|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Регуляция фотосинтеза на уровне растительного организмаВыделяют два механизма регуляции на этом уровне: 1) Гормональный механизм регуляции проявляется через воздействие фитогормонов на скорость реализации генетической информации. Цитокинин увеличивает скорорсть деления хлоропластов. АБК индуцирует закрытие устьиц, тем самым, ингибируя фотосинтез. 2) Концентрационный механизм регуляции сводится к соотношению мощности органов, производящих ассимиляты и потребляющих их. Существует контрольный уровень содержания метаболитов в хлоропластах, изменение которого ведет к усилению или, наоборот, к угнетению фотосинтеза. Органы-потребители ассимилятов расходуют их на эпигенез или же образуют резервные фонды веществ. Этот тип регуляции тесно связан с онтогенезом растений. 3) 3.5.4. Экология фотосинтеза Под экологией фотосинтеза понимают зависимость его интенсивности от факторов внешней среды (концентрации углекислого газа и кислорода, температура, водный режим тканей листа, минеральное питание, интенсивность и спектральный состав света). Существуют понятия истинного и кажущегося (наблюдаемого) фотосинтеза. Кажущийся фотосинтез – это результат сопряженности трех процессов: фотосинтеза, фотодыхания и клеточного дыхания. Интенсивность фотосинтеза можно определить по количеству органического вещества, образованного за единицу времени. При использовании газометрического метода интенсивность фотосинтеза выражают в миллиграммах углекислого газа, поглощенного единицей площади листа за 1 час (мг СО2/1 дм2/1 час) или в миллиграммах выделенного кислорода (мг О2/1 дм2/1 час).
Интенсивность фотосинтеза С3-растений составляет от 5 до 25 мг/дм2/час, С4-растений – 80 мг/дм2/час. Для оценки продуктивности экосистем используют понятия первичной, вторичной и валовой продуктивности. Под первичной продуктивностью понимают продуктивность всех хлорофиллсодержащих организмов. Вторичная продуктивность – продуктивность других трофических уровней. Валовая продуктивность – количество вещества, произведенного в единицу времени трофическим уровнем. Зависимость фотосинтеза от света Большая часть (60%) попадающего на лист солнечного излучения не может участвовать в фотохимических процессах, поскольку имеет длину волны, которая не поглощается пигментами листа. Из оставшихся 40% фотосинтеически активных лучей солнечного спектра длиной волны 400 – 700 нм, часто называемых фотосинтетически активной радиацией (ФАР), 8% отражается, 8% – рассеивается в виде тепла, 19% идет на испарение воды, и только 5% – на фотосинтез. Свет оказывает разностороннее влияние на фотосинтез. Во-первых, он является источником энергии, а во-вторых, оказывает влияние на состояние устьичного аппарата.
Световое насыщение достигается при освещенности, равной примерно половине полной солнечной освещенности. У растений С4-типа световое насыщение происходит при более высокой степени освещенности (= 60 тыс. люксов). Уровень освещенности, при котором количество поглощенного и выделенного СО2 равно, называется световым компенсационным пунктом. Световые реакции происходят очень быстро и кратковременное изменение освещенности влияет на интенсивность фотосинтеза. На интенсивность фотосинтеза также влияет и спектральный состав света, об этом судят по спектру действия света. Спектр действия – это зависимость эффективности биологического действия света от длины его волны. Максимальная интенсивность фотосинтеза достигается при освещении растения красным светом. При выращивании растений на синем свету образуются неуглеводные компоненты (аминокислоты и другие органические кислоты). Больше углеводных продуктов образуется при увеличении освещенности. Видимо, есть регулятор, реагирующий на качество и количество света, который переключает метаболические пути. Возможно, что это флавиновые соединения.
