|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология
|
Строение, развитие и функциональное значение различных отделов нервной системы
Спинной мозг. Спинной мозг представляет собой длинный тяж. Он заполняет полость позвоночного канала и имеет сегментарное строение, соответствующее строению позвоночника.
В центре спинного мозга расположено серое вещество — скопление нервных клеток, окруженное белым веществом, образованным нервными волокнами (рис. 7).
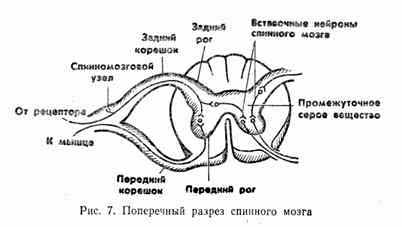
В спинном мозге находятся рефлекторные центры мускулатуры туловища, конечностей и шеи. С их участием осуществляются сухожильные рефлексы в виде резкого сокращения мышц (коленный, ахиллов рефлексы), рефлексы растяжения, сгибательные рефлексы, разные рефлексы, направленные на поддержание определенной позы. Рефлексы мочеиспускания и дефекации, рефлекторного набухания полового члена и извержения семени у мужчин (эрекция и эякуляция) связаны с функцией спинного мозга.
Спинной мозг осуществляет и проводниковую функцию. Нервные волокна, составляющие основную массу белого вещества, образуют проводящие пути спинного мозга. По этим путям устанавливается связь между различными частями ЦНС и проходит импульсация в восходящем и нисходящем направлениях. По этим путям поступает информация в вышележащие отделы мозга, от которых отходят импульсы, изменяющие деятельность скелетной мускулатуры и внутренних органов.
Деятельность спинного мозга у человека в значительной степени подчинена координирующим влияниям вышележащих отделов ЦНС.
Обеспечивая осуществление жизненно важных функций, спинной мозг развивается раньше, чем другие отделы нервной системы. Когда у эмбриона головной мозг находится на стадии мозговых пузырей, спинной мозг достигает уже значительных размеров. На ранних стадиях развития плода спинной мозг заполняет всю полость позвоночного канала. Затем позвоночный столб обгоняет в росте спинной мозг, и к моменту рождения он заканчивается на уровне третьего поясничного позвонка. У новорожденных длина спинного мозга 14—16 см, к 10 годам она удваивается. В толщину спинной мозг растет медленно. На поперечном срезе спинного мозга детей раннего возраста отмечается преобладание передних рогов над задними. Увеличение размеров нервных клеток спинного мозга наблюдается у детей в школьные годы.
Головной мозг. Спинной мозг непосредственно переходит в стволовую часть головного мозга, расположенную в черепе (рис. 8).
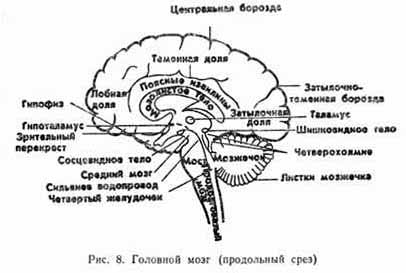
Прямым продолжением спинного мозга является продолговатый мозг, который вместе с мостом мозга (варолиев мост) образует задний мозг. его нервные клетки образуют нервные центры регулирующие рефлекторные функции сосания, глотания, пищеварения, сердечно-сосудистой и дыхательной систем, а также ядра V-XII пар черепных нервов и парасимпатических нервных волокон, идущих в их составе.
Необходимость реализации перечисленных жизненно важных функций с момента рождения ребенка определяет степень зрелости структур продолговатого мозга уже в период новорожденности. К 7 годам созревание ядер продолговатого мозга в основном заканчивается.
