|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Результаты/ОбсуждениеПлотная пространственная сетка из 7150 квази-ориентиров использовалась для создания 3D изображения лиц участников на общей системе координат (рис. 1). Квази-ориентиры определяются здесь как в значительной степени гомологичные вершины в картированной сетке. Сетка строится автоматически, что исключает сложную и подверженную ошибкам процедуру с размещением лицевых ориентиров вручную [8], [9], [17]. Погрешности билатеральной симметрии были удалены с помощью усреднения каждого лица с его зеркальным отображением [18], [19]. С помощью анализа главных компонент 21450 симметрично расположенных 3D-координат квази-ориентиров (X, Y и Z для каждого из 7150 квази-ориентира), который применялся ко всем 592 добровольцам, было получено 44 главные компоненты (PC), которыми были представлены 98% изменчивости формы лица, и которые определяли многомерное пространство лица. Эффекты первых 10 главных компонент показаны на рисунке 2. Некоторые из этих компонент (например, PC4, PC5) отображают изменения только в отдельных частях лица. Тем не менее, множество компонент (например, PC1, PC2, PC3) отображают эффекты на многих частях лица. Кроме того, хотя главные компоненты статистически независимы, любая определенная часть лица зависит от нескольких компонент. Таким образом, вполне вероятно, неверно думать, будто каждая компонента представляет собой отдельную морфологическое черту в результате действия конкретных генов. Использование BRIM, которое объединяет независимые эффекты от главных компонент, не дает информации о их биологическом значении, если таковые имеются, а интегрирует информацию от одной или всех компонент в одну переменную, которая является моделируемой независимой переменной. Таким образом, BRIM также снимает проблему множественного тестирования, присущей другим методам описания общей изменчивости лица. Другими словами, гипотеза, оказывает ли определенный ген значительное влияние на форму лица, может быть проверена с помощью одного статистического теста (текст S1).
A) исходная поверхность, B) исключение нелицевых элементов, C) создание зеркального изображения, D) антропометрическая маска квази-ориентиров, E) картирование, F) отраженное картирование, G) симметризация, H) реконструирование. doi:10.1371/journal.pgen.1004224.g001
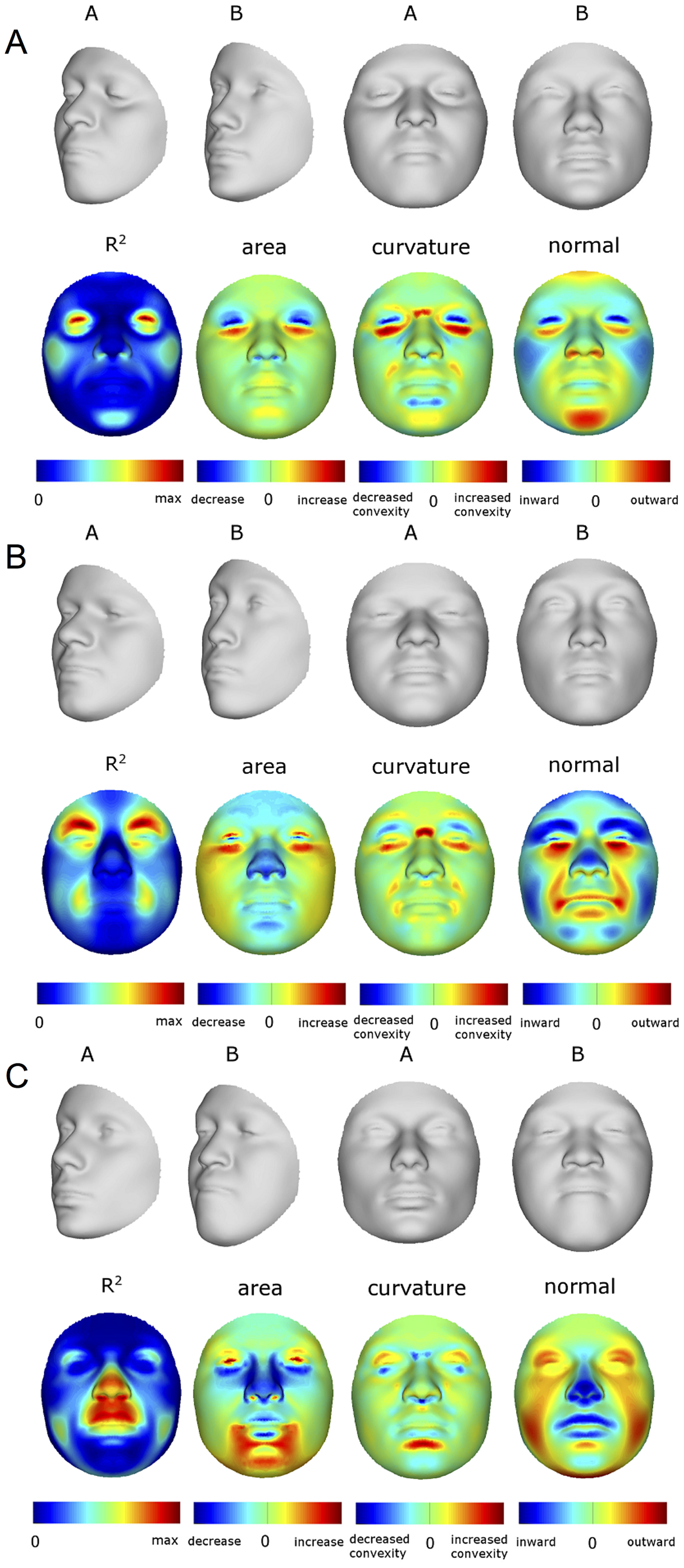
Эффекты первых 10 PCs (A–J), оказываемые на параметры изменения формы лица (FSCPs). Первыми показаны эффекты смещения каждого квази-ориентира, затем крайние варианты преобразований (лица серого цвета), отношение площадей между обоими вариантами, кривизна преобразований, отношения кривизны между обоими вариантами, и, наконец, нормальное смещение, которое показано через магнитуду смещения одного квази-ориентира по нормали к поверхности первого преобразования (лица серого цвета с левой стороны). doi:10.1371/journal.pgen.1004224.g002
