|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
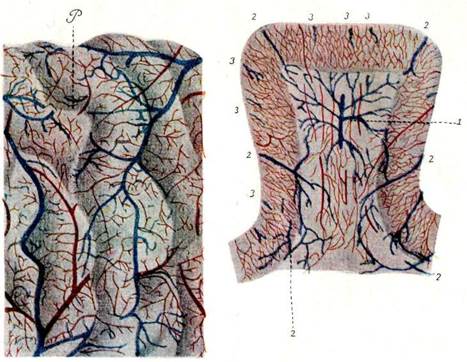
II. Ощущения, восприятия. 9 страницаЕсть основание предполагать, что невроны не представляют собою строго неподвижных образований, а будучи живой тканью, они обладают известной степенью сократительности и как бы амебоидной подвижности; это справедливо по крайней мере по отношению к дендритам, на что указывалось с проведением соответствующих гистологических доказательств Duval’ем136), Demoor 137-138), Stefanowska 139), Querton140), Сухановым 141) и др. По этим авторам, дендриты при известных условиях выпускают по своему протяжению шиловидные, нитевидные и грушевидные отростки, и сами, сокращаясь, могут образовывать ряд утолщений монетовидного или т. наз. монилиформного характера. Значение амебоизма невронов оспаривается, оно далеко еще не выяснено, оспаривается и самый амебоизм; весьма возможно, что он имеет значение при передаче возбуждения с одного неврона на другой посредством контакта. Теория невронов есть одно из крупнейших завоеваний гистологии нервной системы за последнее время; эта теория наилучшим образом об'ясняет физиологические и психологические процессы, происходящие в нервной системе, и потому она стойко держится, несмотря на многочисленные попытки целого ряда весьма солидных исследователей ее опровергнуть. Среди противников теории невронов наиболее видное место занимают Apathy 142-143), Веthе 144), Niss1 125), А. Догель 145-146). Возражения противников сводятся, главнейшим образом, к установлению между невронами наличности анастомозов и к признанию фибрилл не органическими составными частями отдельных невронов, а элементами, лишь проходящими сквозь нервные клетки; последнее далеко еще не представляется доказанным, относительно же анастомозов можно лишь примкнуть к взгляду, высказываемому Verworn'ом147), что невроны взрослых животных во многих случаях связаны между собою теснейшим образом посредством анастомозов, но все-таки они остаются невронами, так как наличность анастомозов не опровергает учения о невронах. Хроматофильное вещество, повидимому, не имеет для неврона значения органической важности, судя по тому, что есть клетки, протоплазма которых почти его не содержит; видимо, значение его питательное. Строение ядра нервной клетки гораздо проще: оно отграничивается от окружающей его протоплазмы отчетливо видимой перепонкой; в центре его помещается ядрышко, энергично впитывающее анилиновые краски; как правило, ядрышко одно, но изредка в ядре бывает два ядрышка и еще реже большее число; в окружности ядрышка видны неправильно расположенные перекладинки, анастомозирующие между собою. Центральные части невронов или нервные клетки различаются не только количеством и расположением хроматофильного вещества, отношением ядра к краскам и количеством протоплазмы, они различаются еще величиною, формою и очертаниями. Эти различия лучше всего выступают при изучении клеточного строения коры головного мозга; различные клетки располагаются в ней слоями, которых большинство исследователей выделяет шесть: поверхностный слой — молекулярный или мелкозернистый, слой малых пирамидных клеток, слой средних и больших пирамидных клеток, зернистый слой, состоящий из мелких клеток, слой глубоких, крупных пирамидных клеток и полиморфный слой, состоящий, как показывает самое название, из разнообразных треугольных и веретенообразных клеток. Отдельные авторы, как, напр., Ramonу Cajal 148), выделяют большее число слоев, различая средние и большие пирамидные клетки, как отдельные слои, различая промежуточные слои, как самостоятельные, но это не имеет существенного значения. Таков общий план расположения клеток в мозговой коре, но расположение слоев и клеток в различных отделах мозговой коры не только не совпадает вполне, но нередко обнаруживает характерные особенности. Так, напр., характерную черту передней центральной извилины составляют гигантские клетки Betz'a и Мержеевского, область cunei характеризуется разделением слоя зерен полоской Vicq d'Azyr'a gyrus cinguli — присутствием в слое глубоких больших пирамид крупных веретенообразных клеток, обонятельная область — значительной толщиной молекулярного