На уровне продолговатого мозга начинается ретикулярная формация, состоящая из сети нервных клеток, с которыми контактируют афферентные и эфферентные пути. Аксоны различных нейронов образуют множественные коллатерали, контактируя с огромным числом ретикулярных клеток. Один аксон может взаимодействовать с 27 500 нейронов. Ретикулярная формация распространяется на уровень среднего и промежуточного мозга. В ретикулярной формации выделяют нисходящую систему, регулирующую, под влиянием воздействия из высших отделов ЦНС, рефлекторную деятельность спинного мозга и мышечный тонус. К ней относятся передняя часть продолговатого мозга и средняя часть варолиева моста. Восходящая система — структуры ствола, среднего и промежуточного мозга — получает импульсы из спинного мозга и сенсорных систем, оказывает общее неспецифическое влияние на вышележащие отделы головного мозга. Ей, как будет показано дальше, принадлежит важнейшая роль в регуляции уровня бодрствования и организации поведенческих реакций.
В состав среднего мозга входят ножки мозга и крыша мозга.
Здесь расположены скопления нервных клеток в виде верхних и нижних бугров четверохолмия, красного ядра, черной субстанции, ядер глазодвигательного и блокового нервов, ретикулярной формации.
В верхних и нижних буграх четверохолмия замыкаются простейшие зрительные и слуховые рефлексы и осуществляется их взаимодействие (движение ушей, глаз, поворот в сторону раздражителя). Черная субстанция участвует в сложной координации движений пальцев рук, актов глотания и жевания. Красное ядро имеет непосредственное отношение к регуляции мышечного тонуса. Позади продолговатого мозга и моста расположен мозжечок. Мозжечок— орган, регулирующий и координирующий двигательные функции и их вегетативное обеспечение. Информация от различных мышечных, вестибулярных, слуховых и зрительных рецепторов, сигнализирующая о положении тела в пространстве и характере выполнения движений, интегрируется в мозжечке с влияниями от вышележащих отделов головного мозга, что обеспечивает реализацию плавного координированного двигательного акта, основанного на принципе обратной связи. Удаление мозжечка не влечет за собой потерю способности к движению, но нарушает характер выполняемых действий.
Усиленный рост мозжечка отмечается на первом году жизни ребенка, что определяется формированием в течение этого периода дифференцированных и координированных движений. В дальнейшем темпы его развития снижаются. К 15 годам мозжечок достигает размеров взрослого.
Важнейшие функции выполняют структуры промежуточного мозга, включающего в себя зрительный бугор (таламус) и подбугровую область (гипоталамус). Гипоталамус, несмотря на небольшие размеры, содержит десятки высокодифференцированных ядер.
Гипоталамус связан с вегетативными функциями организма и осуществляет координационно-интегративную деятельность симпатического и парасимпатического отделов. Пути из гипоталамуса идут к среднему, продолговатому и спинному мозгу, оканчиваясь на нейронах — источниках преганглионарных волокон. Вегетативные эффекты гипоталамуса, разных его отделов имеют неодинаковые направленность и биологическое значение. Задние отделы приводят к возникновению эффектов симпатического типа, передние— парасимпатического. Восходящие влияния этих отделов также разнонаправлены: задние оказывают возбуждающее влияние на кору больших полушарий, передние — тормозящее. Связь гипоталамуса с одной из важнейших желез внутренней секреции — гипофизом — обеспечивает нервную регуляцию эндокринной функции.
В клетках ядер переднего гипоталамуса вырабатывается нейросекрет, который по волокнам гипоталамо-гипофизарного пути транспортируется в нейрогипофиз. Этому способствуют и обильное кровоснабжение, и сосудистые связи гипоталамуса и гипофиза.
Гипоталамус и гипофиз часто объединяют в гипоталамо-гипофизарную систему, играющую важнейшую роль в регуляции желез внутренней секреции.
Одно из крупных ядер гипоталамуса — серый бугор — принимает участие в регуляции функций многих эндокринных желез и обмена веществ. Разрушение серого бугра вызывает атрофию половых желез. Его длительное раздражение может привести к раннему половому созреванию, возникновению язв на коже, язвы желудка и двенадцатиперстной кишки.