Оптимальное для С3 и С4-растений содержание углекислого газа равно 0,03%. Часть СО2 поглощается из почвы. Внесение в почву удобрений положительно на интенсивность фотосинтеза. В растениях поглотителями СО2 являются монокарбрновые органические кислоты, которые, ассимилируя углекислоту, превращаются в дикарбоновые. Содержание СО2 в атмосфере подвержено годичным и суточным изменениям. В течение дня концентрация углекислоты в воздухе снижается на 12%. Зимой концентрация углекислого газа выше, чем в другие сезоны года. Зависимость фотосинтеза от содержания кислорода Снижение содержания кислорода в атмосфере приводит к увеличению интенсивности фотосинтеза (эффект Варбурга). Увеличение концентрации О2 приводит к тому, что фермент РУБИСКО начинает проявлять оксигеназную активность и начинается более интенсивное фотодыхание. Цикл Кальвина у С4-растений без этапа карбоксилирования не происходит. Избыток кислорода приводит к тому, что оксидоредуктаза, восстанавливающая НАДФ+, начинает восстанавливать О2 до воды – происходит псевдоциклическое фотофосфорилирование, в результате которого интенсивность фотосинтеза снижается. Кислород непосредственно снижает активность РДФ-карбоксилазы (присоединение молукулы О2 происходит не к активному центру, а к аллостерическому). Снижение концентрации кислорода до 5 – 7% увеличивает скорость фотосинтеза Зависимость фотосинтеза от минерального питания Минеральное питание необходимо растениям для формирования фотосинтетического аппарата. Калий влияет на состояние устьичного аппарата (влияет на степень обводненности замыкающих клеток). Зависимость фотосинтеза от воды Вода имеет как прямое, так и косвенное влияние на фотосинтез. Косвенное влияние заключается во влиянии содержания воды на состояние устьичного аппарата. Водный дефицит влияет на изменение конформации электронных переносчиков в сторону снижения их способности к переносу электронов.
Высокие температуры вызывают денатурацию белков, тем самым, ингибируя фотосинтез. Оптимальной для процесса фотосинтеза температурой является 25 – 35°C. У С3-растений с увеличением температуры квантовый выход фотосинтеза снижается (из-за инактивации фотодыхания), у С4-растений – не изменяется (при температуре от 12 до 40°C). Все факторы среды влияют на протекание фотосинтеза в комплексе. Суточный ход фотосинтеза отражает суточную динамику интенсивности света. Разные процессы по-разному изменяются при действии одного фактора. Так например, при увеличении температуры интенсивность фотосинтеза снижается, а дыхание активируется. Интенсивность света максимальна в предполуденные часы (≈ в 12 часов). В самые жаркие часы происходит кратковременное снижение фотосинтетической активности, т. к. у растений в это время возникает водный дефицит и они закрывают устьица. В послеполуденные часы наступает второй максимум фотосинтетической активности. Сезонная динамика фотосинтеза определяются как климатическими условиями, так и онтогенезом. В ходе онтогенеза происходит перестройка фотосинтетического аппарата, этот связано с ростом, развитием и старением листа. На начальных стадиях онтогенеза листья сами потребляют ассимиляты, затем они становятся донорами ассимилятов. Листья не обмениваются друг с другом ассимилятами, а стареющие листья поставляют ассимиляты и продукты распада в соседние листья и гетеротрофные органы. ТЕМА 4. Дыхание растений Дыхание – это процесс биологического окисления органических молекул, в результате которого высвобождается энергия и образуются химически активные метаболиты. Это катаболический процесс и его протекание ведет к увеличению энтропии и выделению свободной энергии, которая необходима для биосинтетических процессов, для обеспечения активного транспорта через мембраны и для клеточных движений. Для обеспечения этих процессов энергией происходит поэтапное ее высвобождение, поскольку залповое высвобождение энергии для живых организмов (как при горении) является разрушающим. Высвобождающаяся энергия запасается в АТФ. АТФ следует рассматривать как носитель свободной энергии, а не как форму запасания энергии. При трансформации энергии химических связей органических молекул в энергию макроэргических связей около 60% энергии рассеивается в виде теплоты, конкретное значение КПД при