слоя и отсутствием малых пирамид и т. д. Brodmann, уделивший много внимания изучению строения мозговой коры, насчитывает до 47 участков ее, имеющих различное строение*) 148-152). Разнообразное строение центральных отделов невронов, нервных клеток, как в смысле их величины, количества протоплазмы и ее состава, величины ядра и его состава (отношение протоплазмы и ядра к краскам), так и в смысле их формы (клетки монополярные, биполярные, мультиполярные и др.), а также разнообразие дендритов и аксонов, в смысле их количества, ветвистости, длины и направления, с несомненностью говорит за то, что биологические функции различных невронов различны: напр., функция гигантских клеток двигательной области мозговой коры теснейшим образом связана с движением. Хотя функции эти до настоящего времени настолько мало изучены, что нельзя высказаться о них иначе, как в виде гипотетических предположений, тем не менее они соответствуют по своему назначению различным сторонам психической жизни; различное строение различных участков мозговой коры также указывает на их различную функцию, что в значительной степени представляется установленным, как путем эксперимента на животных, так и посредством наблюдения и изучения клинического материала. Правда, качества ощущений находятся в непосредственной зависимости от строения периферического воспринимающего нервного аппарата, но роль дальнейшей разработки притекающих к мозговой коре возбуждений принадлежит невронам последней. Кроме нервных элементов, в состав нервной системы, а в частности мозговой коры, входят элементы невроглии. Вопрос о происхождении невроглии долго подвергался сомнению, но в настоящее время, повидимому, надо признать, что невроглийные клетки имеют эпителиальное начало 153а); невроглийные клетки под названием эпендимных клеток выстилают все полости центральной нервной системы; в противоположность эпителиальным элементам, нейробластам, дающим по His'y153) начало невронам, первичные эпителиальные клетки, являющиеся источниками происхождения клеток эпендимы, His называет спонгиобластами. Кроме клеток эпендимы есть, еще другой вид невроглийных клеток, содержащихся в изобилии в сером и белом веществе мозга; эти клетки различной величины, с различным количеством отростков самой разнообразной длины, то тонких и нежных, то грубых, ветвящихся; ветви оканчиваются свободно, без каких бы то ни было анастомозов. Характерная форма клеток невроглии послужила основанием для различных их названий: они известны, как паукообразные клетки, клетки Deiters'a, астроциты и, наконец, просто невроглийные или глиозные клетки. Weigert 154-155), предложивший весьма демонстративный, хотя и с трудом удающийся способ окраски невроглии, высказал мысль, что невроглийные волокна не имеют органической связи с клетками, что они самостоятельны; такое мнение основано, однако, повидимому, на недоразумении, так как по способу Weigert'a не окрашивается протоплазма клеток невроглии; изучение же невроглийных элементов с применением других методов окраски, особенно путем серебрения, а также изучение препаратов, окрашенных по способу самого Weigert'a, но из мозгов с особенно сильным патологическим разростанием невроглийных элементов, как, напр., при прогрессивном параличе помешанных, заставляет не согласиться с мнением Weigert'a и рассматривать невроглийные волокна, как отростки невроглийных клеток. Основное значение невроглии, признаваемое большинством исследователей, есть значение стромы, поддерживающей архитектонику нервной системы, и значение ткани, замещающей элементы нервной системы, в случае их разрушения, напр., при различных болезненных процессах. Но некоторые авторы смотрят на функции невроглии гораздо шире. Так, напр., Golgi и Nansen высказывают предположение, что невроглия является посредствующим звеном между стенками сосудов и окончаниями протоплазматических отростков и служит целям питания невронов. По мнению Andriezen'a 156), невроглия, особенно развитая в окружности сосудов, защищает нервные элементы от чрезмерных приливов крови. Weigert смотрит на невроглию, как на ткань, защищающую нервные элементы от вредных внешних влияний механического характера. Наконец, невроглию