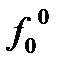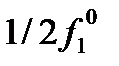|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Закон Харди-Вайнберга для полиплоидов
В популяциях автотетраплоидов (сорта ржи, гречихи, кормовых трав) значительная часть генотипов является гетерозиготами (симплексы, дуплексы, триплексы).При p=q=0,5 доля гомозигот (нулиплексы, квадриплексы) составляет незначительную часть популяции. У диплоидов же при p=q=0,5 половина особей популяции – гомозиготыРавновесие у полиплоидов не может быть достигнуто за одно поколение размножения, поскольку у полиплоидов возрастает число генотипов гамет и для достижения равновесия требуется нескольких поколений размножения.Равновесные доли пяти генотипов в популяции автотетраплоидов задаются выражением: (pA+qa)4=p 4A4+4p3 q A3 a+6p2 q2 A2 a2+4Pq3+q4 a4 7 Закон стабилизирующего скрещивания К. Пирсона (1904 г): Если популяция не находится в состоянии равновесия, то равновесие при случайном скрещивании достигается через одно поколение и не зависит от первоначального состава популяции. Все популяции с одними и теми же частотами генов (хотя частоты генотипов могут и различаться) при случайном подборе пар придут к одному и тому же равновесному состоянию. p=0,5*f1+f2; q=1-p; f'2=p2; f'1=2pq,,, f'0=q2….f2=4f2f0
8 Для определения частот аллелей при неполном доминировании или кодоминировании, т.е. когда все генотипы можно распознать по фенотипу используют формулы максимального правдоподобия Р.Фишера:p=0,5f1+f2 q=0,5f1+f0 p=0.5n1 +n2\Nобщ Ожидаемую частоту аллелей можно оценить следующим образом (при кодоминировании):p=N11+1\2N12\N q=1\2N12+N22\NВ малых выборках расчёты могут не соответствовать в точности частотам аллелей в реальной популяции. Чем больше размер выборки, тем точнее оценки частот аллелей в популяции. О точности таких оценок можно судить по величине вариансы. Варианса частоты рецессивного аллеля: V(q)=q(1-q)\2N Стандартная ошибка средней величины оценок представляет собой корень квадратный из вариансы, потому что оцениваемая частота аллелей сама по себе усреднённая величина. При достаточно больших размерах выборки доверительный интервал с надёжностью в 95% примерно вдвое больше или меньше стандартной ошибки средней величины. Эти величины и варианса дают максимально вероятные оценки параметров.Например, в популяции выявлено 100 особей с красной окраской лепестков, 40 особей с розовой окраской лепестков и 60 с белой.Частоты генотипов составят: p=N11\N=100\200=0,5, H=N12\N, Q=N22\NЧастоты аллелей и вариансы составят: p=N11+1\2N12\N=P+0,5H V(p)=v(q0=q(1-q)\2NРавновесные (теоретические) частоты генотипов составят: P'=p2 =0,62 =0,36 H'=2pq Q'=q2 9 При оценке частоты аллелей предполагается, что у гетерозигот эксперссируются все эти аллели, или они кодоминантны, т.е. в потомстве F1 проявляются признаки обоих родителей. Однако во многих случаях, например при болезнях, связанных с нарушением обмена веществ у человека или при полиморфизмах окраски у разных организмов, гетерозиготы обычно неотличимы от гомозигот. В таких случаях для оценки частот аллелей вводится допущение, что в популяции сохраняются соотношения аллелей, соответствующие закону Харди-Вайнберга. Частота рецессивного аллеля и соответствующая варианса при доминировании рассчитываются по формулам: q=(N22\N)1\2; V(q)=1-q2 \2N=q(1-q)\2N+(1-q)2\4N Первое слагаемое в формуле вариансы равно биномиальной