|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Физиология нервной клеткиФизиология центральной нервной системы ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ Центральную нервную систему составляют спинной и головной мозг. Основными функциями центральной нервной системы являются: 1) регуляция деятельности всех тканей и органов и объединение их в единое целое; 2) обеспечение приспособления организма к условиям внешней среды (организация адекватного поведения соответственно потребностям организма). Управление различными функциями осуществляется и гуморальным путем (через кровь, лимфу, тканевую жидкость), однако нервная система играет главенствующую роль. У высших животных и человека ведущим отделом центральной нервной системы является кора больших полушарий, которая управляет также наиболее сложными функциями в жизнедеятельности человека — психическими процессами (сознание, мышление, память и др.). Физиология нервной клетки
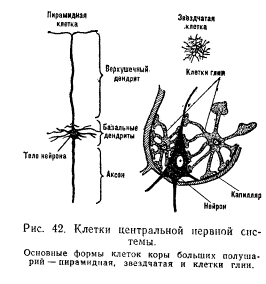
Нейроны разделяются на три основных типа: афферентные, эфферентные и промежуточные нейроны. Афферентные нейроны (чувствительные, или центростремительные) передают информацию от рецепторов в центральную нервную систему. Тела этих нейронов расположены вне центральной нервной системы — в спинномозговых ганглиях и в ганглиях черепно-мозговых нервов. Афферентный нейрон имеет ложноуниполярную форму, т. е. оба его отростка выходят из одного полюса клетки. Далее нейрон разделяется на длинный дендрит, образующий на периферии воспринимающее образование — рецептор, и аксон, входящий через задние рога в спинной мозг. К афферентным нейронам относят также нервные клетки, аксоны которых составляют восходящие пути спинного и головного мозга. Эфферентные нейроны (центробежные) связаны с передачей нисходящих влияний от вышележащих этажей нервной системы к нижележащим (например, пирамидные нейроны коры больших полушарий — рис. 42) или из центральной нервной системы к рабочим органам (например, в передних рогах спинного мозга расположены тела двигательных нейронов, или мотонейронов, от которых идут волокна к скелетным мышцам; в боковых рогах спинного мозга находятся клетки вегетативной нервной системы, от которых идут пути к внутренним органам). Для эфферентных нейронов характерны разветвленная сеть дендритов и один длинный отросток — аксон. Промежуточные нейроны (интернейроны, или вставочные) — это, как правило, более мелкие клетки, осуществляющие связь между различными (в частности, афферентными и эфферентными) нейронами. Они передают нервные влияния в горизонтальном направлении (например, в пределах одного сегмента спинного мозга) и в вертикальном (например, из одного сегмента спинного мозга в другие — выше или нижележащие сегменты). Благодаря многочисленным разветвлениям аксона промежуточные нейроны могут одновременно возбуждать большое число других нейронов (например, звездчатые клетки коры — см. рис. 42). Функциональное значение различных структурных элементов нервной клетки. Различные структурные элементы нейрона имеют свои функциональные особенности и разное физиологическое значение. Нервная клетка состоит из тела, или сомы (рис. 43), и различных отростков. Многочисленные древовидно разветвленные отростки дендриты (от греч. dendron — дерево) служат входами нейрона, через которые сигналы поступают в нервную клетку. Выходом нейрона является отходящий от тела клетки отросток аксон (от греч. axis — ось), который передает нервные импульсы дальше — другой нервной клетке или рабочему органу (мышце, железе). Форма нервной клетки, длина и расположение отростков чрезвычайно разнообразны и зависят от функционального назначения нейрона.