рассматривают, как элемент, несущий функцию обмена веществ, развивающий секроторную деятельность, вырабатывающий гормоны 153а). Ramon у Cajal 157-158) различает невроглийные клетки белого вещества, серого и околососудистые; первые являются, в качестве плохого проводника нервных токов, изолирующим слоем и облегчают обращение лимфы; околососудистые клетки, будучи связаны своими отростками с сосудистыми стенками, вызывают посредством сокращения отростков расширение сосудов и таким образом влияют на кровообращение в мозгу, следовательно, и на психические процессы; клетки серого вещества, сокращая и расслабляя свои отростки, т. е., уменьшая или увеличивая их размеры, тем самым способствуют установлению контактов между невронами или прерывают установившиеся контакты. Эта гипотеза, переносящая центр тяжести в функциях центральной нервной системы с невронов на невроглию, уже вследствие одного этого представляется, по справедливому замечанию v. Gehuchten'a *), мало вероятной. В патологических процессах невроглия (ядра клеток глии) участвует в невронофагии. Из всего изложенного вытекает, что функция невроглии еще далеко не выяснена, но повидимому, ее значение далеко не ограничивается структурными и замещающими функциями. Сосудистая система головного мозга характеризуется некоторыми особенностями своего расположения, на которых следует остановиться. Артерии, питающие мозг, большие его полушария, носят название передних, средних и задних мозговых артерий; эти артерии разветвляются в мягкой мозговой оболочке, анастомозируя между собою; в этом согласны все авторы, изучавшие кровообращение в коре мозговых полушарий, но в то время, как одни признают анастомозы между артериями одноименного и противоположного полушарий (Duret158), Те s tut160)), Heubner161) различает целых две сети артериальных анастомозов, поверхностную и глубокую: первая является результатом анастомозов между ветвями мозговых артерий, а вторая образуется анастомозами ветвей, происходящих от поверхностной сети. Так или иначе, от артериальной сети в мягкой оболочке отходят весьма многочисленные нежные артериальные веточки в направлении перпендикулярном поверхности мозга, проникающие в вещество мозга; эти веточки суть питающие мозг артерии в тесном смысле слова; они разделяются на длинные, проникающие в белое вещество мозга и там ветвящиеся, и на короткие, разветвляющиеся в сером веществе; те и другие переходят в капиллярную сеть, те и другие не анастомозируют между собою, будучи концевыми ветвями. Узлы основания головного мозга получают приток питательного материала при посредстве сильвиевых артерий. Мозговые вены в общем соответствуют артериям, но их заметно меньше; вены мозговых полушарий разделяются на вены внутренней, наружной и нижней поверхности полушарий; первые впадают, главным образом, в верхнюю продольную пазуху и только отчасти в нижнюю, в галенову вену и в переднюю мозговую; наружные — в верхнюю продольную пазуху и в пазухи основания, нижние — в боковую пазуху, в переднюю и среднюю мозговые вены, частью в островковую и в верхнюю продольную. Все эти вены богато анастомозируют между собою, образуя в мягкой оболочке густую венозную сеть. Длинные и короткие артерии извилин переходят в капилляры, дающие в свою очередь начало длинным и коротким венам, вливающимся на поверхности мозга в более крупные венозные стволы. Значение крови для организма и в частности для мозга известно, о некоторых же особенностях и значении черепно-мозгового кровообращения будет сказано ниже. Несколько подробнее следует остановиться на черепно-мозговой жидкости, значение которой до сих пор представляется не вполне выясненным. Принимая во внимание новейшие исследования 162-163), посвященные изучению ее происхождения, обращения в центральной нервной системе и ее значения для мозга, надо представить положение дела в следующем виде. Головной мозг одет тремя оболочками — твердой, паутинной и мягкой; в мягкой оболочке разветвляются сосуды, она плотно покрывает мозг, следуя за всеми его изгибами, проникает не только в углубления и борозды, но дает даже влагалища сосудам, направляющимся в вещество мозга; паутинная оболочка (arachnoidea), напротив, не следует за