вариансе при кодоминантном наследовании двух аллелей. Поэтому в случаях, когда гетерозиготы неотличимы по фенотипу, варианса увеличивается на величину, равную второму слагаемому. Очевидно, что это слагаемое наибольшее тогда, когда частота аллеля А2 мала, а при 10 Для определение частоты носителей рецессивных признаков (гетерозигот) среди нормальных особей (доминантных гомозигот и гетерозигот) используют следующую формулуH=2pq\p2+2pq=2pq\p2+pq+pq=2pq\p(p+q+q)=2q\1+q 11 Для установления соответствия между фактическими частотами генотипов и теоретически ожидаемыми по закону Харди-Вайнберга проводят вычисления в следующем порядке:1.Определяют частоту аллелей по формуле Р.Фишера.2. Рассчитывают теоретические частоты генотипов по формуле Харди-Вайнберга.3. Определяют теоретические численности генотипов.4. Рассчитывают критерий (хи-квадрат). 5. Находят табличное значение критерия хи-квадрат.Если расчетная величина хи-квадрат меньше табличной, то частоты генотипов в изучаемой популяции соответствуют равновесию Харди-Вайнберга. Критерий соответствия (хи-квадрат) рассчитывают по следующей формуле X2= ∑ki-1(O-E)2\E где О и Е – наблюдаемые и теоретически ожидаемые количества генотипов определённого типа, а k – число генотипических (фенотипических) классов. Табличное значение хи-квадрат находят по числу степеней свободы и уровню значимости в специальной таблице.Степень свободы определяет значимость величины хи-квадрат и равна числу фенотипических (генотипических) классов k минус единица и минус число оцениваемых параметров (число независимых величин).При кодоминировании число фенотипических классов, число оцениваемых параметров и число степеней свободы определяют по формулам:k=n(n+1)/2 df=n(n-1)/2 где n - число алалелий Число фенотипических классов при доминировании равно n, число оцениваемых параметров такое же, что и при кодоминировании (n-1), а число степеней свободы равно нулю. Уровень значимости обычно принимается равным 0,05 д.е. или 5%. Это означает, что гипотезу решено считать не соответствующей наблюдениям, если вероятность того, что расхождение между теоретически ожидавшимися и наблюдаемыми в эксперименте данными, обусловленное только случайными причинами, составляет не более 5%. Генное равновесие можно проверить и с помощью критерия соответствия К. Пирсона (1900) - x(хи-квадрат):x=∑ (O-E)2/E где О и Е – наблюдаемые и ожидаемые количества генотипов определённого типа, а k – число генотипических классов. При этом сопоставляют фактическое распределение членов популяции по фенотипам с теоретически ожидаемым. Достоверное расхождение между ними свидетельствует о нарушении генного равновесия в популяции.Например, в стадах каракульских овец, по данным Б.Н. Васина, встречаются овцы трёх типов, различающихся по форме ушной раковины: нормальная ушная раковина (гомозиготные доминанты АА), овцы с уменьшенным размером раковины (гетерозиготы Аа), безухие овцы (гомозиготные рецессивы аа).Сопоставление фактически полученных с теоретически ожидаемым числом овец вычисляют с помощью показателя хи-квадрат, по формуле:.Расчёт даёт следующее:
Суммируя полученные данные по каждому генотипу, получают величину хи-квадрат (0,268+1,698+2,74=4,71). Число степеней свободы (ню)для данного примера составляет: число генотип минус число аллелей, то есть ню=3-2=1Из таблицы значений хи-квадрата находят его теоретическую величину. При ню=1хи-квадрат равен 10,8.Так как хи-квадрат фактический (4,71) меньше табличного (10,8), то можно сделать вывод, что в данной популяции достоверно сохраняется генное равновесие в локусе, обусловливающем форму ушной раковины овец.Таким образом, генетическая структура популяции характеризуется для каждого конкретного локуса определёнными частотами генотипов и аллелей, а также наличием или отсутствием генного равновесия. Для определения генетической структуры популяции и генного равновесия пользуются формулой Харди-Вайнберга и методом хи-квадрат. 12 Признаки, сцепленные с полом, гены гапло-диплоидов. Знание особенностей генетических процессов происходящих в раздельнополых популяциях важно для селекции двудомных раздельнополых растений и для медицины.Например, ряд опасных болезней человека сцеплены с Х-хромосомой. К рецессивным признакам сцепленным с Х-хромосомой относятся: гемофилия, дальтонизм (неспособность различать красный и зелёный цвета), атрофия зрительного нерва и миопатия Дюшена. К доминантным признакам относят - рахит, не поддающийся лечению витамином Д, и тёмная эмаль зубов.При гетерогаметности мужского пола (ХУ) самцы гаплоидны (гемизиготны) по аллелям Х-хромосомы, поэтому гетерозиготны по этим аллелям только самки (ХХ).Для гапло-диплоидов, например Перепончатокрылых, где самцы гаплоидны, а самки диплоидны, гетерозиготность по всем локусам относится только к самкам. 13 Установление равновесного состояния в растительных популяциях при сцеплении с полом Система Х и У хромосом позволяет поддерживать равенство полов в популяции. При детерминации пола цветков у растений с Х и У хромосомами, растения с пестичными цветками, как правило гомогаметны (ХХ), а с тычиночными гетерогаметны (ХУ). В популяции раздельнополых растений (конопля, спаржа, шпинат и др.) скрещиваются растения противоположных фенотипов (диссортативное скрещивание).У растений с тычиночными цветками (ХУ) частоты аллелей и генотипов совпадают, если рассматриваемые гены локализованы в Х хромосоме (аллели генов находятся в гемизиготном состоянии):pA+qa=1Для растений с пестичными цветками (ХХ) частоты генов и генотипов записываются как обычно (каждый ген в Х хромосоме имеет по два аллеля): pA+qa=1; p2 AA+2pqAa+q2 aa=1При случайном переносе пыльцы и случайном оплодотворении частоты генов и генотипов находятся в равновесии, т.е. закон Харди-Вайнберга выполняется и для генов, сцепленных с половыми хромосомами.Однако, в случае сцепления с полом пестичные растения несут две дозы генов, а тычиночные – одну, поэтому равновесие частот генов и генотипов достигается только после нескольких поколений свободного размножения.Приближение к равновесию носит колебательный характер. Это связано с тем, что тычиночные растения получают Х хромосому от пестичного растения, а пестичные – от обоих родительских растений.Равновесная частота аллеля (pe) зависит от исходных частот в мужской (pm0) и в женской (pf0 не равно pm0) частях популяции следующим образом:pe=2/3pf0+1/3pm0Изменение частоты доминантного аллеля в ряду поколений свободного скрещивания в женской и мужской частях популяции при сцеплении с полом
14 Теперь можно рассмотреть ситуацию, когда в мужской и женской частях популяции различаются частоты аллеля А по локусу, находящемуся в аутосоме (р2 и р1). После первого свободного скрещивания частоты генотипов АА, Аа и аа будут: (р1р2, р1q2 + p2q1, q1q2), что видно из данных таблицы.