BRIM – это расширенный метод моделирования связей, который использует зависимые переменные для уточнения и, в некоторых случаях, для преобразования одной или нескольких первоначальных независимых переменных. Другими словами, в отличие от альтернативных методов, BRIM использует многомерную матрицу зависимых переменных для проведения принудительной подстановки по отдельным объектам (leave-one-out forced imputation), чтобы обновить начальные значения независимых переменных и создать новый тип переменной - независимую переменную, полученную методом подстановок зависимых переменных или RIP-переменную (the response-based imputed predictor variable, RIP) (рис. S2). BRIM – это процесс на основе бутстреппинга, и, таким образом, можно отслеживать с каждой итерацией преобразование независимых переменных (номера S5, S6, S7, S8, S9). BRIM также нужен для исправления погрешности измерений, ошибок присвоения значений независимым переменным, и других источников статистических ошибок (Текст S1). В итерационной схеме бутстреппинга контроль по отдельным объектам используется, чтобы избежать переобучения модели и допустить использование стандартных статистических методов в тестировании гипотез, такие как корреляционный анализ, ANOVA и анализ кривой операционных характеристик приемника или ROC-кривой (receiver operating characteristic curve analysis, ROC) [20] для проверки значимости связи между независимыми переменными и RIP-переменными. Кроме того, отношения между RIP-переменными и зависимыми переменными, т.е. 450 параметрами лица, допускают визуализацию и количественную оценку влияния первых на форму лица. RIP-переменные, моделирующие влияние пола (RIP-S) и происхождения генома (RIP-А), а также те переменные, что учитывают эффект конкретных генетических маркеров (RIP-GS), могут быть визуализированы с помощью двух основных методов: преобразование формы и использование теплокарт. Мы использовали три типа сводных данных (отношение площадей, нормальное смещение (normal displacement) и разность кривизны), которые можно представить в виде теплокарт для количественной оценки отдельных изменений на лице. Эти показатели изменения лица, наряду с определенными расстояниями, углами и пространственными отношениями между ориентирами, могут быть названы параметрами изменения формы лица (FSCPs). FSCPs являются способом преобразования изменений параметров лица из абстрактной формы в визуальное изображение и в «слова». Такие термины используются в клинических и антропологических описаниях лиц и, таким образом, мы можем сравнить их с результатами BRIM (например, рис S28, S29, S30, S31, S32, S33, S36, S37, S38, а в таблице S1). Статистическая значимость этих и связанных с ними FSCPs может быть проверена с помощью метода перестановок.