мягкой в глубину борозд, а проходит над углублениями мозга, благодаря чему в таких местах получаются т. наз. подпаутинные пространства; из них более значительные — cisterna magna, расположенная между мозжечком и продолговатым мозгом и сообщающаяся с четвертым желудочком, cisterna basalis, находящаяся на основании мозга и варолиева моста, дающая продолжения вперед и в стороны к сильвиевым ямкам (cicternae fossae Sylvii**)); в этих цистернах или подпаутинных пространствах и в мозговых желудочках (сейчас не идет речь о спинном мозге) находится черепно-мозговая жидкость. Тщательные исследования, клинические и экспериментальные, особенно последние, основанные на выделении впрыснутого животным уранина, окрашивающего ткани в желтый цвет (Kafka*)), заставили прийти к выводу, что черепно-мозговая жидкость не является трансудатом крови, как думали многие авторы, а что она есть продукт деятельности, секрет сосудистого сплетения, plexus chorioideus, главнейшим образом; некоторое участие в выработке этого секрета при нормальных условиях принимает эпендима желудочков и центрального канала. При нормальных условиях жидкость имеет присущий ей состав, приближающийся к составу жидкости передней глазной камеры, при патологических же условиях состав ее меняется, так как к ней примешиваются воспалительные эксудаты. Движение черепно-мозговой жидкости происходит в направлении из желудочков в подпаутинные пространства, из боковых желудочков и третий, через сильвиев водопровод в четвертый, откуда, через, отверстие Magendie и Luschka в подпаутинные пространства головного и спинного мозга; далее жидкость проникает в периваскулярные пространства вен и в самые вены, а также через оболочки нервов (Nervenscheiden) в лимфатические пространства; она находится в перицеллюлярных, околоклеточных пространствах. Движение жидкости чрезвычайно медленное, оборот ее совершается от 6—7 раз в течение суток (Mestrezat), главнейшими двигателями являются дыхательные и пульсовые колебания. На черепно-мозговую жидкость долго смотрели, как на элемент исключительно физического, механического значения; уже позднее на нее стали смотреть, как на жидкость, в которую уносятся продукты метаморфоза нервной ткани, ей было отведено важное место в процессе обмена веществ в организме. Это значение за черепно-мозговой жидкостью сохранилось и до настоящего времени, но в связи с новейшими исследованиями и открытиями значение ее существенно расширилось; уже самый источник ее происхождения, секреция plexus chorioideus, заставляет вывести ее роль далеко за пределы только посредствующей среды; ее способность вырабатывать при известных условиях бактериеубивающие вещества, ферменты, удерживать одни вещества, выделяя другие, говорит за самостоятельность ее функций; ее богатство ферментами при заболеваниях центральной нервной системы, присутствие в ней иммунных веществ, особенно при заболевании мозговых оболочек, заставляют признать, что ей принадлежит защитительная роль при инфекционных поражениях нервной системы. Принимая во внимание новейшие реакции с черепно-мозговой жидкостью (Wassermann'а, Nonne - Аре1t'a и др.), надо полагать, что значение ее для организма еще далеко не выяснено. После приведенного здесь очерка элементов, входящих в состав центральной нервной системы, необходимо установить, каким образом происходит сообщение центральной нервной системы с периферией и каким образом устанавливается связь между различными отделами головного мозга. В анатомии нервной системы различаются двоякого рода связи или системы, служащие указанным целям; они известны под названием проекционных или проводниковых систем или путей и ассоциационных или сочетательных; первые включают в себя пути, служащие для передачи возбуждений от центров к периферии, т. е. двигательные, центробежные системы, и для передачи раздражений, действующих на воспринимающие аппараты органов чувств, от периферии к центрам, т. е., чувствительные, центростремительные системы; вторые включают в себя пути, связывающие между собою соседние извилины, отдельные области одного и того же полушария и области противоположных полушарий головного мозга. Ход и развитие проекционных систем подробно