|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Деление клетки
Ранее на с. 19, представлены условные обозначения, которые используются при составлении схем митоза и мейоза: рис. 8-9 – максимально спирализованная метафазная хромосома, графическое изображение которой представлено на рис. 3 (разная растушевка морфологически одинаковых хроматид обозначает их несестринское происхождение); рис. 10 и 11 – та же хромосома в полуспирализованном состоянии (несестрин-
Митоз
Митоз – наиболее совершенный и распространенный тип деления клетки, при котором происходит редупликация хромосом и равное их распределение по дочерним клеткам. Длительность периода от одного деления до другого, совокупность процессов, происходящих при этом в клетке, называется митотическим циклом. В процессе митотического цикла происходит сначала удвоение, а затем равномерное распределение наследственного материала, заключенного в хромосомах, между вновь образующимися дочерними клетками. В результате митоза из одной материнской клетки образуется две дочерних с тем же набором хромосом и генетической информацией, что и в исходной материнской клетке. Деление ядра при митозе называется кариокинезом, а деление цитопласта – цитокинезом. Митотический цикл условно разбивается на ряд последовательных фаз: 1) интерфаза; 2) профаза, 3) метафаза, 4) анафаза, 5) телофаза, 6) интерфаза. Между двумя митотическими делениями клетка растет, функционирует, подготавливаясь к последующему митозу. Это состояние клетки называется интерфазой (рис. 18). Обычно масса живого вещества клетки и масса вещества хромосом удваивается. В интерфазе различают 3 периода: пресинтетический, синтетический, на котором происходит редупликация находящейся в составе хромосом ДНК, и постсинтетический. В эту фазу важнейшие компоненты ядра – хромосомы – обычно не удается увидеть в световой микроскоп. Они представлены в виде тонких длинных нитей. В интерфазном ядре весьма интенсивно идут важные биосинтетические процессы, имеющие большое генетическое значение. Продолжительность этой фазы 10-20 часов.
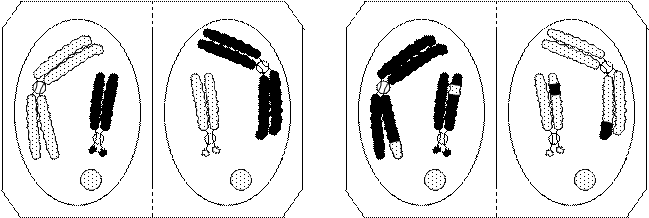
Деление клетки начинается с профазы. В профазе хромосомы хорошо видны. Каждая хромосома профазного ядра состоит из двух нитей – хроматид. Постепенно хромосомы в профазе утолщаются и укорачиваются вследствие спирализации хроматид и превращаются в хорошо видимые в световом микроскопе тельца, интенсивно окрашивающиеся основными красителями, откуда и произошло их название (хромосома по-гречески означает «красящиеся тельца»). К концу профазы хромосомы перемещаются к оболочке ядра (рис. 19). Такое центробежное движение хромосом указывает на приближение конца профазы. В конце профазы оболочка ядра растворяется. Сразу после этого наступает следующая фаза – метафаза. Хромосомы в этот период оказываются в общей массе цито- и кариоплазмы, называемой миксоплазмой. Начинает формироваться митотический аппарат. Он образуется главным образом из белков, синтезируемых в клетке во время интерфазы, и состоит из веретена и экваториальной плоскости. В масштабе клетки митотический аппарат представляет собой крупное образование. В нем имеются полюса, к которым устремляются хромосомы, а его экватор устанавливает плоскость, по которой происходит деление клетки. Хромосомы выстраиваются в плоскости экватора, переходя в метафазу под влиянием натяжения равной силы, исходящего от обоих полюсов (рис. 20). Центромеры располагаются в одной экваториальной плоскости, к ним прикрепляются ахроматиновые нити, связывающие хромосомы с полюсами клетки. Расположение хромосом в экваториальной плоскости называют метафазной пластинкой. Спирализация хромосом в эту фазу достигает максимума. В конце метафазы начинают делиться центромеры. Как только центромеры разделяются, наступает анафаза (рис. 21).
