|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Свойства нервных центровПонятие о нервном центре. В сложных многоклеточных организмах животных и человека отдельная нервная клетка не в состоянии регулировать какие-либо функции. Все основные формы деятельности нервной системы связаны с участием в ее функциях определенных групп нервных клеток—нервных центров. Нервным центром называют совокупность нервных клеток, необходимых для осуществления какой-либо функции. Эти центры отвечают соответствующими рефлекторными реакциями на внешнее раздражение, поступившее от связанных с ними рецепторов. Клетки нервных центров реагируют и на непосредственное их раздражение веществами, находящимися в протекающей через них крови (гуморальные влияния). Для выявления функций нервного центра используют ряд методов: метод раздражения (усиление его деятельности), метод разрушения или удаления — экстирпации (нарушение регулируемой им функции), электрофизиологический метод — регистрация электрических явлений в нервном центре и др. Так, функциональное значение, а также локализация дыхательного центра в продолговатом мозгу были выявлены путем последовательных перерезок головного мозга. При перерезках выше области его расположения (выше продолговатого мозга) дыхание сохранялось, а при перерезках ниже продолговатого мозга дыхательные движения исчезали. Сложные реакции в целостном организме обычно связаны с участием многих нервных центров, расположенных в различных этажах центральной нервной системы. Например, произвольное изменение дыхания осуществляется человеком при участии центров коры больших полушарий, особого центра в среднем мозгу, дыхательного центра продолговатого мозга и центров спинного мозга, иннервирующих дыхательные мышцы. Физиологические свойства нервных центров определяют характер ответных реакций. Эти свойства в значительной мере связаны с особенностями проведения нервных импульсов через синапсы, соединяющие различные нервные клетки. Одностороннее проведение возбуждения. В отличие от нервного волокна, в котором возможно двустороннее проведение волны возбуждения от места нанесения раздражения, в нервных центрах волна возбуждения проводится только в одном направлении: раздражение задних (чувствительных) корешков спинного мозга вызывает возникновение нервного импульса в передних двигательных корешках, однако при раздражении передних корешков волна возбуждения в задних корешках не появляется. Это обусловлено особенностями синаптической передачи возбуждения в отдельных нервных клетках. Поскольку проведение волны возбуждения от одного нейрона к другому через синапс происходит химическим путем—с помощью медиатора, а медиатор содержится лишь в пресинаптической части синапса и отсутствует в постсинаптической мембране, — проведение нервных влияний через синапс возможно только от пресинаптической мембраны к постсинаптической и невозможно в обратном направлении (см. рис. 45). В связи с этим поток нервных импульсов в рефлекторной дуге имеет определенное направление: от афферентных нейронов к вставочным и затем к эффекторным — мотонейронам и вегетативным нейронам. Замедленное проведение возбуждения. В основе проведения нервных импульсов по цепочке нейронов лежат два различных механизма: электрический (проведение потенциала действия — ПД — по нервным волокнам) и химический (передача через синапс с помощью медиатора). Первый осуществляется с большой скоростью (до 100—140 м/сек), второй—в тысячу раз медленнее. Замедление проведения связано с затратой времени на процессы, происходящие от момента прихода пресинаптического импульса в синапс до появления в постсинаптической мембране возбуждающих или тормозных потенциалов. Этот интервал называется синаптической задержкой и составляет в мотонейронах и большинстве других клеток Центральной нервной системы примерно 0,3 мсек. В вегетативной нервной системе длительность синаптической задержки больше — не менее 10 мсек. За это время пресинаптический импульс вызывает опорожнение синаптических пузырьков, происходит диффузия медиатора через синаптическую щель, увеличение под его влиянием ионной проницаемости постсинаптической мембраны и возникает постсинаптический потенциал. После этого требуется еще около 1,2 мсек. на развитие возбуждающего постсинаптического потенциала (ВПСП) до максимума, чтобы возник (в случае достижения критического уровня деполяризации) ПД. Таким образом, весь процесс передачи нервного импульса (от ПД одной клетки до ПД следующей клетки) через один синапс занимает примерно 1,5 мсек. При утомлении, охлаждении и ряде других воздействий длительность синаптической задержки возрастает. Если же для осуществления какой-либо реакции требуется участие большого числа нейронов (многих сотен и даже тысяч), то суммарная величина задержки проведения по нервным центрам — так называемое центральное время проведения — может составить десятые доли секунды и даже целые секунды. Поскольку проведение импульсов по нервным