Верхняя строка каждой панели показывает преобразование формы в диапазоне трех стандартных отклонений выше и ниже среднего RIPs в этой выборке. Вторая строка показывает R2 (доля общей изменчивости для каждого квази-ориентира) и три первичных параметра изменения формы лица: отношение площади, разность кривизны и нормальное смещение. На нижней строке желтым цветом обозначены участки лица, отличия между двумя преобразованиями которых статистически значимо (p <0.001). Максимальные значения R2 для RIP-A и RIP-S равны 40.83% и 38.21%, соответственно. doi:10.1371/journal.pgen.1004224.g003
Как и ожидалось, многие части лица зависимы как от этно-популяционного происхождения, так и от пола. На рисунке 3 показаны частичные эффекты, оказываемые RIP-А и RIP-S на форму лица, с использованием преобразования формы и теплокарт для оценки величины эффекта (R2) и трех основных FSCPs. Участки лица, которые статистически значимы (р<0,001) по величине эффекта и FSCPs, показаны на рисунке 3 желтым цветом (не зеленые участки на нижней панели). Преобразование формы по RIP-А и RIP-S показано в диапазоне трех стандартных отклонений плюс-минус средние уровни RIP-А и RIP-S в этих образцах. Как видно на панелях, показывающих размер эффекта (R2) на рисунке 3, доля общей изменчивости отдельных черт лица, зависящая от RIP-A и RIP-S, может быть существенной. В общем, до трети всей изменчивости определенных черт лица объясняется этими двумя переменными. RIP-А, в первую очередь, оказывает влияние на нос и губы и, в меньшей степени, на округлость лица, нижнюю челюсть и надбровные дуги. Пол оказывает гораздо больший эффект, чем родословная, на надбровные дуги и щеки, и меньший эффект - на нос и участки под глазами. FSCPs может продемонстрировать конкретные пути, с помощью которых отдельные RIP-переменные влияют на лицо. Например, отношение площадей показывает увеличенную площадь поверхности для медиальной глазной щели, сторон носа и передней части подбородка у европейского конца RIP-А-диапазона, и большую площадь поверхности для ноздрей и губ у западно-африканского конца RIP-А-диапазона. Разность кривизны демонстрирует верхнюю часть губного желобка в качестве лицевого элемента, который очень выпуклый на европейском конце диапазона и очень вогнутый на западноафриканском конце спектра RIP-A-диапазона. Разность кривизны, зависящая от RIP-A, также наблюдается для спинки носа, надбровных дуг и подбородка. RIP-S показывает наибольший эффект на надбровные дуги, спинку носа, носовую перегородку, скулы и щеки. Нос, губы, медиальная угол глазной щели и нижняя челюсть также имеют зависимостьот RIP-S. Крупнейшие различия в кривизне лица, связанные с изменением RIP-S, характерны для надбровных дуг и спинки носа. Несмотря на сложность проявления эффекта от RIP-А и RIP-S на черты лица, эти переменные являются полезными показателем степени, в которой у определенных лиц проявляется эффект этно-популяционного происхождения и пола соответственно. Это проявляется в прочной связи, наблюдаемой между RIP-А и происхождением генома и оцененной с помощью 68 маркеров этно-популяционного происхождения (г = 0,81, р <0,001; рисунок 4А). Примерно две трети изменчивости в RIP-A в трех западноафриканских /европейских смешанных популяциях объясняется происхождением генома. Аналогичным образом, как показано на рисунке 4В, RIP-S очень отличается между полами. Анализ кривой операционных характеристик приемника (рис. S32) показывает, что площадь под кривой RIP-S равна 0,994 (р <0,001). Происхождение генома независимо от пола объясняет 9,6% общих изменений лица, в то время как пол независимо от происхождения объясняет 12,9% общей изменчивости лица (табл. S3). Большая доля изменчивости лица, как и генетическая изменчивость человека в целом, является общей для различных человеческих популяций и представителей обоих полов.