В крупных нейронах почти 1/3 — 1/4 величины их тела составляет ядро. Оно содержит довольно постоянное количество дезоксирибонуклеиновой кислоты (ДНК). Входящие в его состав ядрышки участвуют в снабжении клетки рибонуклеиновыми кислотами (РНК) и протеинами. В моторных клетках при двигательной деятельности ядрышки заметно увеличиваются в размерах. Нервная клетка покрыта плазматической мембраной—полупроницаемой клеточной оболочкой, которая обеспечивает регуляцию концентрации ионов внутри клетки и ее обмен с окружающей средой. При возбуждении проницаемость клеточной мембраны изменяется, что играет важнейшую роль в возникновении потенциала действия и передаче нервных импульсов. Аксоны многих нейронов покрыты миелиновой оболочкой, образованной Шванновскими клетками, многократно «обернутыми» вокруг ствола аксона. Однако начальная часть аксона и расширение в месте его выхода из тела клетки — аксонный холмик лишены такой оболочки. Мембрана этой немиелинизированной части нейрона — так называемого начального сегмента — обладает высокой возбудимостью. Внутренняя часть клетки заполнена цитоплазмой, в которой расположены ядро и различные органоиды. Цитоплазма очень богата ферментными системами (в частности, обеспечивающими гликолиз) и белком. Ее пронизывает сеть трубочек и пузырьков — эндоплазматический ретикулюм. В цитоплазме имеются также отдельные зернышки — рибосомы и скопления этих зернышек — тельца Ниссля, представляющие собой белковые образования, содержащие до 50% РНК. Это белковые депо нейронов, где также происходит синтез белков и РНК. При чрезмерно длительном возбуждении нервной клетки, вирусных поражениях центральной нервной системы и других неблагоприятных воздействиях величина этих рибосомных зернышек резко уменьшается. В специальных аппаратах нервных клеток — митохондриях совершаются окислительные процессы с образованием богатых энергией соединений (макроэргических связей АТФ). Это энергетические станции нейрона. В них происходит трансформация энергии химических связей в такую форму, которая может быть использована нервной клеткой. Митохондрии концентрируются в наиболее активных частях клетки. Их дыхательная функция усиливается при мышечной тренировке. Интенсивность окислительных процессов нарастает в нейронах более высоких отделов центральной нервной системы, особенно в коре больших полушарий. Резкие изменения митохондрий вплоть до разрушения, а следовательно, и угнетение деятельности нейронов отмечаются при различных неблагоприятных воздействиях (длительном торможении в центральной нервной системе, при интенсивном рентгеновском облучении, кислородном голодании и гипотермии). Обмен веществ в нейроне. Основной особенностью обмена веществ в нейроне является высокая скорость обмена и преобладание аэробных процессов. Потребность мозга в кислороде очень велика (в состоянии покоя поглощается около 46 мл/мин кислорода). Хотя вес мозга по отношению к весу тела составляет всего 2%, потребление кислорода мозгом достигает в состоянии покоя у взрослых людей 25% от общего его потребления организмом, а у маленьких детей — 50%. Даже кратковременное нарушение доставки кислорода кровью может вызвать необратимые изменения в деятельности нервных клеток: в спинном мозгу — через 20 — 30 мин., в стволе головного мозга — через 15 — 20 мин., а в коре больших полушарий — уже через 5 — 6 мин. Энерготраты мозга составляют 1/6 — 1/8 суточных затрат организма человека. Основным источником энергии для мозговой ткани является глюкоза. Мозг человека требует для обмена около 115 г глюкозы в сутки. Содержание ее в клетках мозга очень мало, и она постоянно черпается из крови. Деятельное состояние нейронов сопровождается трофическими процессами—усилением в них синтеза белков. При различных воздействиях, вызывающих возбуждение нервных клеток, в том числе при мышечной тренировке, в их ткани значительно возрастает количество белка и РНК, при тормозных же состояниях и утомлении нейронов содержание этих веществ уменьшается. В процессе восстановления оно возвращается к исходному уровню или превышает его. Часть синтезированного в нейроне белка компенсирует его расходы в теле клетки во время деятельности, а другая часть перемещается вдоль по аксону (со скоростью около 1—3 мм в сутки) и, вероятно, участвует в биохимических процессах в синапсах. Кровоснабжение нервных клеток. Высокая потребность нейронов в кислороде и глюкозе обеспечивается интенсивным кровотоком. Кровь протекает через мозг в 5—7 раз