Суммация. В ответ на одиночную афферентную волну, идущую от мышц к мотонейронам, в пресинаптической части синапса освобождается 1 квант нейромедиатора. При этом в пресинаптической мембране мотонейрона обычно возникает ВПСП подпороговой величины – 0,1 – 0,2 мв, недостаточной для вызова ответной реакции клетки. Чтобы величина ВПСП достигла критического уровня (порога возникновения ПД – примерно 10 мв.), требуется суммация на мембране клетки многих подпороговых ВВСП. Различают пространственную и временную суммацию. Пространственная суммация наблюдается в случае одновременного поступления нескольких импульсов в один и тот же нейрон по разным пресинаптическим волокнам. Одновременное возбуждение синапсов в различных участках мембраны мотонейрона повышает амплитуду суммарного ВПСП до пороговой величины, возникает ответный импульс мотонейрона и рефлекторно осуществляется двигательная реакция. Так, при синхронном раздражении трех афферентных нервах волокон—от короткой малоберцовой мышцы, длинного разгибателя пальцев и длинной малоберцовой мышцы — в мотонейроне длинной малоберцовой мышцы регистрируется ВПСП, равный сумме отдельных ВПСП, получаемых при изолированном раздражении каждого волокна (рис. 49, Б}. Обычно требуется одновременная активация 50—100 афферентных волокон для получения ответа двигательной клетки спинного мозга. Временная суммация происходит при активации одного и того же афферентного пути серией последовательных раздражении. Если интервалы между поступающими импульсами достаточно коротки и ВПСП мотонейрона от предыдущего раздражения не успевают затухать, то последующие ВПСП накладываются друг на друга, пока деполяризация мембраны не Достигнет критического уровня для возникновения ПД (см. рис. 49, Л). Явления суммации могут наблюдаться при осуществлении отдельных рефлексов (например, рефлексов чихания и кашля в ответ на слабые раздражения слизистой оболочки дыхательных путей). Они лежат в основе формирования нейронных комплексов, определяющих поведение целого организма, в основе развития условных рефлексов. Фоновая активность. Многие нервные клетки обладают способностью к постоянной импульсной активности. Их мембранный потенциал периодически колеблется, то увеличиваясь, то уменьшаясь. Каждый раз, когда его увеличение превышает критический уровень деполяризации (порог возбудимости клетки), возникает импульсный разряд. Такая автоматическая деятельность называется фоновой активностью нейрона (рис. 50). Ее могут вызывать случайные импульсы, циркулирующие в нервной системе, но она может проявляться и при отсутствии приходящих к нейрону раздражении—в результате изменении ионных и обменных процессов, колебаний температуры и других причин. В последнем случае ее называют спонтанной.
Бездеятельные. Это облегчает протекание многих рефлекторных реакций. Во-вторых, клетки, обладающие постоянным ритмом фоновой активности, могут иметь значение водителя ритма (пейсмекера), т. е. задавать этот ритм другим нервным клеткам. При этом возникают большие комплексы синхронно работающих клеток, в которых благодаря общему ритму особенно хорошо выражено взаимодействие между отдельными нейронами. В-третьих, некоторые такие клетки выполняют роль счетчиков времени в центральной нервной системе. Их деятельность помогает поддерживать определенный ритм движений, субъективно оценивать интервалы времени, через определенные сроки включать необходимые функции и т. п. Автоматическая деятельность присуща клеткам дыхательного центра. Их спонтанные разряды позволяют поддерживать ритмические дыхательные движения не только при бодрствовании, но и во время сна человека, в условиях наркоза; периодические разряды дыхательного центра регистрируются даже при полной изоляции продолговатого мозга от других отделов центральной нервной системы.