(A) RIP-A и этно-популяционное происхождение генома, которое рассчитывается с использованием исходной панели из 68 AIMs. RIP-A рассчитывается по оценке происхождения генома трех популяций (N = 592). В легенде популяции обозначены: США – черные кружки, Бразилия – красные кружки и Капа-Верде – синие кружки. (B) Гистограммы RIP-S получены по данным анкетирования. doi:10.1371/journal.pgen.1004224.g004
Мы использовали альтернативные подмножества маркеров этно-популяционного происхождения и альтернативные выборки населения, чтобы проверить надежность оценки этно-популяционного происхождения черт лица (RIP-А). Значения RIP-A были получены с помощью различных начальных независимых переменных и затем сравнивались. Попарные корреляции оценок RIP-A оказались высокими (R2> 0,99), показывая, что от различных панелей AIM (рис. S9) и альтернативных выборок населения (рис. S10, S11) выводятся очень схожие оценки этно-популяционного происхождения черт лица. Надежность оценок RIP-А как от маркеров, так и выборок населения доказывает общую применимость и, таким образом, практическую полезность этих моделей. Мы также видим, что оценки RIP-A, полученные с помощью наборов AIM с низкой информативностью по происхождению, демонстрируют более высокие корреляции с более точными оценками происхождения генома, чем с теми, что были использованы для их создания (рис. S9). Для дальнейшей оценки эффективности BRIM в условиях меньшей доступности информации, мы провели эксперименты с внесением шумов путем сложения или вычитания случайно определенных значений из оценок происхождения генома и неверного определения пола в выборке (рис. S4, S5, S6, S7, S8 и рисунки S12, S13, S14, соответственно). Эти эксперименты демонстрируют те же самые паттерны, что указывались выше использования альтернативных AIM: определение точных RIP-переменных для этих двух параметров возможно и при неверном определении пола и неточными оценками происхождения генома. Начальные значения независимых переменных для обоих полов и происхождения могут быть уменьшены в точности на 30% (т.е. r2 = 0,7 между исходной независимой переменной и независимой переменной с введенным шумом) и все еще показывать коэффициенты корреляции около г = 0,95 между значениями RIP-переменных, полученными в условиях шума и значениями RIP-переменных, полученных при первоначальных оценках (рис. S8 и рис S14). BRIM является эффективным инструментом использования скрытой ковариационной структуры лицевых главных компонент для раскрытия диапазона эффектов, оказываемых полом и этно-популяционным происхождением на структуру лица, и может точно определить относительные положения определенных лиц в этих диапазонах по RIP-S и RIP-A соответственно.
Люди также очень умело распознают лица и могут замечать многие аспекты изменчивости среди лиц [21], [22]. Учитывая это, мы пытались проверить, может ли человек-наблюдатель быть средством проверки RIP-A- и RIP-S-переменных. Наблюдателям были показаны 3D анимированные gif-анимации в условных цветах, изображающие лица добровольцев, участвующих в исследовании. Их попросили оценить долю западно-африканского происхождения (от 0% до 100%), а также женственности (по шкале Лайкерта от 1 до 7). Наблюдателей также попросили определить пол и группу населения. Как показано на фиг.5А и 5В, между RIP-А-переменными и значениями доли компоненты происхождения, а также группами населения, определенными наблюдателями, показаны сильные корреляции (всех г> 0,85 и р <0,0001). Показатели RIP-S-переменных и оцененные наблюдателями значения женственности лица и определение ими пола также высоко коррелируют (г> 0,85 и р <0,0001; рисунки 5C и 5D). Эти результаты обеспечивают дополнительное доказательство того, что RIP-A RIP-S являются информативным сводом статистических данных, представляющих относительные уровни этно-популяционного происхождения и женственности лиц. Подобно полу и геномной родословной, -генотипы с маркерами однонуклеотидного полиморфизма (SNP) могут быть использованы в качестве исходных независимых переменных в BRIM, один SNP – это одна RIP-G-переменная. Мы провели частный BRIN-анализ, моделирующий эффект генотипов независимо от пола и происхождения для каждого из 76 SNP-маркеров западноафриканского /европейского происхождения и расположенных в 46 генах-кандидатах, определяющих черепно-лицевую структуру. Эти 46 генов были, в основном, отобраны среди 50 черепно-лицевых генов, которые также демонстрировали геномные сигнатуры ускоренной эволюции по результатам исследования 199 генов (Таблица S2). Поскольку анализ генетических ассоциаций в смешанных популяциях является эффективным подходом обнаружения генов, влияющих на признаки, которые отличаются между популяциями, а RIP-А-анализ является эффективным средством определения общего этно-популяционного происхождения лицевой структуры, пожалуй, несколько контринтуитивно, что в частном BRIM-моделировании отдается большее предпочтение обусловленности со стороны RIP-А-переменных, чем обусловленности со стороны происхождения генома (рис. S15, S16, S17, S18, S19, S20 и S27). Также в частном BRIM-моделировании для оценки RIP-G-переменных RIP-S оказалась лучшей обуславливающей переменной (conditioning variable), чем пол (рис. S21, S22, S23, S24, S25, S26). Мы провели тест ANOVA для проверки средней разности RIP-G-переменных по категориям генотипов (например, CC, CT, и TT кодируется как -1, 0, и 1, что предполагает дополнительные аллельные эффекты). Учитывая существенное априорное доказательство, а именно, что эти гены указывают на наличие ускоренной эволюции в одной или обеих родительских популяциях, и что мутации в этих генах могут привести к явной черепно-лицевой дисморфологии мышей или человека, мы считаем, что для каждого гена должен быть отдельный статистический анализ, и поэтому не требуется корректировки для многократного анализа. Двадцать четыре из 76 RIP-G переменных (в 20 различных генах) показывают р <0,1 (Таблица S2). Относительно низкий порог объясняется сильным априорным доказательством, что RIP-G-анализ для каждого гена характеризует определяемые им черты, и ожидаемый небольшой эффект отдельных генов на нормальный диапазон изменчивости сказывается на лицо в целом. Кроме того, учитывая общий вывод, что клинически значимые гены также могут влиять на субклиническую изменчивость и нормальный диапазон изменчивости (например, [23]), мы постфактум провели подробные описания эффектов этих RIP-G-переменных с помощью FSCPs (рис. S34, S35, показатели S39, S40, S41, S42, S43, S44 и Таблица S4).