Равновесия ещё нет. Равновесное состояние с частотой р=1/2(p1+p2)и частотами генотипов р2, 2рq, q2 наступает только после ещё одного скрещивания.Например скрещиваются мужские растения с р1=0,3 и женские с р2=0,6. В результате образуется популяция, у которой в мужской и женской частях структура по аутосомному гену одинакова [f2(AA)=p1p2=0,18; f1(Aa)=p1q2+p2q1=0,54;f0(aa)=q1q2=0,28Частота аллеля А в этой популяции: p'=f2+1/2f1=0,45=(0,3+0,6)/2 Равновесие по генотипической структуре ещё не достигнуто, поскольку равновесная доля гомозигот АА по закону Харди-Вайнберга должна составлять (p')2=0,2025, что не равно полученному f2 =0,18. После ещё одного скрещивания популяция переходит в равновесие с частотами генотипов в обеих частях [f2'=(0,45)2;f'1=2pq;f'0=q2 (А.В. Смиряев, 2003).Частоты аллелей у двух полов можно оценить, исходя из числа генотипов, как и для аутосомных локусов. Частота аллеля А2 составит у самок и самцов, соответственно:qf=1/2N12+N22/Nf, qm=N2/Nm,где N12, N22 и Nf – число особей с генотипом А1А2, А2А2 и общее число самок, соответственно, а N2 и Nm – число самцов с генотипом А2А2 и общее самцов в выборке (популяции). Например, можно оценить частоту аллелей Х -сцепленного гена черепаховой окраски у кошек. Окраску шерсти определяет ген yellow. Гетерозиготные по этому гену кошки имеют черепаховую окраску, или окраску набивного сица (calico). Гомозиготные по аллелю yellow кошки и гемизиготные самцы имеют жёлтую окраску. Данные по этому локусу, полученные в лондонских ветеринарных лечебницах, где усыпляли бродячих животных, представлены в таблице. В результате первого свободного скрещивания А//А х а/ → А//а + А/генетическая структура женской части популяции потомства определяется в равной степени частотами аллелей в обеих исходных частях:pf0=1/2p0f+1/2pm0=1/2*1+1/2*0=1/2; qf1=1-1/2=1/2; pm0=1; qm1=1-1=0 После первого свободного скрещивания:Таким образом, частоты генотипов АА, Аа и аа после первого свободного скрещивания будут следующие:p1p2;p1p2+p2q1,,q1q2Равновесие популяции ещё не достигнуто, что видно из примера:Равновесная частота доминантного аллеля равна f2=p2=0,452Однако эта частота не равна полученной при первом свободном скрещивании – 0,18. Следовательно популяция ещё не находится в равновесном состоянии. Равновесие наступит только после ещё одного свободного скрещивания. 15 Определение частот аллелей при сцеплении с полом При Х-сцепленном кодоминировании частота рецессивного аллеля определяется по следующим формулам:qf=0,5*N12+N22/Nf; qm=N2/NmВ случае Х-сцепленного доминирования частота рецессивного аллеля определяется по формулам:qf=корень N22/Nf qm=N2/NmПример для Х-сцепленного кодоминирования Рецессивный ген yellow - y, определяющий окраску шерсти у кошек сцеплен с Х-хромосомой. Гетерозиготные кошки (+y) имеют черепаховую окраску. Гомозиготные по аллелю y генотипы кошек (yy) и гемизиготные cамцы (y) имеют жёлтую окраску меха. Остальные кошки (++) и коты (+) имеют обычную окраску (дикий тип).Подсчёт животных в ветклинике дал следующие результаты:++ - 277 +y – 54y y – 7+ - 311y - 42 Из полученных данных можно определить частоты рецессивного аллеля у кошек и котов qf=0,5*54+7/277+54+7=34/338=0,101; q=42/311+42=42/353=0,119 16 Пример для Х-сцепленного доминирования У людей локус ma (макроглобулин а) содержит сцепленный с полом ген, кодирующий -макроглобулин сыворотки крови. Наличие в сыворотке крови этого антигена () доминантно по отношению к его отсутствию (). В одной из выборок среди населения Норвегии распределение фенотипов было следующим: у женщин – 57 и 44 у мужчин – 23 и 77 Следовательно, частота аллеля у женщин: Частота этого же аллеля у мужчин: qm=77/100=0,77 Частота аллеля в целом по всей популяции составляет: 17 Определение частоты встречаемости рецессивных фенотипов в мужской части популяции по сравнению с женской частью популяции Поскольку частоты двух гемизиготных генотипов в мужской части популяции при сцеплении с полом совпадают с частотами соответствующих аллелей у женских особей предыдущего поколения, то фенотипы, определяемые рецессивными генами, у мужских особей будут встречаться чаще, чем у женских.