Усвоение и трансформация ритма. При ритмических раздражениях вызванная активность нейрона может настроиться на ритм приходящих импульсов (явление усвоения ритма—рис. 51). Эта настройка деятельности нейрона на заданный извне ритм (реакция усвоения, перестройки или навязывания ритма) имеет большое значение для организации взаимодействия между различными нервными центрами и различными этажами нервной системы, в частности для организации ритмических движений у человека. Активность нейрона может перестроиться как на более высокий, так и на более низкий ритм. В результате такой способности многие нервные клетки получают возможность сонастраиваться, т. е. работать сообща, в одном комплексе, не мешая друг другу различной по темпу импульсацией. Впервые вопрос о значении усвоения ритма для протекания реакций организма был поставлен А. А. Ухтомским и его учениками (Н. В. Голиковым, Е. К. Жуковым и др.). Особенно важно, что в процессе ритмического раздражения рефлекторной дуги нервные центры способны усвоить ритм активности, превышающий их первоначальную лабильность. Это явление лежит в основе повышения работоспособности отдельных рефлекторных дуг, а также целостного организма (в частности, это имеет место в спортивной тренировке). Очень часто вызванная активность нейрона не соответствует ритму приходящей к данной клетке импульсации. Нервные клетки обладают свойством изменять частоту передающихся импульсов, т.е. свойством трансформации ритма. Характер ответного разряда зависит, во-первых, от свойств раздражителя и, во-вторых, от функционального состояния нейрона (его мембранного заряда, возбудимости, лабильности). В обычных условиях чем сильнее раздражение, тем чаще проявляется ответная импульсация. При высокой возбудимости нейрона может возникать учащение импульсации (мультипликация ритма). В этом случае афферентная волна вызывает длительные надпороговые ВПСП в клетке, и на.каждое одиночное раздражение клетка отвечает залпом из нескольких ПД (пока ВПСП превышает пороговый уровень). При низком уровне возбудимости и лабильности нейрона чаще происходит урежение ритма, так как несколько приходящих импульсов должны суммироваться, чтобы наконец достичь порога возникновения ПД. В результате на несколько приходящих импульсов нейрон отвечает лишь одним импульсом. В некоторых крупных клетках, например мотонейронах спинного мозга, имеются специальные механизмы, регулирующие ритм ответных реакций. После каждого ПД в этих клетках наблюдаются выраженные следовые процессы — следовая гиперполяризация мембраны. В этот момент возбудимость клетки понижена. Следующий разряд возникает в клетке лишь тогда, когда заканчивается этот следовой потенциал. Поэтому ритм разрядов мотонейрона, направляющихся к скелетной мышце, обычно не превышает 60 импульсов в 1 сек. Мелкие клетки, например вставочные нейроны, такого механизма не имеют и могут давать в начальные моменты ответа очень высокую частоту разрядов—до 1600 импульсов в 1 сек., которая, однако, быстро затухает. Следовые процессы. После окончания действия раздражителя активное состояние нервной клетки или нервного центра обычно продолжается еще некоторое время. Длительность следовых процессов различна: небольшая в спинном мозгу (несколько секунд или минут), значительно больше в центрах головного мозга (десятки минут, часы и даже дни) и очень велика в коре больших полушарий (до нескольких десятков лет). Различают явные и скрытые следы прежних раздражении. В явной форме сохраняются лишь кратковременные следовые процессы или последствия. Они могут быть одноименными — продолжение возникшего в клетке состояния (тормозное или экзальтационное последействие) или контрастными (возбуждение вслед за торможением, торможение вслед за возбуждением). Поддерживать состояние возбуждения или торможения в нервном центре могут импульсы, блуждающие в нервных цепях—задержавшиеся на длинных путях передачи или возвращающиеся повторно к нейрону по замкнутым цепям