(A) RIP-A и оценка доли этно-популяционного происхождения(r = 0.854, p <0.0001), (B) RIP-A и происхождение (r = 0.859, p <0.0001), (C) RIP-S и женственность(r = 0.860, p <0.0001), (D) RIP-S и пол (r = 0.856, p <0.0001). doi:10.1371/journal.pgen.1004224.g005
Характеристика эффектов 3 из этих 24 RIP-G переменных (rs1074265 в SLC35D1, rs13267109 в FGFR1 и rs2724626 в LRP6), представленных на рисунках 6А, 6В и 6С, иллюстрирует эти результаты. Детальный анализ и описание эффектов каждого из 24 однонуклеотидных полиморфных маркеров с помощью FSCPs дано в дополнительном материале (Текст S1). Ген, кодирующий первого члена подгруппы D 35-ой группы белков системы трансмембранных переносчиков SLC (SLC35D1; OMIM # 610804), расположен на хромосоме человека 1p31.3 [24]. Мутации в SLC35D1, как было показано, приводят к дисплазии Шнекенбекена (OMIM # 269250), для которой характерны «к верху ориентированные орбит" (superiorly oriented orbits). Нормальный диапазон изменчивости однонуклеотидного полиморфизма в rs1074265 в SLC35D1 (рис. 6А) демонстрирует сильный эффект, оказываемый на глаза и периорбитальную область, включая заметную разницу в надглазничной области, также как на среднюю зону лица и подбородок. Мутации в гене рецептора 1 факторов роста фибробластов человека (FGFR1; OMIM # 136350), расположенного на хромосоме 8p21.23-p21.22, может привести к четырем аутосомно-доминантным черепно-лицевых нарушениям: синдром Джексон-Вайса (OMIM # 123150), который характеризуется краниостенозом и гипоплазией средней зоны лица; тригоноцефалия (OMIM # 190440), которая характеризуется килевидный лбом, в результате чего сверху череп выглядит треугольным; остеоглофоническая дисплазия (OMIM # 166250), которая характеризуется краниосиностозом надбровных дуг и депрессией корня носа; и синдром Пфайффер (OMIM # 101600), который характеризуется гипоплазией средней зоны лица и, в зависимости от подтипа, глазным экзофтальмом, коротким основанием черепа, и клеверовидным черепом. Нормальный диапазон изменчивости SNP rs13267109 в FGFR1, изображенной на рисунке 6В, показывает сильное влияние на надбровные дуги, глаза, среднюю зону лица, нос и уголки рта. Самые сильные различия в преобразовании формы показывает лоб, надбровные дуги и спинка носа. Мышиный гомолог гена, кодирующего человеческий белок 6, связанный с рецептором липопротеинов низкой плотности (LRP6; OMIM # 603507) который, как известно, является важным для развития губ у мышей, в результате нокаута приводит к образованию двусторонней расщелины губ у мышей [25]. У человека пока еще не описаны черепно-лицевые нарушения, связанные с геном LRP6 или хромосомным участком 12p13.2, хотя продукт гена как известно, взаимодействует на молекулярном уровне с сигнальным путем WNT. При наблюдении за преобразованием формы на рисунке 6С, можно заметить переход от выступающих губ, в том числе утолщенной и выпуклой границы губ, к менее выступающим губам, в том числе, тонкой и менее выпуклой (более вогнутой) границе губ. Это подтверждается путем проверки нормального смещения и карт значений, в которых губы четко очерчены (рис. S43).