Если частота сцепленного с полом рецессивного аллеля равна, то частота определяемого им фенотипа будет равна для мужских особей и для женских особей. Отношение этих двух величин составляет: q/q2=1/qЧем меньше значение q, тем выше отношение частоты определяемого рецессивным геном фенотипа у мужских особей к его частоте у женских особей.Пример Частота рецессивного сцепленного с полом аллеля, вызывающего дальтонизм у людей (неспособность различать красный и зелёный цвета), составляет 0,08; следовательно, этот дефект встречается у мужчин в 1/0,08=12,5 раза чаще, чем у женщин. 18 В природе многие локусы содержат более двух аллелей. В случае множественного аллелизма равновесие по частотам всех диплоидных генотипов в популяции AiAj(I,j=1…n), так же, как и при двух аллелях, достигается уже после однократного случайного скрещивания.Для любого числа аллелей справедливо следующее правило: Частота аллеля гена при множественном аллелизме равна сумме частот гомозигот по этому аллелю и половины частот гетерозигот, несущих данный аллель.Если генотипAiAi встречается с частотой Pii,pi=0,5*(Pi1+Pi2+…+2Pii+…+Pij+…+Pin)=Pii+0,5∑Pijгде j Решётка частот объединения гамет имеет вид: Таким образом равновесные частоты генотипов в случае с тремя аллелями будут следующие: f'(A1 A1)=p2=0,0625; f'(A 1A2)=2p1 p2=0,15; f'(A 1A3)=0,225; f'(A2 A2)=0,09; f'(A2 A3)=0,27; f'(A3 A3)=0,2025
Как и для двух аллелей, частота генотипов и аллелей при множественном аллелизме оценивается при помощи выборок. При кодоминантности изучаемых в популяции аллелей методы вычисления частот генов и сопоставления фактического распределения фенотипов с теоретически ожидаемым по формуле Харди-Вайнберга проще.По формуле Харди-Вайнберга можно вычислить ожидаемую частоту и количество особей всех шести генотипов. Как видно из таблицы, ожидаемые значения очень близки к наблюдаемым. Ожидаемая гетерозиготность равна 0,594, а наблюдаемая в этой популяции – лишь немного ниже значения 0,561. 19 Определение частот электрофоретических аллелей (кодоминантные аллели)Метод гель-электрофореза ферментов позволяет оценить генетическую изменчивость популяций.А. Растворимые фракции, полученные из гомогенезированных образцов исследуемых тканей, наносят на поверхность геля и подвергают действию электрического поля.Ферменты и другие белки, содержащиеся в образцах, перемещаются и занимают определённые положения в геле.Б. После выключения электрического тока гель обрабатывают специальным химическим раствором для «проявления» пятен, соответствующих ферментам. Генотип особи по локусу, кодирующему данный фермент, определяют по характерному расположению пятен в геле.Гель, окрашенный после электрофореза для выявления фермента фосфоглюкомутазы
Были проанализированы данные по локусу малатдегидрогеназы (Mdh) у пресноводного рачка Daphnia magna. Закон Харди-Вайнберга применим и к большему числу аллелей. Это важно, поскольку известно, что многие локусы, особенно микросателлиты или гены, идентифицируемые по аллельной частоте, содержат более двух аллелей. Когда один аутосомный локус в популяции диплоидных организмов имеет не два, а более двух аллельных состояний: (А1, А2,..., Аn), n > 2, частоты этих аллелей обозначаются p1, p2,..., pn. В подобных случаях равновесие по частотам всех диплоидных генотипов вида AiAj (i,j=1...n), так же, как и при двух аллелях, достигается уже после однократного случайного скрещивания. В такой расширенной двухаллельной модели частота аллеля мультилокусного гена равна сумме частот гомозигот по этому аллелю и половины частот гетерозигот, несущих данный аллель. Это правило справедливо для любого числа аллелей. Закон Харди-Вайнберга справедлив для множественных аллелей при условиях случайности сочетаний гамет и скрещиваний. При соблюдении этих условий расширенные равновесные частоты для гомозиготных генотипов равны pi2 при i=j, а гетерозиготных – 2pipj при i Примеры трёхаллельных генетических систем (ST=стандарт, АR=аллель Arrowhead, CH=аллель Chiricahua, F=быстрый (Fast), M=средний (Medium), S=медленный (Slow), him.