Рис. 20. Метафаза Рис. 21. Анафаза
Хроматиды получают самостоятельность, становясь сестринскими, и расходятся к полюсам. При этом отталкиваются вначале центромеры и хромосомы движутся к полюсам центромерами вперед. Число ахроматиновых нитей соответствует числу хромосом. Эти нити сокращаются и увлекают за собой однохроматидные хромосомы. В конце анафазы хромосомы концентрируются у полюсов, а веретено на экваторе уплотняется и принимает бочонковидную форму. Дочерние хромосомы деспирализуются и утрачивают видимую индивидуальность. Ядро реконструируется в обратном порядке, восстанавливаются ядрышки. В это время начинается симметричное разделение тела клетки. Изменения в обоих ядрах протекают синхронно. Веретено деления разрушается на полюсах и уплотняется в экваториальной зоне, где образуется клеточная стенка (рис. 22). Эта фаза называется телофазой.
Рис. 22. Телофаза
Как только образуется два ядра и клеточная перегородка, ядра переходят в интерфазное состояние (рис. 23). Рис. 23. Интерфаза до воспроизведения хроматид Хромосомы полностью деспирализованы. Здесь происходит воспроизведение хроматид, накопление энергии. Когда клетка достигает определенного размера, она вступает в следующую фазу, то есть начинает делиться вновь. Так идет деление клеток во всех меристемах, камбиальных тканях. В результате из одной клетки образуются две с тем же набором хромосом, что и исходная, т.е. генетически одинаковые.
Мейоз
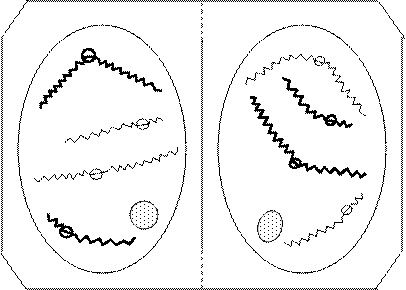
При половом размножении каждый потомок наследует генетический материал от двух родителей. Это, как правило, результат слияния двух половых клеток (гамет). Однако число хромосом в соматических клетках особей данного вида остается постоянным из поколения в поколение, так как половые клетки всегда содержат гаплоидное число хромосом. Образуются клетки с гаплоидным набором хромосом в результате мейоза. Мейоз – это совокупность двух последовательных делений, при котором ядро делится дважды, а хромосомы – один раз, в результате из одной диплоидной клетки возникают четыре гаплоидные. Как и при митозе, важным представляется поведение хромосом (оно показано на рис. 24-31). В основе мейоза лежит тот же митотический цикл, что и в основе митоза – спирализация и деспирализация хромосом. При схематическом изображении основных фаз мейоза использованы те же условные обозначения. Ради простоты проследим за поведением только двух пар хромосом (как при митозе). Интерфаза мейоза ничем не отличается от интерфазы митоза. Начинается мейоз с профазы (рис. 24-25).
Рис. 24. Ранняя профаза Рис. 25. Поздняя профаза
В самом начале профазы первого деления мейоза (профаза I) хромосомы представлены длинными тонкими нитями. Эта стадия называется лептонема. Затем хромосомы начинают спирализоваться и сближаться гомологичными участками по всей длине. Гомологичные хромосомы объединяются попарно. Процесс попарного сближения хромосом называется конъюгацией, а стадия, на которой это происходит, называется стадией зигонемы, или стадией соединения нитей. Еще позднее они полностью сливаются между собой, образуя двойные нити. Комплекс, состоящий из двух конъюгирующих хромосом, называется бивалентом, а каждая из составляющих хромосом – унивалентом. Эта стадия – стадия пахинемы, или стадия толстых нитей. Далее спирализация хромосом усиливается, они утолщаются и хорошо видны под микроскопом. Хромосомы оказываются уже значительно спирализованными, центромеры гомологичных хромосом начинают отталкиваться друг от друга (рис. 25), начинается расщепление сблизившихся хромосом. При расхождении центромер (поздняя профаза) точки соприкосновения несестринских хроматид сползают к концам хромосом. Образуются иксообразные фигуры, называемые хиазмами. Это стадия диплонемы, или стадия двойных нитей. Заключительная стадия профазы I называется стадией диакинеза. Хиазмы при этом смещаются к концам хромосом (терминализация хиазм). При конъюгации или разрыве хиазм несестринские хроматиды могут обмениваться гомологичными участками. Это явление называется перекрестом, или кроссинговером, и играет важную генетическую роль. Окончание профазы и переход к метафазе связаны с растворением ядра и формированием ахроматинового веретена. В экваториальной плоскости метафазы I (рис. 26) располагаются биваленты. Их местонахождение случайно по отношению друг к другу, как и ориентация унивалентов к полюсам.