Рисунок 6.Карты преобразования и теплокарты, показывающие зависимость формылица от трехRIP-G переменных. Исходные независимые переменные - SNPs в генах (A) SLC35D1 (B) FGFR1, и (C) LRP6. Верхняя строка каждой панели показывает преобразование формы поблизости от крайних значений RIP-G-переменной. Вторая строка показывает R2 (доля общей изменчивости лиц), три исходных параметра изменения формы лица: отношение площади, разность кривизны и нормальное смещение. Максимальные значения R2 для A, B, and C и - 11.68%, 15.16% и 10.10%, соответственно. doi:10.1371/journal.pgen.1004224.g006
В целом, некоторые RIP-G-переменные показывают локализованные эффекты (например, rs1074265 в SLC35D1), изменяя только некоторые аспекты формы лица, в то время как другие показывают изменения в нескольких участках лица (например, rs13267109 в FGFR1). Сводные статистические данные о базовых распределениях величины эффекта по всем опорным точкам представлены в таблице S3. В случае, когда смоделированы несколько однонуклеотидных полиморфных маркеров в одном и том же гене, перекрывающиеся и схожие эффекты показаны для различных маркеров одного и того же гена (например, Dnmt3b и SATB2) и для различных генов, относящихся к одному и тому же биологическому эффекту (например, Wnt3, Fgfr1, и FGFR2). С помощью графического интерфейса пользователя (GUI) эффекты изменений в этих 24 RIP-G переменных, RIP-A, RIP-S, или любой из первых 44 PC-переменных могут быть представлены более подробно. Эти преобразования могут быть визуализированы с помощью текстурной карты с тем же успехом, как и с помощью одной только формы. Кроме того, графический интерфейс (http://tinyurl.com/DNA2FACEIN3D) позволяет иллюстрировать сравнения трансформированных лиц с консенсусными лицами (consensus face) с использованием трех основных FSCPs. Так как категориальные и непрерывные переменные могут быть смоделированы с помощью BRIM, этот подход может быть использован для анализа отношений между чертами лица и другими факторами, например, возрастом, ожирением и темпераментом. Методы, показанные здесь, также обеспечивают развитие диагностических инструментов на основе моделирования подтвержденных случаев явной черепно-лицевой дисморфологии. Наши методы обеспечивают средства идентификации генов, которые влияют на форму лица, и позволяют моделировать влияние этих генов, чтобы сгенерировать предсказанную модель лица. Хотя многое еще предстоит сделать, прежде чем мы узнаем, сколько генов должны определять некоторым образом форму лица, и многие другие группы населения должны быть изучены, прежде чем мы сможем понять, как обобщить полученные данные, эти результаты дают толчок и обеспечивают аналитическую базу последующих исследований.
Дата добавления: 2014-12-11 | Просмотры: 728 | Нарушение авторских прав |
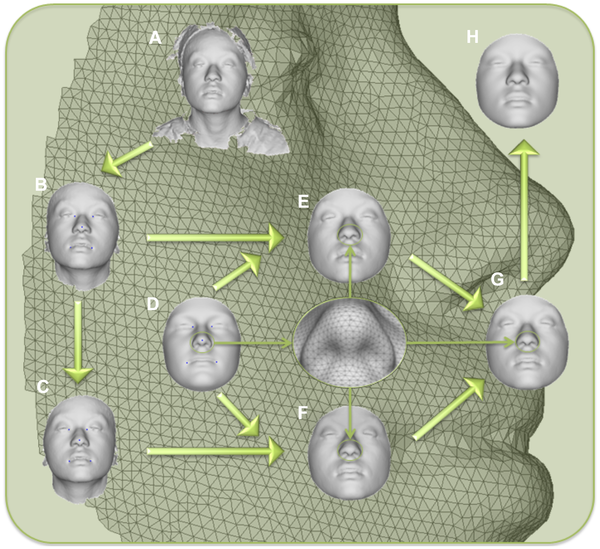 Рисунок 1. Процесс 3D сканирования лица.
Рисунок 1. Процесс 3D сканирования лица.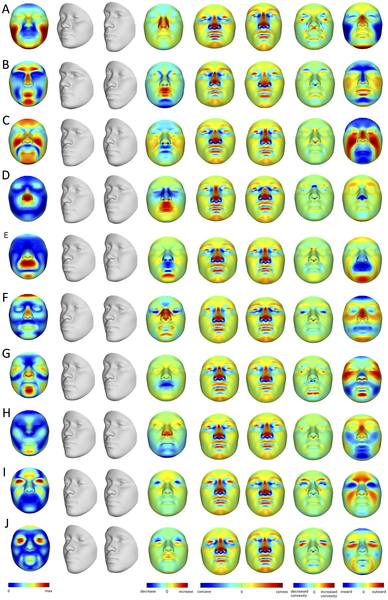 Рисунок 2.ЭффектыPCA, оказываемые на морфологию лица.