=гималайская (himalayan), ins.= аллель insularia, carb.= аллель carbonaria, typ.=аллель typical, null – отсутствие белкового продукта, обнаруживаемого при электрофорезе. 20 Наблюдаемая и ожидаемая гетерозиготность популяции Гетерозиготность популяции – один из показателей изменчивости (полиморфности) популяцииНаблюдаемая гетерозиготность оценивается по формуле: H0= ∑Nij /N где I не =jОжидаемая гетерозиготность (частота всех гетерозигот) определяется по формуле: HE =1- ∑i=1n pi2Ожидаемая гетерозиготность максимальна, когда аллели встречаются с одинаковой частотой, т.е.: pi=1/nПри равных долях всех аллелей ожидаемая гетерозиготность равна: HE =1- ∑(1/n)2 =n-1/nЧем больше аллелей в локусе, тем больше ожидаемая гетерозиготность.Нехватка гетерозигот говорит об отклонении от панмиксии, а избыток гетерозигот - об их селективном преимуществе. 21 Определение частот множественных аллелей при доминировании (наследование окраски крыльев у берёзовой пяденицы) Бабочка с тёмными крыльями (carbonaria) на лишайнике – слева;Бабочки со светлыми крыльями (typical) и с тёмными (carbonaria) на тёмном стволе - справа.Бабочки с промежуточной окраской не показаны (insularia). Три фенотипа бабочки контролируются тремя аллелями: p1 >p2 >p3Исходя из этого тёмную окраску крыльев (Carbonaria) будут иметь следующие генотипы:A1 A1, A1 A2, A1 A3Промежуточную окраску (insularia) будут иметь генотипы:A2 A2, A2 A3Светлую окраску крыльев (typical) будет иметь генотип – A3A3Частоты трёх рассматриваемых в данном примере аллелей определяются по формулам:p1=1-кореньN22+N23+N33/N=1-(p2+P3) p2=корен(ьN22+N23+N33/N)(N33/N)=корень(N22+N23+N33/N)-p3 ;p= корень N33/NРасчеты следует начинать с последней формулы 22 Определение частот аллелей при доминантно-кодоминантном типе наследования признаков Смешанное доминантно-кодоминантное наследование приводит к появлению заметного числа гомозигот по рецессивному аллелю (например, по локусу группы крови АВО) или же наоборот к снижению частоты рецессивного аллеля, так что гомозиготы по этому аллелю практически не встречаются (например, аллоферментные или микросателлитные локусы с двумя и более кодоминантными аллелями и одним нуль-аллелем, который не продуцирует белкового продукта, обнаруживаемого в электрофоретическом геле).Пример определения частот аллелей при доминантно-кодоминантном типе наследования:У человека аллели А и В локуса АВО кодоминантны и оба – доминируют над аллелем О. Фенотипу О соответствует генотип ОО (I группа крови), фенотипу А - генотипы АА и АО (II группа крови), фенотипу В – генотипы ВВ и ВО (III группа крови) и фенотипу АВ – генотип АВ (IV группа крови).Универсальные доноры – люди I группы крови, т.к. у них отсутствуют оба антигена (А и В), универсальные реципиенты – люди IV группы крови.Например, в выборке взятой из популяции африканских пигмеев, 7 человек имели фенотип АВ, 44 – А, 27 – В, 88 – О. p 1(A)=1- корень((N22 +N 23)+N33)/N); p2(B)=1-корень(N11 +N13)+N33/N); p3(O)=корень (N33\N); p 1(A)=p 2(B)=p 3(O) Смешанное доминантно-кодоминантное наследование приводит к появлению заметного числа гомозигот по рецессивному аллелю (например, по локусу группы крови АВ0) или же к снижению частоты рецессивного аллеля, так что гомозиготы по этому аллелю практически не встречаются (например, аллоферментные или микросателлитные локусы с двумя и более кодоминантными аллелями и одним нуль-аллелем, который не продуцирует белкового продукта, обнаруживаемого в электрофоретическом геле).