Гипоталамус принимает участие в регуляции температуры тела. Доказана его роль в регуляции водного обмена, обмена углеводов. Ядра гипоталамуса участвуют во многих сложных поведенческих реакциях (половые, пищевые, агрессивно-оборонительные). Гипоталамус играет важную роль в формировании основных биологических мотиваций (голод, жажда, половое влечение) и эмоций положительного и отрицательного знака. Многообразие функций, осуществляемых структурами гипоталамуса, дает основание расценивать его как высший подкорковый центр регуляции жизненно важных процессов, их интеграции в сложные системы, обеспечивающие целесообразное приспособительное поведение.
Дифференцировка ядер гипоталамуса к моменту рождения не завершена и протекает в онтогенезе неравномерно. Развитие ядер гипоталамуса заканчивается в период полового созревания.
Таламус (зрительный бугор) составляет значительную часть промежуточного мозга. Это многоядерное образование, связанное двусторонними связями с корой больших полушарий. В его состав входят три группы ядер. Релейные ядра передают зрительную, слуховую, кожно-мышечно-суставную информацию в соответствующие проекционные области коры больших полушарий. Ассоциативные ядра передают ее в ассоциативные отделы коры больших полушарий. Неспецифические ядра (продолжение ретикулярной формации среднего мозга) оказывают активизирующее влияние на кору больших полушарий.
Центростремительные импульсы от всех рецепторов организма (за исключением обонятельных), прежде чем достигнут коры головного мозга, поступают в ядра таламуса. Здесь поступившая информация перерабатывается, получает эмоциональную окраску и направляется в кору больших полушарий.
К моменту рождения большая часть ядер зрительных бугров хорошо развита. После рождения размеры зрительных бугров увеличиваются за счет роста нервных клеток и развития нервных волокон.
Онтогенетическая направленность развития структур промежуточного мозга состоит в увеличении их взаимосвязей с другими мозговыми образованиями, что создает условия для совершенствования координационной деятельности его различных отделов и промежуточного мозга в целом. В развитии промежуточного мозга существенная роль принадлежит нисходящим влияниям корковых полей конечного мозга.
Конечный, или передний, мозг, включает в себя базальные ганглии и большие полушария.
Основной частью конечного мозга, достигающей наибольшего развития у человека, являются большие полушария.
Большие полушария головного мозга расположены над передней дорзальной поверхностью ствола мозга. Они соединены крупными пучками нервных волокон, образующих мозолистое тело. У взрослого человека масса больших полушарий составляет около 80% массы головного мозга и в 40 раз превышает массу ствола.
Структурно-функциональная организация коры головного мозга. Кора больших полушарий представляет собой тонкий слой серого вещества на поверхности полушарий. В процессе эволюции поверхность коры интенсивно увеличивалась по размеру за счет появления борозд и извилин. Общая площадь поверхности коры у взрослого человека достигает 2200—2600 см2. Толщина коры в различных частях полушарий колеблется от 1,3 до 4,5 мм. В коре насчитывается от 12 до 18 млрд. нервных клеток. Отростки этих клеток образуют огромное количество контактов, что и создает условия для сложнейших процессов обработки и хранения информации.
На нижней и внутренней поверхности полушарий расположены старая и древняя кора, или архи- и палеокортекс. Функционально эти отделы коры больших полушарий тесно связаны с гипоталамусом, миндалиной, некоторыми ядрами среднего мозга. Все эти структуры составляют лимбическую систему мозга. Как будет показано дальше, лимбическая система играет важнейшую роль в формировании эмоций и внимания. В старой и древней коре расположены также высшие центры вегетативной регуляции.
На наружной поверхности полушарий расположена филогенетически наиболее новая кора, появляющаяся только у млекопитающих и достигающая наибольшего развития у человека. Это неокортекс.