скорее, чем через покоящиеся мышцы. Мозговая ткань обильно снабжена кровеносными сосудами. Наиболее густая сеть их находится в коре больших полушарий (занимает около 10% объема коры). В отдельных слоях ее средняя длина капиллярной сети достигает у человека 1 м на 1 мм3 ткани. Каждый крупный нейрон имеет несколько собственных капилляров у основания тела клетки, а группы мелких клеток окутаны общей капиллярной сетью. При активном состоянии нервной клетки она нуждается в усиленном поступлении через кровь кислорода и питательных веществ. Вместе с тем жесткий каркас черепа и малая сжимаемость нервной ткани препятствуют резкому увеличению кровоснабжения мозга при работе. Однако это компенсируется выраженными в мозгу процессами перераспределения крови, в результате которых активный участок нервной ткани получает значительно больше крови, чем находящийся в покое. Возможность перераспределения крови в мозгу обеспечена наличием в основаниях артериальных ветвей крупных пучков гладких мышечных волокон — сфинктерных валиков. Эти валики могут уменьшать или увеличивать диаметр сосудов и тем самым производить раздельную регуляцию кровоснабжения разных участков мозга. Мышечная работа вызывает снижение тонуса стенок мозговых артерий. При развитии физического или умственного утомления тонус артериальных сосудов повышается, что ведет к уменьшению кровотока через нервную ткань. В головном мозгу имеется богато развитая система анастомозов между различными артериями, между венозными сосудами и между артериями и венами. Эта система уменьшает пульсацию внутричерепного кровотока, обусловленную ритмическими сокращениями сердца и дыхательными движениями грудной клетки. Уменьшение пульсовых колебаний способствует улучшению тканевого кровотока. Благодаря наличию артериовенозных анастомозов пульсовые колебания кровотока передаются с артерий мозга на вены, минуя капилляры. Анастомоз между системами сонных и позвоночных артерии (Виллизиев круг) гарантирует постоянство кровотока в различных отделах головного мозга при любом положении головы по отношению к туловищу и направлению силы тяжести, связанном с изменением положения тела в пространстве. Клетки глии. В процессах питания нервных клеток и их обмене веществ участвуют также окружающие нейрон клетки глии (глиальные клетки, или нейроглия). Эти клетки заполняют в мозгу все пространство между нейронами. В коре больших полушарий их примерно в 5 раз больше, чем нервных клеток. Капилляры в центральной нервной системе плотно окружены клетками глии, которые покрывают сосуд полностью или оставляют небольшую часть (15%) свободной. Выросты некоторых глиальных клеток расположены частично на кровеносных сосудах и частично на нейроне (см. рис. 42). Полагают, что расположение этих клеток между сосудом и нейроном указывает на их роль в снабжении нервных клеток питательными веществами из крови. Глиальные клетки активно участвуют в функционировании нейрона. Показано, что при длительном возбуждении в нейроне высокое содержание белка и нуклеиновых кислот поддерживается за счет клеток глии, в которых их количество соответственно уменьшается. В процессе восстановления после работы запасы белка и нуклеиновых кислот сначала нарастают в клетках глии, а затем и в цитоплазме нейрона. Глиальные клетки обладают способностью перемещаться в пространстве по направлению к наиболее активным нейронам. Это наблюдается при различных афферентных раздражениях и при мышечной нагрузке. Например, уже через 20 мин. плавания у крыс было обнаружено увеличение числа глиальных клеток вокруг мотонейронов переднего рога спинного мозга. Возможно, клетки глии участвуют в условно-рефлекторной деятельности мозга и в процессах памяти. Участвуют, но не функционально, а как вспомогательное окружение. Механизмы рефлексов функционально полностью описываются свойствами нейронов и синапсов. Основные функции нервной клетки. Основными функциями нервной клетки являются восприятие внешних раздражении (рецепторная функция), их переработка (интегративная функция) и передача нервных влияний на другие нейроны или различные рабочие органы (эффекторная функция). Особенности осуществления этих функций позволяют разделить все нейроны центральной нервной системы на 2 большие группы: 1) клетки, передающие информацию на большие расстояния (из одного отдела центральной нервной системы в другой, от периферии к центру, от центров к исполнительному органу). Это крупные, афферентные и эфферентные нейроны, имеющие на своем теле