этом зависит от пути дыхания. Включение того или иного пути дыхания зависит как от внутренних (наличие того или иного субстрата), так и от внешних (наличие или отсутствие кислорода) причин. Для биосинтеза некоторых молекул требуется наличие не только АТФ, но и восстановительного эквивалента, поэтому один из путей дыхания (пентотзофосфатный) является поставщиком НАДФН2. Биологическая роль процессов дыхания связана не только с обеспечением организма энергией, но и с образованием химически активных метаболитов, в роли которых выступают разнообразные промежуточные соединения. Например органические кислоты цикла Кребса используются для синтеза аминокислот; биосинтез ароматических аминокислот (фен, три, тир) связан с шикимовой кислотой, которая образуется из метаболитов пентозофосфатного пути; предшественником пуринов также является один из метаболитов процессов дыхания – сукцинил-КоА. Т. Соссюр, используя количественные методы анализа, установил, что в темноте растение поглощает столько кислорода, сколько выделяет углекислого газа. Отношение количества молекул СО2, выделяемого при дыхании, к количеству поглощенных молекул кислорода называют дыхательным коэффициентом (ДК). Оказалось, что ДК не всегда равен единице, а только в случае, если дыхательным субстратом являются углеводы. Это следует из общего уравнения дыхания: С6Н12О6 + 6О2 → 6СО2 + 6Н2О Субстратом дыхания может быть любое органическое соединение. Например, катаболизм стеариновой кислоты протекает по следующей схеме: С17Н35СООН + 26О2 → 18СО2 + 18Н2О ДК = 18/26 ≈ 0,7
ДК = 4 Величина ДК зависит от степени восстановленности субстрата. Выделяющиеся в процессе дыхания газы можно учитывать при помощи аппарата Варбурга. На основании этого учета можно вычислить ДК и предположить, что именно является дыхательным субстратом в данный момент. В истории изучения дыхания было два подхода. Сторонники одного из них считали, что суть дыхания сводится к активизации кислорода; а другого – к активизации водорода. Это противоречие было устранено в 1925 г. в связи с открытием фермента цитохромоксидазы. Этот фермент снимает электроны с атомов водорода непосредственно на кислород, т. е. в процессе дыхания происходит активизация и кислорода и водорода. 4.1. Гликолиз (анаэробная фаза аэробного дыхания) Углеводы по степени окисленности-восстановленности занимают промежуточное положение. Конкретно субстратом дыхания являются моносахариды. Включение глюкозы в катаболические процессы предваряется ее активацией с помощью киназ. Гликолиз протекает в цитоплазме, строме хлоропластов, матриксе митохондрий. Это кислородонезависимый процесс. Процесс гликолиза делят на 3 этапа: 1) подготовительный этап; 2) первое субстратное фосфорилирование; 3) второе субстратное фосфорилирование. Подготовительный этап гликолиза (схематично)
I субстратное фосфорилирование
II субстратное фосфорилирование
Суммарное уравнение процесса гликолиза:
С6Н12О6 + 2АДФ + 2Фн + 2НАД+ → 2С3Н4О3 + 2АТФ + 2НАДН2
Дальнейшее превращение ПВК может протекать разными путями:
Второй фазой аэробного дыхания считается процесс окислительного декарбоксилирования пирувата. Этот процесс катализируется пируватдегидрогеназным комплексом, который в своем составе имеет 5 различных коферментов. В суммарном виде окислительное декарбоксилирование ПВК выглядит следующим образом:
Цикл Кребса Третьей фазой аэробного дыхания является цикл Кребса, локализованный в матриксе митохондрий. Это кислородозависимый процесс, в ходе которого ацетильный компонент полностью окисляется до СО2. Схема реакций цикла Кребса Суммарное уравнение цикла Кребса:
АцКоА + 3Н2О + 3НАД+ + ФАД + АДФ + Фн → 2СО2 + 3НАДН2 + ФАДН2 + АТФ 4.3. Окислительное фосфорилирование НАД и ФАД содержат по паре электронов с высоким потенциалом переноса, при этом высвобождается энергия, которая идет на синтез АТФ. Под окислительным фосфорилировнием понимают процесс синтеза АТФ, сопряженный с переносом электронов по цепи переносчиков, который осуществляется во внутренней митохондриальной мембране. В процессе обратного тока электронов за счет АТФ-синтазного комплекса, при этом идет сопряжение процессов окисления и фосфорилирования. Движущей силой этих процессов является потенциал переноса электронов, присущий дегидгореназам.