Рис. 26. Метафаза I К одному полюсу могут ориентироваться униваленты разного происхождения. Но возможны и другие сочетания. Число этих возможных сочетаний тем больше, чем больше у организма хромосом. Вслед за метафазой первого деления наступает анафаза I. В отличие от митоза анафаза I характеризуется не разделением центромер, а разрывом последних хиазм, поэтому к полюсам отходят не однохроматидные, а двухроматидные хромосомы (рис. 27). В результате у каждого полюса оказывается гаплоидный набор хромосом.
Рис. 27. Анафаза I Телофаза I отличается от телофазы митоза только тем, что деспирализация хромосом здесь не доходит до конца. Разделившиеся хромосомы обособляются в два гаплоидных ядра (рис. 28).
Иногда в этой фазе происходит цитокинез (сукцессивный, т.е. неодновременный тип деления), иногда он наблюдается только после второго мейотического деления (симультантный, одновременный тип деления). Телофаза I протекает недолго и не доходит до конца. Первое деление еще не закончилось, но уже начинается второе. Между первым и вторым делением нет интерфазы. Хромосомы не успевают деспирализоваться, а уже начинают вновь закручиваться. Наступает особая фаза, называемая интеркинезом (см. рис. 28). Она отличается от интерфазы не только тем, что хромосомы подвергаются лишь частичной деспирализации, но и тем, что в клетке не осуществляется обычный для интерфазы синтез, в том числе не синтезируется вещество хромосом. Итак, в результате первого деления каждая из двух клеток получает по набору хромосом. Поскольку пара хромосом расходится по полюсам независимо от других пар, то любая гаплоидная клетка может содержать произвольную рекомбинацию родительских хромосом. Чем больше число хромосом, тем больше возможное число рекомбинаций. При двух парах хромосом возможны четыре соединения – два родительских и два рекомбинантных, для 23 пар (как у человека) число возможных соединений равно 223 = 8388608. Поскольку в мейозе рекомбинируются между собой не только хромосомы, но и отдельные участки гомологичных хромосом, то качественное генотипическое разнообразие гаплоидных клеток на самом деле большее. После короткого интеркинеза в каждой из двух клеток, образовавшихся при первом делении, одновременно происходит митоз, или второе деление. Начинается оно с профазы, которая отличается от профазы митоза тем, что характеризуется более значительной спирализацией хромосом. Во всех остальных делениях этот механизм ничем не отличается от митоза. Схематическое изображение метафазы второго деления представлено на рис. 29.
Хромосомы выстраиваются своими центромерами в экваториальной плоскости. В отличие от митоза, число их уменьшается в два раза. В анафазе II происходит деление центромеры хромосом. Сестринские хромосомы (монады) расходятся к полюсам, образуя генотипически неравноценные дочерние клетки (рис. 30).
Рис. 30. Анафаза II
В телофазе II (рис. 31) завершается расхождение хромосом к полюсам, каждый из них получает по набору хромосом, вокруг которого формируется ядерная оболочка. В каждой из четырех образовавшихся клеток вдвое меньше (в сравнении с исходной) не только число хромосом, но и количество ДНК. Рис. 31. Телофаза II
Первое мейотическое деление, приводящее к переходу от диплоидного ядра к гаплоидному, часто называют редукционным, второе мейотическое деление – эквационным. Таким образом, мейоз в такой же мере увеличивает наследственную изменчивость в половом потомстве, в какой соматический митоз предотвращает ее возникновение в пределах индивида и в вегетативном потомстве исходного организма. Кроме уменьшения вдвое исходного числа хромосом, в результате мейоза изменчивость увеличивается еще как от свободного комбинирования отцовских и материнских хромосом, так и от перестройки их структуры в результате кроссинговера, происходящего в профазе I. Мейоз с его стадиями и фазами развития половых клеток является лишь одним из этапов процесса полового размножения. После мейоза наступает этап формирования зрелых половых клеток.
Дата добавления: 2015-01-18 | Просмотры: 2845 | Нарушение авторских прав |