нейронов. Значительно сложнее по природе скрытые следы, характеризующиеся отсутствием импульсации в нервных центрах. Предполагают, что длительное сохранение в нервной клетке скрытых следов со всеми характерными свойствами раздражителя основано на изменении структуры составляющих клетку белков (возможно, также и белков глиальных клеток). Согласно биохимической теории памяти (X. Хиден), в процессе запоминания происходят структурные изменения в молекулах рибонуклеиновой кислоты (РНК), от которой зависит образование белков клетки. В результате этого в нервной клетке начинают синтезироваться «измененные» белки с отпечатками прежних раздражении, составляющие биохимическую основу памяти. Эксперименты на крысах и других животных показали, что способность их запоминать следы прежних раздражении или способность к обучению коррелирует с изменением содержания рибонуклеиновой кислоты в нервной ткани. Интересные опыты были проведены на червях. У них вырабатывали условные рефлексы — включение света сопровождалось электрическим ударом, вызывающим у червя реакцию сжимания. После ряда сочетаний этих раздражении одно включение света заставляло червей сжиматься. Эту способность реагировать на включение света двигательной реакцией приобретали также и «необученные» черви после того, как их кормили кашицей из растолченных «обученных» червей или вводили им рибонуклеиновую кислоту, полученную из тканей «обученных» червей. Следовательно, приобретенный опыт может передаваться от одного организма к другому с химическими веществами. При формировании новых двигательных навыков увеличивается содержание измененной рибонуклеиновой кислоты в соответствующих нервных центрах. Такие изменения, сохраняющиеся в двигательных нейронах, могут составлять основу так называемой «мышечной памяти». Известно, что человек, забывший в результате ранения одного из участков мозга начертания букв алфавита, может писать слова автоматически, пользуясь сохранившейся моторной памятью. Мышечная память позволяет человеку вспомнить заученные движения, которые он забыл после длительного перерыва. Непродолжительные последействия (длительностью до 1 часа) лежат в основе так называемой кратковременной памяти, а длительные следы, связанные с биохимическими перестройками в клетках,—в основе долгосрочной памяти. Кратковременная память служит человеку недолго (например, спросив по телефону справочной необходимый номер телефона, можно набрать его и сразу забыть). Долгосрочная память обеспечивает длительное сохранение следов. Она имеет важнейшее значение в обучении человека (например, для усвоения определенного объема теоретических знаний, фиксирования в мозгу выработанных двигательных навыков). Дата добавления: 2015-02-02 | Просмотры: 1309 | Нарушение авторских прав |
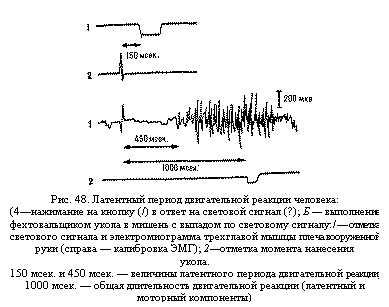 волокнам от периферических рецепторов в нервные центры и от нервных центров к исполнительным органам занимает сравнительно небольшое время, общее время от момента нанесения внешнего раздражения до появления ответной реакции организма (латентный период рефлекса) определяется центральным временем проведения. Величина латентного периода рефлекса служит важным показателем функционального состояния нервных центров и широко используется в практике. Латентный период двигательной реакции измеряется от момента подачи сигнала до момента начала движения (или до появления электрической активности соответствующей мышцы). При осуществлении человеком простых движений в ответ на внешние сигналы, например нажимание на кнопку при световом сигнале (рис. 48, А), длительность латентного периода двигательной реакции составляет около 120 – 220 мсек., а при утомлении, недостатке кислорода и пр. она может увеличиваться до 300 – 500 и более миллисекунд. При более сложных движениях, например выполнении фехтовальщиком укола в мишень с выпадом в ответ на световой сигнал, этот период значительно больше – около 300 – 400 мсек. и более (см. рис. 48, Б).