описывается при изложении курса нервных болезней, что позволяет ограничиться только упоминанием, что в состав их входят двигательные пути, пути общей чувствительности, зрительные, слуховые, обонятельные и вкусовые пути, несущие периферические раздражения к соответствующим областям мозговой коры (к затылочным и височным долям, к крючковидной извилине и к operculum). Проекционные системы подходят к выпуклым частям извилин пучками, образуя значительную часть видимых хорошо на поперечных срезах через извилины радиальных волокон. Своим направлением от различных участков коры проекционные системы образуют лучистый венец и проходят через внутреннюю капсулу и задние отделы зрительного бугра (чувствительный путь). Сочетательные системы разделяются на короткие, длинные и спаечные. Короткие служат для связи между собою коры соседних извилин; длинные связывают различные области коры своего полушария; спаечные соединяют симметричные и несимметричные отделы противоположных полушарий. Если провести поперечный разрез через мозговую извилину, то при известных способах окраски, напр., по сп. Раl’я, можно легко различить на срезе ряд полосок, состоящих из пучков волокон, идущих параллельно мозговой поверхности. Не во всех извилинах эта слоистость выражена одинаково, особенно отчетлива она в извилинах теменной доли. Самый поверхностный слой носит название слоя тангенциальных волокон; тотчас под ним, почти примыкая к нему, лежит слой волокон, описанный Бехтеревым164) и Каеs'ом 165); еще ниже, соответственно уровню слоя зерен, а также слою глубоких пирамид в его пограничной части с полиморфным слоем, находятся две полоски Baillarger, наружная (Genneri) и внутренняя; между полосками Бехтерева и Baillarger, а также ниже внутренней полоски Baillarger, находятся два рыхлых слоя волокон, первый из которых называется plexus superradialis, а второй plexus interradialis. Полоски Baillarger образуются отчасти горизонтально идущими коллатералями пирамидных клеток. Еще глубже располагается резко выраженный слой, т. наз. интракортикальных или внутрикорковых волокон, слой сочетательных волокон Meynert'a, хорошо развитый и соединяющий соседние извилины. Волокна Meynert'a, известные под названием fibrae arcuatae breves, и составляют главную часть коротких сочетательных систем; к сочетательным системам относятся и вышеописанные поверхностные слои волокон. Кроме поперечно идущих волокон, в коре мозга различаются радиально расположенные пучки; они состоят из волокон проекционных систем, было сказано выше, и из волокон, принадлежащих системам сочетательным. Длинные сочетательные системы далеко еще не установлены в окончательном виде, вопрос об их анатомическом положении во многих отношениях представляется спорным и неопределенным, пучки, признаваемые одними авторами за ассоциационные или сочетательные системы, рассматриваются другими в качестве проекционных путей, а иногда даже совсем отвергаются. В настоящее время можно считать анатомически более или менее установленными следующие сочетательные системы *) 166-168). 1) Cingulum, поясной, дугообразный пучек, идущий в стреловидном направлении от обонятельных отделов в белом веществе внутренней части первой лобной извилины и средней части сводчатой извилины (g. fornicatus); эта система составляет главную массу белого вещества первой и второй краевых извилин (g. limbici) и доходит до переднего конца (полюса) височной доли. Надо полагать, что далеко не все волокна, образующие эту систему, проходят по всей ее длине, большинство же их гораздо короче (Quensel**)). 2) Fasciculus uncinatus, крючковатый пучек, идущий от височного полюса к глазничной поверхности лобной доли; пучек проходит на уровне порога островка, расширяясь в поперечном направлении от subst. perforata ant. до capsula externa. Резкий изгиб пучка при переходе из височной доли в лобную и дает основание для наименования его «крючковатым». 