Кора больших полушарий имеет 6—7 слоев, различающихся формой, величиной и расположением нейронов (рис. 9). Между нервными клетками всех слоев коры в процессе их деятельности возникают как постоянные, так и временные связи.
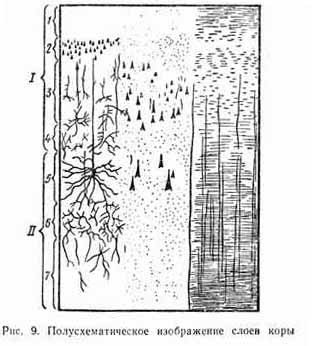
По особенностям клеточного состава и строения кору больших полушарий разделяют на ряд участков. Их называют корковыми полями.
Под корой располагается белое вещество больших полушарий. В составе белого вещества различают ассоциативные, комиссуральные и проекционные волокна. Ассоциативные волокна связывают между собой отдельные участки одного и того же полушария. Короткие ассоциативные волокна связывают между собой отдельные извилины и близкие поля. Длинные волокна — извилины различных долей в пределах одного полушария. Комиссуральные волокна связывают симметричные части обоих полушарий. Большая часть их проходит через мозолистое тело. Проекционные волокна выходят за пределы полушарий. Они входят в состав нисходящих и восходящих путей, по которым осуществляется двусторонняя связь коры с нижележащими отделами ЦНС.
Известны случаи рождения детей, лишенных коры больших полушарий головного мозга. Это анэнцефалы. Они обычно живут всего несколько дней. Но известен случай жизни анэнцефала в течение 3 лет 9 месяцев. После его смерти при вскрытии оказалось, что большие полушария отсутствовали полностью, на их месте были обнаружены два пузыря. В течение первого года жизни этот ребенок почти все время спал. На звук и свет не реагировал. Прожив почти 4 года, он не научился говорить, ходить, узнавать мать, хотя врожденные реакции (некоторые) у него проявлялись: он сосал, когда ему вкладывали в рот сосок материнской груди или соску, глотал и т. п.
Наблюдения над животными с удаленными полушариями головного мозга и над анэнцефалами показывают, что в процессе филогенеза резко возрастает значение высших отделов ЦНС в жизни организма. Происходит кортиколизация функций, подчинение сложных реакций организма коре больших полушарий. Все, что приобретается организмом в течение индивидуальной жизни, связано с функцией больших полушарий головного мозга. С функцией коры больших полушарий связана высшая нервная деятельность. Взаимодействие организма с внешней средой, его поведение в окружающем материальном мире связаны с большими полушариями головного мозга. Вместе с ближайшими подкорковыми центрами, стволом мозга и спинным мозгом большие полушария объединяют отдельные части организма в единое целое, осуществляют нервную регуляцию функций всех органов.
В опытах с удалением различных участков коры, их раздражением и при регистрации электрической активности мозга установлено наличие трех типов корковых областей: сенсорные, моторные и ассоциативные (рис. 10).
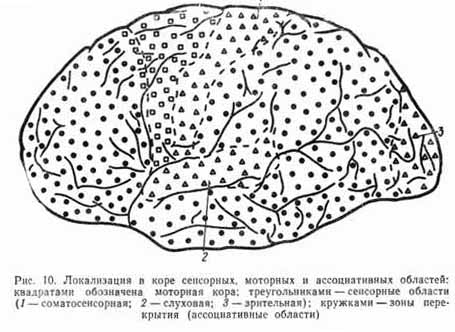
Сенсорные области коры больших полушарий. Афферентные волокна, несущие сигналы от различных рецепторов, приходят к определенным зонам коры. Каждому рецепторному аппарату соответствует в коре определенная область. И. П. Павловым эти области были названы корковым ядром анализатора. В сенсорных зонах выделяют первичные и вторичные проекционные поля.