и отростках большое количество синапсов, как возбуждающих, так и тормозящих, и способные к сложным процессам переработки поступающих через них влиянии; 2) клетки, обеспечивающие межнейроальные связи в пределах ограниченных нервных структур (промежуточные нейроны спинного мозга, коры больших полушарий и др.). Это мелкие клетки, воспринимающие нервные влияния только через возбуждающие синапсы. Эти клетки не способны к сложным процессам интеграции локальных синаптических влияний потенциалов, они служат передатчиками возбуждающих или тормозящих влияний на другие нервные клетки. Воспринимающая функция нейрона. Все раздражения, поступающие в нервную систему, передаются на нейрон через определенные участки его мембраны, находящиеся в области синаптических контактов. В большинстве нервных клеток эта передача осуществляется химическим путем с помощью медиаторов. Ответом нейронов на внешнее раздражение является изменение величины, мембранного потенциала. Чем больше синапсов на Чем сложнее интегративная функция нейрона, тем большее развитие имеют аксодендритические синапсы (в первую очередь те, которые расположены на шипиках). Особенно они характерны для нейрональных связей пирамидных клеток в коре больших полушарий. Промежуточные нейроны (например, звездчатые клетки коры) таких шипиков лишены. Приходящие в пресинаптическую часть контакта нервные импульсы вызывают опорожнение синоптических пузырьков с выведением медиатора в синаптическую щель (рис. 45). Веществами, передающими нервные влияния в синапсах нервных клеток, или медиаторами, могут быть ацетилхолин (в некоторых клетках спинного мозга, в вегетативных ганглиях), норадреналин (в окончаниях симпатических нервных волокон, в гипоталамусе), некоторые аминокислоты и др. Диаметр пузырьков примерно равен ширине синаптической щели. В клетках передней центральной извилины коры больших полушарий у людей 18 — 30 лет синаптические пузырьки имеют диаметр 250 — 300 ангстрем при ширине синаптической щели 200 — 300 ангстрем. Выделение медиатора облегчается тем, что синаптические пузырьки скапливаются вблизи от синаптической щели— в так называемых активных, или оперативных, зонах. Чем больше нервных импульсов проходит через синапс, тем больше пузырьков перемещается в эту зону и прикрепляется к пресинаптической мембране. В результате облегчается выделение медиатора последующими нервными импульсами. Эффекты, возникающие при активации синапса, могут быть возбуждающими или тормозящими. Это зависит от качества медиатора и свойств постсинаптической мембраны. Возбуждающие нейроны выделяют возбуждающий медиатор, а тормозные — тормозной. Кроме того, один и тот же медиатор может оказывать различное воздействие в разных органах (например ацетилхолин возбуждает скелетные мышечные волокна и тормозит сердечные). В состоянии покоя мембрана поляризована: снаружи регистрируется положительный заряд, а внутри — отрицательный. В нервной клетке мембранный потенциал покоя равен примерно 70 мв. При возбуждающих влияниях увеличение проницаемости мембраны обусловливает вхождение положительно заряженных ионов натрия в клетку и, следовательно, уменьшение разности потенциалов по обе стороны мембраны, т. е. ее деполяризацию. В постсинаптической мембране данного участка клетки при этом регистрируется небольшое отрицательное колебание мембранного потенциала с амплитудой около 10 мв, или возбуждающий постсинаптический потенциал (сокращенно ВПСП), нарастающий примерно за 1, 2 мсек. до своего максимума и затем убывающий. При тормозных воздействиях проницаемость мембраны увеличивается незначительно — главным образом для ионов калия (диаметр гидратированного иона калия меньше, чем диаметр иона натрия). Поскольку ионов калия больше внутри клетки, они выходят из нее наружу, увеличивая поляризацию мембраны, т. е. вызывают ее гиперполяризацию. При этом регистрируется положительное колебание с амплитудой около 5 мв — тормозной постсинаптический потенциал (сокращенно ТПСП). Обычно ВПСП и ТПСП длятся всего несколько миллисекунд. Однако в некоторых нейронах головного мозга постсинаптические потенциалы могут иметь значительно большую длительность: ВПСП — до 80 мсек., ТПСП — более 100 мсек. Интегративная функция нейрона. Общее изменение мембранного потенциала нейрона является результатом сложного взаимодействия (интеграции) местных ВПСП и ТПСП всех многочисленных активированных синапсов на теле и дендритах клетки. На мембране нейрона происходит процесс алгебраического суммирования