3) Fasciculus longitudinalis inferior, нижний продольный пучек, имеет стреловидное направление; мощный пучек, идущий от извилин височной доли к извилинам затылочной, соединяющий слуховые и зрительные центры; проходит вблизи наружной границы заднего и нижнего рогов бокового желудочка; волокна пучка также различной длины, направление их меняется в зависимости от положения соединяемых ими височных и затылочных извилин. Сочетательное значение этой системы подвергается сомнению F1есhsig'ом, который помещает в ней центростремительный зрительный путь. 4) Fasciculus longitudinalis superior, верхний продольный пучек, он же fasc. arcuatus, дугообразный; располагается между лобной, теменной и височной долями; проходит на уровне мозолистого тела дугообразной выпуклостью вверх. Система служит не только для соединения конечных пунктов, между которыми она находится, но соединяет и более близкие участки указанных областей, по крайней мере, из вторичных перерождений ее видно, что она состоит из сравнительно коротких сочетательных волокон. 5) Fasciculus occipito-frontalis, затылочно-лобный пучек, располагающийся между хвостатым и мозолистым телом, по мнению Quensel'я***), не имеет права на существование в качестве сочетательной системы; поводимому, к проекционным системам должен быть отнесен и fasc. subcallosus (подмозолистый пучек) Муратова****). Выделяются еще более короткие сочетательные системы: fasc. transversus cunei (Sachs) и fasc. transversus g. lingualis, оба пучка поперечного направления, первый соединяет верхнюю губу fiss. calcarinae с нижней наружной поверхностью затылочной доли, а второй идет от нижней губы той же борозды (Violet), no Quensel’ю*), однако, значение обеих систем следует признать спорным. Fasc. temporo-occiрitalis verticalis, височно-затылочный вертикальный пучек, находящийся между нижними затылочными, третьей височной и нижней теменной долькой; fasc. centro-parietalis, связывающий центральные извилины с нижней теменной долькой, и fasc. centro-frontalis — между центральными и лобными извилинами. Значение сочетательной системы придается также волокнам наружной капсулы, caps, externa. Спаечные системы также относятся к числу сочетательных систем; эти системы иначе называются коммиссуральными. К ним относится обширнейшая спайка между полушариями головного мозга, известная под названием мозолистого тела (corpus callosum), и передняя спайка (commissura anterior). Спаечные системы соединяют между собою не только строго симметричные участки обоих полушарий, как принято было думать прежде, но также и несимметричные, а следовательно, содержат перекрещивающиеся волокна (с. callosum). Мозолистое тело представляется спаечной сочетательной системой для всех извилин обоих полушарий головного мозга, за исключением обонятельных и передних отделов височных, сочетающихся между собою при посредстве волокон передней спайки. Поводимому, в состав мозолистого тела входят волокна, происходящие от корковых слоев малых и средних пирамидных и полиморфных клеток. Из всего приведенного здесь описания сочетательных систем головного мозга с несомненностью вытекает, что поверхностные пучки, короткие, длинные и спаечные сочетательные системы настолько совершенно связывают между собою клеточные элементы, центры соседних извилин, различно удаленные одна от другой области одного и того же полушария, симметричные и асимметричные участки противоположных полушарий, что возбужденное состояние одного центра легко может передаваться другому центру или другим центрам, находящимся от первоначального источника возбуждения на различном расстоянии.
X. Метод развития; теория ассоциационных центров Flechsig'a; ее слабые стороны; ее значение. Механизм процесса, лежащего в основе душевной деятельности; учение Сеченова и его значение; теория условных рефлексов И. П. Павлова и ее значение; приложение В. М. Бехтеревым об'ективной методики и условных рефлексов к изучению душевной деятельности человека. Схема процессов торможения в мозговой коре по Блуменау. Биологическая сущность нервно-психического процесса. Теория молекулярного притяжения Meynert'a. Связь психических процессов с явлениями кровообращения (Моssо и др.). Химический характер биологического процесса, лежащего в основе душевной деятельности. Не все отделы головного мозга имеют одинаковое значение для душевной жизни; помимо экспериментальных доказательств, основанных, главным образом, на методе выпадения функции, в пользу этого говорит и различное