Нейроны проекционных первичных полей выделяют отдельные признаки сигнала. В области зрительной проекции, например, анализируются место объекта в поле зрения, направление движения, контур, цвет, контраст. Разрушение этой области приводит к потере способности к первичному анализу внешних стимулов в определенной части поля зрения. При раздражении первичной зрительной зоны во время операций отмечается появление световых мельканий, цветовых пятен; при раздражении проекционного поля слуховой коры пациент слышит тоны, отдельные звуки.
При ограниченном поражении вторичных, например зрительных, полей больной отчетливо видит отдельные элементы изображения, но не может объединить их в целостный образ, узнать знакомый предмет (зрительная агнозия). Раздражение вторичных сенсорных зон у человека во время операции вызывает оформленные предметные зрительные и сложные слуховые галлюцинации: звуки музыки, речи и т. д.
Сенсорные зоны локализованы в определенных областях коры: зрительная сенсорная зона располагается в затылочной области обоих полушарий, слуховая — в височной области, зона вкусовых ощущений — в нижней части теменных областей, соматосенсорная зона, анализирующая импульсацию с рецепторов мышц, суставов, сухожилий, кожи, располагается в области задней центральной извилины (см. рис. 10).
Моторные области коры. Зоны, раздражение которых закономерно вызывает двигательную реакцию, называют моторными или двигательными. Они расположены в области переднецентральной извилины. Моторная кора имеет двусторонние внутрикорковые связи со всеми сенсорными областями. Это обеспечивает тесное взаимодействие сенсорных и моторных зон.
Ассоциативные области коры. Кора больших полушарий человека" характеризуется наличием обширной области, не имеющей прямых афферентных и эфферентных связей с периферией. Эти области, связанные обширной системой связей ассоциативных волокон с сенсорными и моторными зонами, получили название ассоциативных или третичных корковых зон. В задних отделах коры они расположены между теменными, затылочными и височными областями, в передних отделах они занимают основную поверхность лобных долей. Ассоциативная кора либо отсутствует, либо слабо развита у всех млекопитающих до приматов. У человека заднеассоциативная кора занимает примерно половину, а лобные области 25% всей поверхности коры. По строению они отличаются особенно мощным развитием верхних ассоциативных слоев клеток в сравнении с системой афферентных и эфферентных нейронов. Их особенностью является также наличие полисенсорных нейронов — клеток, воспринимающих информацию из различных сенсорных систем.
В ассоциативной коре расположены и центры, связанные с речевой деятельностью. Ассоциативные области коры рассматриваются как структуры, ответственные за синтез поступающей информации, и как аппарат, необходимый для перехода от наглядного восприятия к абстрактным символическим процессам. С ассоциативными зонами коры связано формирование свойственной только человеку второй сигнальной системы.
Клинические наблюдения показывают, что при поражении заднеассоциативных областей нарушаются сложные формы ориентации в пространств, конструктивная деятельность, затрудняется выполнение всех интеллектуальных операций, которые осуществляются с участием пространственного анализа (счет, восприятие сложных смысловых изображений). При поражении речевых зон нарушается возможность восприятия и воспроизведения речи. Поражение лобных отделов коры приводит к невозможности осуществления сложных программ поведения, требующих выделения значимых сигналов на основе прошлого опыта и предвидения будущего.
Развитие коры больших полушарий как филогенетически нового образования происходит в течение длительного периода онтогенеза. К моменту рождения ребенка кора больших полушарий имеет такой же тип строения, как у взрослого. Однако поверхность ее после рождения значительно увеличивается за счет формирования мелких борозд и извилин. В течение первых месяцев жизни развитие коры идет очень быстрыми темпами. Большинство нейронов приобретает зрелую форму, происходит миелиниза-ция нервных волокон. Различные корковые зоны созревают неравномерно. Наиболее рано созревает соматосенсорная и двигательная кора,несколько позже зрительная и слуховая. Созревание проекционных (сенсорных и моторных) зон в основном завершается к 3 годам. Значительно позже созревает ассоциативная кора. К 7 годам отмечается значительный скачок в развитии ассоциативных областей (рис. 11).