Рисунок 2.ЭффектыPCA, оказываемые на морфологию лица.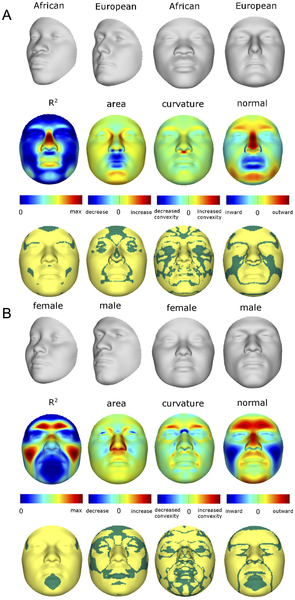 Рисунок 3.Карты преобразования и теплокарты, показывающие зависимость формы лица от (A) RIP-A и (B) RIP-S.
Рисунок 3.Карты преобразования и теплокарты, показывающие зависимость формы лица от (A) RIP-A и (B) RIP-S.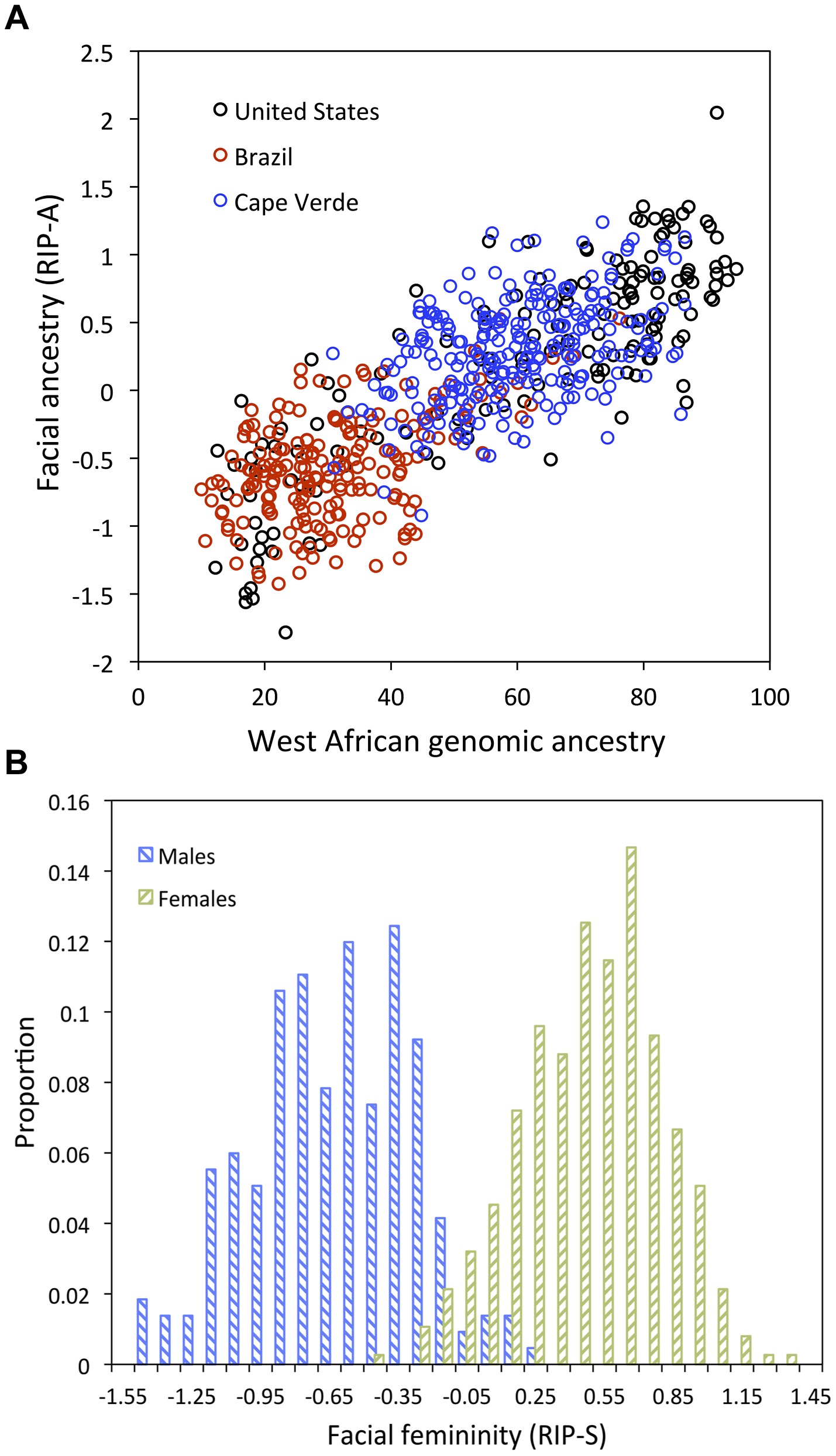 Рисунок 4.Отношения между RIP-переменными происхождения и пола и их первоначальными независимыми переменными.