положительных и отрицательных колебаний потенциала. При одновременной активации нескольких возбуждающих синапсов общий ВПСП нейрона представляет собой сумму отдельных местных ВПСП каждого синапса. При одновременном возникновении двух различных синаптических влияний — ВПСП и ТПСП — происходит взаимное вычитание их эффектов. В конечном итоге реакция нервной клетки определяется суммой всех синаптических влияний. Преобладание тормозных синаптических воздействий приводит к гиперполяризации мембраны и торможению деятельности клетки. При сдвиге мембранного потенциала в сторону деполяризации повышается возбудимость клетки. Ответный разряд нейрона возникает лишь тогда, когда изменения мембранного потенциала достигают порогового значения — критического уровня деполяризации. Для этого величина ВПСП клетки должна составлять примерно 10 мв. В крупных (афферентных и эфферентных) нейронах возбудимость различных участков мембраны неодинакова. В области начального сегмента нервной клетки (аксонного холмика и начальной немиелинизированной части аксона) имеется низкопороговая зона, мембрана которой обладает в несколько раз более высокой возбудимостью, чем на других участках клетки (порог возбудимости мембраны начального сегмента равен 10 мв, а порог возбудимости соматодендритической мембраны—20—30 мв). В этой зоне с момента достижения критического уровня деполяризации начинается лавинообразное вхождение натрия в клетку и регистрируется потенциал действия (ПД). В ПД различают кратковременную высоковольтную часть, или спайк (пик), и длительные низкоамплитудные колебания — следовые потенциалы. ПД мотонейронов имеют амплитуду пика около 80 — 100 мв и длительность его около 1,5 мсек. Эффекторная функция нейрона. С появлением ПД, который в отличие от местных изменений мембранного потенциала (ВПСП и ТПСП) является распространяющимся процессом, нервный импульс начинает проводиться от тела нервной клетки вдоль по аксону к другой нервной клетке или рабочему органу, т. е. осуществляется эффекторная функция нейрона. Синапсы, расположенные ближе к возбудимой низкопороговой зоне на теле клетки (аксосоматические), оказывают большее влияние на возникновение потенциала действия, чем более удаленные, расположенные на окончаниях дендритов (аксодендритические). Импульсы, приходящие через аксосоматический синапс, как правило, вызывают ответный разряд нейрона, а импульсы, действующие на аксодендритический синапс —лишь подпороговое изменение его возбудимости. Так, разряды мотонейронов спинного мозга и пирамидных нейронов коры, вызывающие двигательные реакции организма, являются ответом на специфические аксосоматические влияния. Но возникнет ли этот ответ или нет, определяется характером воздействий, поступающих через аксодендритические синапсы от других нервных путей. Так складываются адекватные реакции, зависящие от многих раздражении, действующих на организм в данный момент времени, и осуществляется тонкое приспособление поведения к меняющимся условиям внешней среды. Процессы, происходящие в активном нейроне, можно представить в виде следующей цепи: потенциал действия в пресинаптическом окончании предыдущего нейрона —> выделение медиатора в синаптическую щель —> увеличение проницаемости постсинаптической мембраны —> ее деполяризация (ВПСП) или гиперполяризация (ТПСП) —> взаимодействие ВПСП и ТПСП на мембране сомы и дендритов нейрона —> сдвиг мембранного потенциала в случае преобладания возбуждающих влияний —> достижение критического уровня деполяризации —> возникновение потенциала действия в низкопороговой зоне (мембране начального сегмента) нейрона —> распространение потенциала действия вдоль по аксону (процесс проведения нервного импульса) —> выделение медиатора в окончаниях аксона (передача нервного процесса на следующий нейрон или на рабочий орган). Таким образом, передача информации в нервной системе происходит с помощью двух механизмов — электрического (ВПСП, ТПСП, потенциалы действия) и химического (медиаторы). Дата добавления: 2015-02-02 | Просмотры: 1333 | Нарушение авторских прав |
 Основным структурным элементом нервной системы является нервная клетка, или нейрон. Через нейроны осуществляется передача информации от одного участка нервной системы к другому, обмен информацией между нервной системой и различными участками тела. В нейронах происходят сложнейшие процессы обработки информации. С их помощью формируются ответные реакции организма (рефлексы) на внешние и внутренние раздражения.