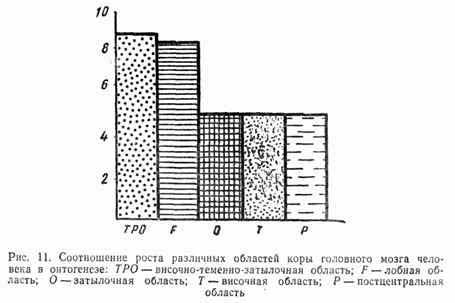
Однако их структурное созревание— дифференцировка нервных клеток, формирование нейронных ансамблей и связей ассоциативной коры с другими отделами мозга — происходит вплоть до подросткового возраста. Наиболее поздно созревают лобные области коры. Как будет показано ниже, постепенность созревания структур коры больших полушарий определяет возрастные особенности высших нервных функций и поведенческих реакций детей дошкольного и младшего школьного возраста.
Электроэнцефалограмма. Для изучения функциональной активности коры больших полушарий и ее взаимоотношения с подкорковыми структурами широко используется метод регистрации биотоков мозга. Суммарная постсинаптическая активность нервных элементов коры больших полушарий, зарегистрированная с поверхности черепа, называется электроэнцефалограммой.
Метод регистрации электроэнцефалограммы (ЭЭГ), позволяющий объективно изучать функциональную активность мозга, образно называют зеркалом мозга.
Современная электронная аппаратура позволяет регистрировать биоэлектрические потенциалы одновременно с многих точек поверхности черепа. Многоканальные электроэнцефалографы — приборы, включающие усилители биопотенциалов и регистратор (обычно чернилописец), позволяют одновременно регистрировать электрическую активность из области проекции на черепе различных участков коры больших полушарий.
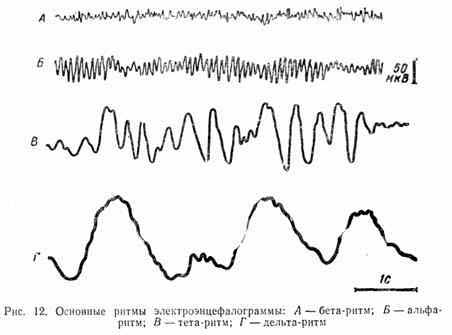
В ЭЭГ взрослого человека выделяют 4 основных типа ритмических электрических колебаний (рис. 12), каждый из которых характерен для определенного функционального состояния.
Альф (α) - ритм регистрируется в состоянии спокойного бодрствования. Показано, что он является оптимальным фоном для приема и переработки информации, поступающей в кору больших полушарий. Частота этого ритма — от 8 до 13 колебаний в 1 с. Для взрослого человека наиболее характерен α- ритм частотой 10 Гц.
Бета (β) - ритм характерен для состояния активного бодрствования. Он наблюдается при действии неожиданного раздражителя, при умственном напряжении. β -ритм, отражающий возбуждение нервных элементов коры, характеризуется высокой частотой (14—50 Гц).
Тета (θ) - и дельта (δ) - ритмы наблюдаются во время сна. Их частота соответственно 4—7 и 1—3 Гц. В бодрствующем состоянии эти ритмы у взрослого человека наблюдаются при патологических состояниях, связанных с повышенной активностью подкорковых диэнцефальных структур мозга. Тета-ритмы также могут возникать при крайнем эмоциональном напряжении.
Возрастные особенности электроэнцефалограммы детей и подростков, Анализ электроэнцефалограмм детей различного возраста показывает, что подкорковые структуры, являющиеся наиболее филогенетически древними образованиями мозга и играющие важнейшую роль в обеспечении жизненно важных функций, созревают значительно раньше высших отделов центральной нервной системы (коры больших полушарий). Медленная активность в электроэнцефалограмме, отражающая их функционирование, практически сформирована уже в грудном возрасте и регистрируется уже у новорожденных детей. В результате этого опережающего созревания подкорковых структур ЭЭГ сна детей грудного и раннего детского возраста практически не отличается от взрослого типа. Существенные изменения претерпевает ЭЭГ спокойного бодрствования (рис. 13).