У человека аллели А (IA) и В (IB) локуса АВ0 кодоминантны и оба – доминируют над аллелем 0 (H (I0)). Локус расположен на хромосоме 9. Продукты аллелей А и В являются химическими модификациями антигена H. Фенотипу А соответствуют генотипы АА и АО (I группа крови), фенотипу В – генотипы ВВ и ВО (II группа крови), фенотипу АВ – генотип АВ (III группа крови), фенотипу 0 – генотип 00 (IV группа крови). «Универсальные доноры», как известно, люди IV группы крови (генотип 00 (I0I0)), так как их эритроциты не содержат антигенов А и В, но в крови таких людей присутствуют антитела против этих антигенов. Лицам с группой крови АВ (III группа) можно переливать кровь любой группы, так как у них нет антител против антигенов А и В. Такие лица считаются «универсальными реципиентами». 23 Определение равновесных частот гамет и генотипов Если рассматривать каждый из двух генов (А-а, В-b) отдельно, то уже после первого случайного скрещивания популяция переходит в равновесие как по частотам генотипов АА, Аа, аа, так и по частотам ВВ, Вb, bb. Однако соотношение долей генотипов ААВВ, АаВВ, АаВb, …, aabb будет приближаться к равновесному постепенно в ряду поколений.Скорость этого приближения зависит:1) от начального соотношения частот генотипов;2) от степени сцепления между локусами А и В.
Исходя из этого для двух локусов частоты аллелей определяются по формулам: pA=g11+g10,pB=g11+g01 ,qa=1-pA=g01+g00,qb=1-pB=g10+g00
Постепенно в ряду поколений популяция приближается к равновесным частотам гамет, а частоты генотипов приближаются к произведениям равновесных частот гамет.Генотип ААВВ образуется в результате объединения гамет АВ и АВ. При случайном скрещивании вероятность объединения этих гамет в потомстве равна произведению их частот:g11g11
Оценка близости популяции к равновесию по двум локусам, показатель неравновесия по сцеплению Для оценки близости популяции к равновесию по двум локусам в конкретном поколении случайных скрещиваний используют показатель D, который называется неравновесием по сцеплению:D=g11g00-g10g01 -0,25<=D<=0,25Показатель неравновесия по сцеплению – это произведение частот гамет в фазе притяжения минус произведение частот гамет в фазе отталкивания. Данный показатель разработали Левонтин и Коджима для оценки отклонения от случайной ассоциации между аллелями в разных локусах. От поколения к поколению D приближается к нулю, а популяция к равновесию.В поколении с номером n показатель Dn зависит от двух параметров: 1. D0 - показатель неравновесия в исходной популяции Dn=(1-rf)n*D0 2) rf - частота рекомбинации между локусами А и В Dn=0,5n*D0Из этих формул следует, что популяция быстрее всего приближается к равновесию при максимальном значении rf=0,5, т.е. когда сцепления нет.Цифровой пример:Пусть в исходной панмиктической популяции содержится два генотипа в равных долях – ААВВ (0,5); ааbb (0,5). Сцепления генов нет (rf=0,5).Исходная популяция:0,5ААВВ + 0,5ааbb Влияние сцепления на скорость приближения популяции к равновесию Dn/D0=D1/D0=0,125/0,25=0,5 D2=0,52 *0,25 D2/D0=0,0625/0,25=0,25 D1=(1-0,25)1*0,25=0,1875 25 Инбридинг – скрещивание особей, находящихся между собой в более тесном родстве, чем это имеет место при скрещиваниях в панмиктической популяции.Обратным по смыслу является понятие – аутбридинг. Ни инбридинг, ни аутбридинг, сами по себе, не приводят к изменению частот аллелей, но оба приводят к перераспределению аллелей в генотипах. В инбредной популяции частота гомозигот повышена, а частота гетерозигот понижена относительно соотношений Харди-Вайнберга. К инбридингу приводят три основные причины:1) Ограничения в размерах популяции.2) Изменчивость в системе скрещиваний внутри популяции (например, тенденция к скрещиванию растений, находящихся в пространственной близости друг от друга).3) Различные приспособления в цветках, способствующие самооплодотворению (например, опыление в закрытых цветках – клейстогамия).