Основным структурным элементом нервной системы является нервная клетка, или нейрон. Через нейроны осуществляется передача информации от одного участка нервной системы к другому, обмен информацией между нервной системой и различными участками тела. В нейронах происходят сложнейшие процессы обработки информации. С их помощью формируются ответные реакции организма (рефлексы) на внешние и внутренние раздражения. Среди нейронов встречаются самые крупные клеточные элементы организма. Размеры их поперечника колеблются от 6—7 мк (мелкие зернистые клетки мозжечка) до 70 мк (моторные нейроны головного и спинного мозга). Плотность их расположения в некоторых отделах центральной нервной системы очень велика. Например, в коре больших полушарий человека на 1 мм3 приходится почти 40 тыс. нейронов. Тела и дендриты нейронов коры занимают в целом примерно половину объема коры.
Среди нейронов встречаются самые крупные клеточные элементы организма. Размеры их поперечника колеблются от 6—7 мк (мелкие зернистые клетки мозжечка) до 70 мк (моторные нейроны головного и спинного мозга). Плотность их расположения в некоторых отделах центральной нервной системы очень велика. Например, в коре больших полушарий человека на 1 мм3 приходится почти 40 тыс. нейронов. Тела и дендриты нейронов коры занимают в целом примерно половину объема коры. нервной клетке, тем больше воспринимается различных раздражений и, следовательно, шире сфера влияний на ее деятельность и возможность участия нервной клетки в разнообразных реакциях организма. На телах крупных мотонейронов спинного мозга насчитывают до 15000—20000 синапсов. Разветвления аксонов могут образовывать синапсы на дендритах (аксодендритические синапсы) и на соме (теле) нервных клеток (аксосоматические синапсы), в ряде случаев—на аксоне (аксоаксональные синапсы). Наибольшее число (до 50%) синапсов находится на дендритах. Особенно густо они покрывают средние части и окончания дендритных отростков, причем многие контакты расположены на специальных шипиковидных выростах, или шипиках (рис. 44), которые еще больше увеличивают воспринимающую поверхность нейрона. В мотонейронах спинного мозга и пирамидных клетках коры поверхность дендритов в 10—20 раз больше поверхности тела клетки.
нервной клетке, тем больше воспринимается различных раздражений и, следовательно, шире сфера влияний на ее деятельность и возможность участия нервной клетки в разнообразных реакциях организма. На телах крупных мотонейронов спинного мозга насчитывают до 15000—20000 синапсов. Разветвления аксонов могут образовывать синапсы на дендритах (аксодендритические синапсы) и на соме (теле) нервных клеток (аксосоматические синапсы), в ряде случаев—на аксоне (аксоаксональные синапсы). Наибольшее число (до 50%) синапсов находится на дендритах. Особенно густо они покрывают средние части и окончания дендритных отростков, причем многие контакты расположены на специальных шипиковидных выростах, или шипиках (рис. 44), которые еще больше увеличивают воспринимающую поверхность нейрона. В мотонейронах спинного мозга и пирамидных клетках коры поверхность дендритов в 10—20 раз больше поверхности тела клетки.