Рисунок 4.Отношения между RIP-переменными происхождения и пола и их первоначальными независимыми переменными.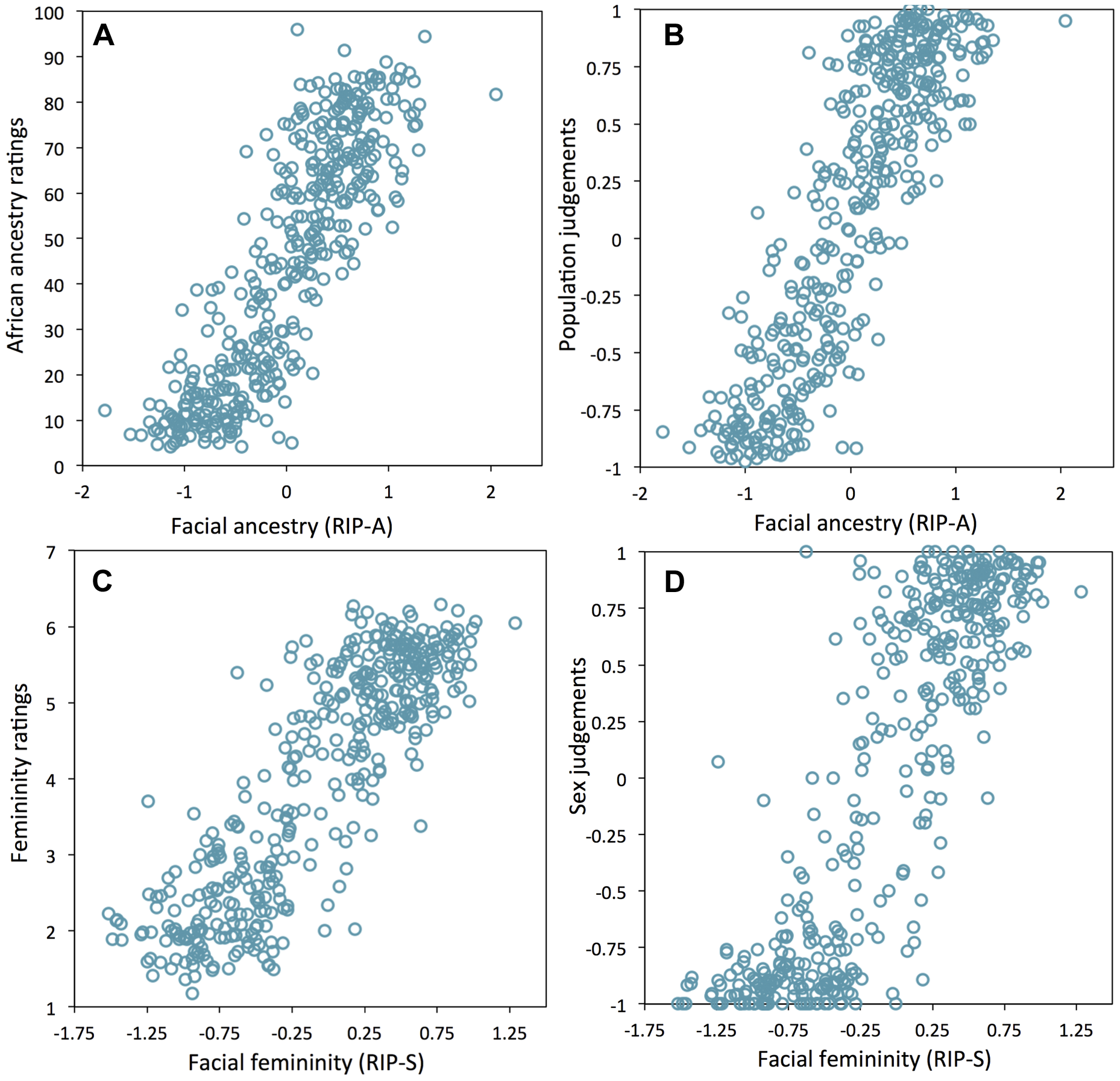 Рисунок 5.Взаимоотношение между количественной оценкой наблюдателей и определением этно-популяционного происхождения и пола.
Рисунок 5.Взаимоотношение между количественной оценкой наблюдателей и определением этно-популяционного происхождения и пола.