Δσ°НАД = – 2 · 23 (0,82 – (– 0,32)) = – 52,6 ккал Δσ°ФАД = – 2 · 23 (0,82 – (– 0,04)) = – 39,8 ккал Изменения свободной энергии показывают число остатков Фн, включенных в органические молекулы, в расчете на 1 атом потребляемого кислорода. Отличия процесса окислительного фосфорилирования в растительных и животных клетках: 1) Растительные митохондрии содержат НАД-завимую дегидрогеназу, которая находится на наружной поверхности митохондриальной мембраны. Из-за нее идет окисление цитоплазматической НАДН2 (из гликолиза). Т. к. нормальная мембрана проницаема для дегидрогеназ, то не нужны челночные системы как в животных клетках. Подключение НАД-зависимой дегидрогеназы к процессу идет сразу на убихинон. 2) Митохондрии растений имеют два пути переноса электронов. Один из них блокируется цианидом, а другой – цианидустойчивый. Расхождение этих путей идет после убихинона. Механизм переключения не известен. 3) Окисление НАД сопровождается фосфорилированием только одной молекулы АДФ, при этом выделяется большое количество тепла. Этот механизм объясняет образование теплоты в цветках и соцветиях (термогенных тканях) энтомофильных растений. Тепло здесь используется на испарение пахучих соединений. Скорость окислительного фосфорилирования определяется содержанием АДФ. Если клетке не нужна АТФ, то переноса электронов не происходит. Клетке важно полное восстановление кислорода до воды. При неполном восстановлении образуется перекись водорода или О2- (крайне токсичен для клетки, окисляет липиды мембран). Аэробная клетка защищается от этих активных форм кислорода благодаря работе двух ферментов: супероксиддисмутазы (СОД) и каталазы:
Энергетический выход аэробного дыхания 1) Гликолиз: 4АТФ, но 2 из них идут на фосфорилирование субстрата, поэтому общий выход составляет 2АТФ и 2НАДН2 = 4АТФ. Итого 6АТФ. 2) Окислительное декарбоксилирование ПВК: 2НАДН2 = 6АТФ. 3) Цикл Кребса: 6НАДН2 = 18АТФ; 2ФАДН2 = 4АТФ; 2 АТФ. Итого 24АТФ. 4) Окислительное декарбоксилирование (суммарный выход): 36 АТФ. Суммарное уравнение аэробного дыхания:
С6Н12О6 + 6О2 +36АДФ + 36Фн → 6СО2 + 6Н2О + 36АТФ Дата добавления: 2015-12-15 | Просмотры: 1980 | Нарушение авторских прав |


 Зависимость фотосинтеза от содержания углекислоты
Зависимость фотосинтеза от содержания углекислоты
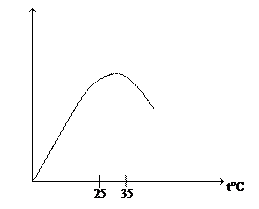 Зависимость фотосинтеза от температуры
Зависимость фотосинтеза от температуры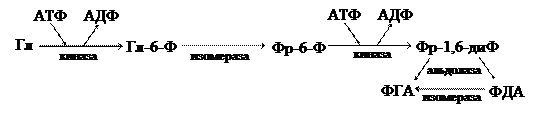
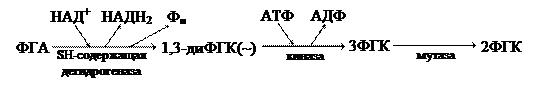

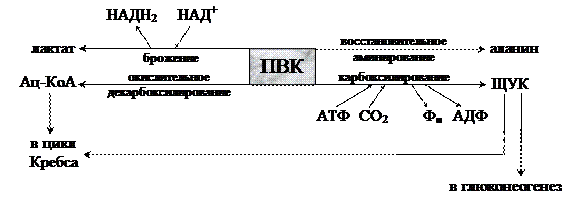
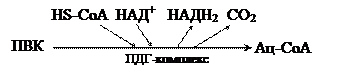
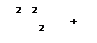 ЕО /Н О = + 0,12 В – электроноакцепторная система;
ЕО /Н О = + 0,12 В – электроноакцепторная система;
 ЕНАДН /НАД = – 0, 32 В
ЕНАДН /НАД = – 0, 32 В ЕФАДН /ФАД = – 0,04 В
ЕФАДН /ФАД = – 0,04 В