Основной ритм ЭЭГ покоя — альфа-ритм, отражающий функциональную активность коры больших полушарий, в ходе индивидуального развития появляется впервые у детей в 3-месячном возрасте в виде отдельных групп колебаний, перемежающихся большим количеством медленных волн. С возрастом, по мере структурно-функционального созревания коры больших полушарий, продолжительность времени регистрации альфа-ритма и соответственно его представленность в ЭЭГ увеличивается. Однако еще в 5-летнем возрасте альфа-ритм не является доминирующей формой активности и ЭЭГ покоя носит полиритмичный характер. Существенные изменения ЭЭГ покоя обнаруживаются в 6 лет, когда четко выявляется ведущая частота в диапазоне альфа-ритма. Это дает основание рассматривать возраст 6 лет как существенный этап в организации состояния покоя как оптимального фона для восприятия внешней информации. Однако в 6 лет, так же как и в 7—8 лет, альфа-ритм характеризуется сниженной частотой (8—9 Гц) и нестабильностью. При нагрузках, в процессе школьного обучения частота альфа-ритма снижается выраженность его уменьшается и увеличивается выраженность колебаний типа тета. Колебания этого типа в состо; покоя наблюдаются у взрослых только при патологической активности подкорковых структур или резко выраженных эмоциональных состояниях Их наличие в ЭЭГ во время спокойного бодрствования у здоровых детей есть результат возрастной специфики корково-подкоркового взаимодействия, результат меньшей, чем у взрослого, степени подавляющего воздействия коры на подкорковые структуры.
По мере созревания коры характер корково-подкоркового взаимодействия существенно изменяется. Близкие к типу взрослого корково-подкорковые взаимоотношения с выраженным тормозным влиянием коры на подкорковые структуры устанавливаются к 10—12 годам, когда по показателям ЭЭГ кора больших полушарий достигает значительной зрелости. В ЭЭГ регистрируется альфа-ритм, по своему рисунку, амплитуде, частоте существенно не отличающийся от такового взрослых. ЭЭГ приобретает устойчивый, стабильный характер. Частота основного ритма и его представленность в ЭЭГ не меняются в течение учебного года.
В 12—15-летнем возрасте снова наблюдается усиление подкорковой активности. Это период полового созревания. Он характеризуется повышенной активностью одного из отделов промежуточного мозга — гипоталамуса, функция которого тесно связана с деятельностью желез внутренней секреции. На ЭЭГ это находит свое отражение в уменьшении частоты альфа-ритма и его нестабильности в течение учебного года, увеличивается и представленность медленноволновой активности. В поведении подростков в этот период отмечаются повышенная нервозность, несдержанность, неустойчивость эмоциональных реакций. К завершению подросткового возраста отмеченные на ранних стадиях полового созревания отклонения в ЭЭГ покоя исчезают. В ЭЭГ покоя четко доминирует альфа-ритм, параметры которого соответствуют взрослому.
Прослеженное при анализе ЭЭГ покоя структурно-функциональное созревание коры больших полушарий является чрезвычайно важным фактором в формировании поведенческих реакций ребенка. Оно способствует нарастанию сдержанности, контролируемости и осмысленности поступков. Наблюдаемое в течение длительного периода индивидуального развития ребенка усиление организующей роли коры больших полушарий является основой для формирования высших нервных и психических функций.
Дата добавления: 2014-12-11 | Просмотры: 2732 | Нарушение авторских прав
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | 40 | 41 | 42 | 43 | 44 | 45 | 46 | 47 | 48 | 49 | 50 | 51 | 52 | 53 | 54 | 55 | 56 | 57 | 58 | 59 | 60 | 61 | 62 |
|