецепторов в онтогенезе. Центральные рецепторы содержатся в гипоталамусе (всего в передоптичний зоне). Некоторое их количество находится в шейно-грудном отделе спинного мозга, в мышцах абдоминальной зоны. Здесь также содержатся оба типа рецепторов (преимущественно тепловые). Эти рецепторы играют основную роль в регулировании теплообмена, потому что они контролируют температуру ядра. В целом терморегуляция новорожденного значительно несовершеннее, чем в зрелом возрасте. Однако активно функционируют периферические и центральные терморецепторы, центр терморегуляции гипоталамуса. С возрастом терморегуляторные механизмы совершенствуются. Увеличивается эффективность потоотделения, появляется способность к дрожательному термогенезу, возрастает значимость рефлекторных механизмов поддержания температурного гомеостаза. К 15-16 годам механизмы терморегуляции в основном соответствуют зрелому организму. 56. Суточный расход энергии. Зависимость суточного расхода энергии от возраста и уровня физической активности. Суточный расход энергии у здорового человека значительно превышает величину основного обмена и складывается из следующих компонентов: - основного обмена; - рабочей прибавки, т. е. энергозатрат, связанных с движением и с выполнением той или иной работы; - специфического-динамического действия пищи - увеличения интенсивности обмена веществ и энергозатрат, связанных с приемом пищи, процессами пищеварения и всасывания. I группа - работники умственного труда - 2800-3000 ккал; - II группа - работники механизированного труда и сферы обслуживания - 3000-3500 ккал; - III группа - работники умеренно тяжелого труда, связанного со значительными физическими усилиями - 3500-4000 ккал; - IV группа - работники тяжелого, немеханизированного труда - 4000-4500 ккал; - V группа - работники очень тяжелого физического труда - 4500-5000 ккал.
57. Требования к соотношению питательных веществ в пищевых рационах. - энергетическая достаточность - достаточность и сбалансированность поступления Б, Ж, У - соотношения в рационе Б:Ж:У=1:1(1,2):4 -достаточность содержания витаминов и минеральных солей - кратность приёма пищи и % распределение приёма пищи
58. Способы теплоотдачи, её регуляция. Излучение — это способ отдачи тепла в окружающую среду поверхностью тела человека в виде электромагнитных волн инфракрасного диапазона (а = 5—20 мкм). Количество тепла, рассеиваемого организмом в окружающую среду излучением, пропорционально площади поверхности излучения и разности средних значений температур кожи и окружающей среды. Теплоотдача путем излучения возрастает при понижении температуры окружающей среды и уменьшается при ее повышении. В условиях постоянной температуры окружающей среды излучение с поверхности тела возрастает при повышении температуры кожи и уменьшается при ее понижении. Теплопроведение — способ отдачи тепла, имеющий место при контакте, соприкосновении тела человека с другими физическими телами. Количество тепла, отдаваемого организмом в окружающую среду этим способом, пропорционально разнице средних температур контактирующих тел, площади контактирующих поверхностей, времени теплового контакта и теплопроводности контактирующего тела. Конвекция — способ теплоотдачи организма, осуществляемый путем переноса тепла движущимися частицами воздуха (воды). Для рассеяния тепла конвекцией требуется обтекание поверхности тела потоком воздуха с более низкой температурой, чем температура кожи. При этом контактирующий с кожей слой воздуха нагревается, снижает свою плотность, поднимается и замещается более холодным и более плотным воздухом. испарение — это способ рассеяния организмом тепла в окружающую среду за счет его затраты на испарение пота или влаги с поверхности кожи и влаги со слизистых оболочек дыхательных путей («влажная» теплоотдача). У человека постоянно осуществляется выделение пота потовыми железами кожи («ощутимая», или железистая, потеря воды), увлажняются слизистые оболочки дыхательных путей («неощутимая» потеря воды) Если внешняя температура превышает среднее значение температуры кожи, то организм не может отдавать во внешнюю среду тепло излучением, конвекцией и теплопроведением. Организм в этих условиях начинает поглощать тепло извне, и единственным способом рассеяния тепла становится усиление испарения влаги с поверхности тела. Такое испарение возможно до тех пор, пока влажность воздуха окружающей среды остается меньше 100 %. При интенсивном потоотделении, высокой влажности и малой скорости движения воздуха, когда капли пота, не успевая испариться, сливаются и стекают с поверхности тела, теплоотдача путем испарения становится менее эффективной 59. Транзиторная лихорадка у младенцев, причины и последствия. Одновременно с физиологической потерей в весе на 3-4 день жизни у новорожденного возникает транзиторная лихорадка. Причин этому переходному состоянию несколько. Это недостаточное поступление в организм ребенка жидкостей (в это время кроха еще получает молозиво, которое намного гуще «зрелого» молока и содержит большое количество белка). Также у малыша еще не отлажена система терморегуляции и кожа испаряет больше жидкости, чем организм получает с пищей и питьем. В результате возникает сухость кожи, слизистых и общее обезвоживание организма. Проявляется транзиторная лихорадка повышением температуры тела до 38-40 градусов, ребенок становится возбужденным и беспокойным. Такое состояние продолжается 1-2 суток, а затем температура нормализуется.
60. Механизмы поддержания постоянства температуры организма при воздействии низких и высоких температур. Высокая температура вызывает интенсивное потоотделение Низкие температуры рефлекторно уменьшается теплоотдача и усиливается теплопродукция. Уменьшение теплоотдачи происходит за счёт спазма (сужения) сосудов, увеличения термического сопротивления тканей организма. Рост теплопродукции при охлаждении достигается усилием окислительных обменных процессов в организме (понижение температуры тела на 1°С сопровождается приростом обменных процессов на 10°С). Терморегуляцию можно разделить на два основных вида: 1. Физическая терморегуляция: – Испарение (потоотделение); – Излучение (радиация); – Теплопроведение (кондукция); – Конвекция. 2. Химическая терморегуляция. – Сократительный термогенез; – Несократительный термогенез.
Дата добавления: 2016-06-06 | Просмотры: 955 | Нарушение авторских прав |