строение различных отделов мозга; мозговая кора, являющаяся собственно органом душевной деятельности, как видно особенно из упомянутых выше исследований Brodmann'a*), также имеет различное строение в различных областях. Для выяснения функций различных отделов мозговой коры Flechsig воспользовался разработанным им методом эмбриологическим или методом развития или миэлинизации. Метод основывается на общем принципе, по которому нервные волокна эквивалентные, принадлежащие одному пучку, несущие общую функцию, обкладываются мякотной оболочкой одновременно, волокна же разных систем, направленные к выполнению различных функций, миэлинизируются в различное время. Так, посредством изучения миэлинизации различных путей чувствительности, движения, коротких и длинных сочетательных систем удается выделить в мозговой коре определенные миэлогенетические участки или поля, миэлинизация которых, в смысле своей последовательности во времени, подчиняется определенному закону. На основании своего метода Flechsig выделил около 40 участков (полей) мозговой коры, которые по времени миэлинизации он расчленил на три группы: ранней, поздней и промежуточной миэлинизации. Аналогично проекционным и ассоциационным системам головного мозга, Flechsig соединяет упомянутые участки коры в более обширные центры, — проекционные и сочетательные. Проекционных центров четыре, из них обширнейший — центр телесных ощущений и движений, охватывает обе центральные извилины, g. supramarginalis и частью задние отделы лобных извилин; этот центр лежит в основе процессов самосознания, с ним связаны двигательные процессы, направленные на удовлетворение телесных нужд (питание, жевание, дыхание, схватывание предметов и т. под.); кроме этого центра кожномышечных ощущений, Flechsig выделяет зрительный, слуховой, обонятельный и вкусовой центры. Ассоциационных или сочетательных центров установлено F1есhsig'ом три: задний или теменно-височный, передний или лобный и островковый. Проекционные центры занимают около трети мозговой коры, на долю сочетательных приходится две трети. Теменно-височный центр распространяется на теменную, большую часть затылочной доли и вторую и третью извилины обоих полушарий; с деятельностью этого центра связано положительное знание, процессы воображения и фантазии, способность речи, высшая сочетательная деятельность, все то, что связано с понятием “духовных” процессов, с развитием гениальности. Лобный центр охватывает поверхность лобных долей, в нем локализируются чувствования и волевые процессы, связанные с “я” человека; в нем закрепляются следы, память сознательных переживаний. Островковый центр ограничивается областью insula Reilii, его значение не так определенно, он связан с речевыми функциями. Сочетательные центры связаны между собою длинными сочетательными системами 169-171). Нельзя не признать, что Flechsig сделал смелую научную попытку дать анатомо-физиологическое обоснование локализации душевной деятельности, и в этом его большая заслуга. Однако, теория F1есhsig'a не осталась без существенных возражений, высказанных особенно Hitzig'ом172), v. Mоnakоw’ым173) и Vogt'ом174) в заседании неврологической секции ХIII-го международного медицинского конгресса в Париже в 1900 году. Было указано, что принцип одновременности и разновременности миэлинизации, как признак тождества или различия функций соответствующих областей, а также принадлежности к одной или различным системам, далеко еще не является доказанным и общепризнанным. По мнению v. Monakow'a, в этом процессе миэлинизации имеет значение не столько функция, сколько васкуляризация, снабжение миэлинизирующейся области сосудами. Время миэлинизации различных участков подвержено довольно значительным индивидуальным колебаниям. Неправильно слишком резкое разделение проекционных и сочетательных систем, которые несомненно представляются довольно смешанными. В своей теории Flechsig не оценивает достаточно значения подкорковых узлов. По мнению Vogt'a, время миэлинизации волокон теснейшим образом связано с их калибром, а именно: более толстые волокна обкладываются миэлином раньше, нежели тонкие. Дата добавления: 2016-03-26 | Просмотры: 298 | Нарушение авторских прав |