волокнам от периферических рецепторов в нервные центры и от нервных центров к исполнительным органам занимает сравнительно небольшое время, общее время от момента нанесения внешнего раздражения до появления ответной реакции организма (латентный период рефлекса) определяется центральным временем проведения. Величина латентного периода рефлекса служит важным показателем функционального состояния нервных центров и широко используется в практике. Латентный период двигательной реакции измеряется от момента подачи сигнала до момента начала движения (или до появления электрической активности соответствующей мышцы). При осуществлении человеком простых движений в ответ на внешние сигналы, например нажимание на кнопку при световом сигнале (рис. 48, А), длительность латентного периода двигательной реакции составляет около 120 – 220 мсек., а при утомлении, недостатке кислорода и пр. она может увеличиваться до 300 – 500 и более миллисекунд. При более сложных движениях, например выполнении фехтовальщиком укола в мишень с выпадом в ответ на световой сигнал, этот период значительно больше – около 300 – 400 мсек. и более (см. рис. 48, Б). Укорочение латентного периода одной и той же двигательной реакции человека в процессе спортивной тренировки может отражать повышение функционального состояния нервных центров (ускорение проведения) и перестройку нервных цепей, участвующих в передаче нервных импульсов (укорочение проводящего пути).
Укорочение латентного периода одной и той же двигательной реакции человека в процессе спортивной тренировки может отражать повышение функционального состояния нервных центров (ускорение проведения) и перестройку нервных цепей, участвующих в передаче нервных импульсов (укорочение проводящего пути). Фоновая активность играет важную роль в деятельности целого организма. Во-первых, импульсы, приходящие по афферентным путям, легче возбуждают клетки со спонтанной активностью, чем
Фоновая активность играет важную роль в деятельности целого организма. Во-первых, импульсы, приходящие по афферентным путям, легче возбуждают клетки со спонтанной активностью, чем Вызванная активность. Импульсные разряды нейрона, возникающие в ответ на внешнее раздражение, называются вызванной активностью. Ответ нервной клетки может возникнуть в форме одиночного ПД, серии импульсов с затухающей частотой, а также в виде пачек импульсов, появляющихся через определенные интервалы. В различных нейронах наблюдаются разные типы вызванной активности. Одни клетки дают разряды только в момент включения раздражителя (эффект включения) —это наиболее частая реакция клеток, другие — только в момент его выключения (эффект выключения), третьи—в обоих случаях. В центральной нервной системе имеются клетки, дающие разряды лишь на новые раздражители, что обеспечивает ориентировочную реакцию. Известны также клетки, в которых отмечается вызванная активность при неправильных реакциях организма, т. е. сигнализирующие об ошибках, что позволяет вносить коррекции в действия человека.
Вызванная активность. Импульсные разряды нейрона, возникающие в ответ на внешнее раздражение, называются вызванной активностью. Ответ нервной клетки может возникнуть в форме одиночного ПД, серии импульсов с затухающей частотой, а также в виде пачек импульсов, появляющихся через определенные интервалы. В различных нейронах наблюдаются разные типы вызванной активности. Одни клетки дают разряды только в момент включения раздражителя (эффект включения) —это наиболее частая реакция клеток, другие — только в момент его выключения (эффект выключения), третьи—в обоих случаях. В центральной нервной системе имеются клетки, дающие разряды лишь на новые раздражители, что обеспечивает ориентировочную реакцию. Известны также клетки, в которых отмечается вызванная активность при неправильных реакциях организма, т. е. сигнализирующие об ошибках, что позволяет вносить коррекции в действия человека.