Пространственное размещение растений и система приспособлений к самоопылению, действую совместно, даёт суммарный эффект инбридинга. В популяциях со смешанным типом размножения – панмиксией и инбридингом – различают два типа гомозигот по одному и тому же гену: 1)аутогомозиготы (или аутозиготы); 2)обычные гомозиготы. В диплоидной популяции аутогомозиготой называется особь, у которой оба гена одного локуса несут идентичные по структуре последовательности ДНК вследствие того, что они – копии одного и того же гена. У гомозигот, не являющихся аутозиготами, оба гена одного локуса идентичны по состоянию – копии двух разных по происхождению аллелей. Родственными скрещиваниями в популяциях растений считаются1)самооплодотворение (автогамия), 2)брат-сестринские скрещивания (скрещивание потомков одного или двух растений), 3)скрещивания типа родитель – потомок и т.п. 4)Показателем доли родственных скрещиваний в популяции со смешанным размножением служит коэффициент инбридинга растений F, а показателем доли случайных скрещиваний – коэффициент панмиксии (панмиктический индекс) P. 5)Оба коэффициента находятся в отношении:F+P=1В популяциях со смешанным типом размножения – панмиксией и инбридингом – различают два типа гомозигот по одному и тому же гену: 6)аутогомозиготы (или аутозиготы); 7)обычные гомозиготы. В диплоидной популяции аутогомозиготой называется особь, у которой оба гена одного локуса несут идентичные по структуре последовательности ДНК вследствие того, что они – копии одного и того же гена. У гомозигот, не являющихся аутозиготами, оба гена одного локуса идентичны по состоянию – копии двух разных по происхождению аллелей. Неслучайное в отношении фенотипа скрещивание происходит в популяциях, где скрещивающиеся особи находятся в большем или меньшем родстве друг к другу. Такие скрещивания называются инбридингом в отличие от случайного аутбридинга. Ни инбридинг, ни аутбридинг, сами по себе, не приводят к изменению частот аллелей, но оба приводят к перераспределению аллелей в генотипах. В инбредной популяции частота гомозигот повышена, а частота гетерозигот понижена относительно соотношений Харди-Вайнберга. При аутбридинге наоборот: частота гетерозигот повышена, а гомозигот - понижена по сравнению с пропорциями при случайном скрещивании. Значительное влияние перекрёстного оплодотворения на частоты генотипов наблюдается только тогда, когда особи предпочтительно скрещиваются с особями из другой популяции (или субпопуляции). Внутри популяции, не имеющей структуры, эффект перекрёстного оплодотворения (аутбридинга) не может быть велик. Можно отметить несколько общих закономерностей инбридинга в популяциях:1. Генотипические изменения, обусловленные инбридингом, проявляются во всех локусах генома. Генный поток влияет на все локусы, а отбор или мутация действуют только на единственный локус (или на тесно сцепленные с ним локусы).2. Влияние на частоты генотипов может быть совершенно незначительным, если система скрещивания изменится. Например, высокая частота гомозигот, являющаяся результатом самооплодотворения, может быть полностью уничтожена одним поколением случайного скрещивания.3. Инбридинг и генетический дрейф, в общем, сходно влияют на гетерозиготность, но результаты их действия различны. Инбридинг (в большой популяции) может привести к понижению гетерозиготности без изменения частот аллелей данного локуса в популяции. С другой стороны, генетический дрейф может вызвать изменение частоты аллелей, но не приведёт к понижению гетерозиготности в популяции. Только при рассмотрении усреднённого влияния генетического дрейфа на данный локус во многих популяциях, или на многие локусы в данной популяции, можно выявить зависимость уровня гетерозиготности от генетического дрейфа.
26 Изменение частот генотипов при полном самоопылении Наиболее крайним типом обнаруживаемого при половом размножении инбридинга является самооплодотворение.
При полном самооплодотворении популяция разделяется на серии линий, которые быстро становятся гомозиготными. Только при самооплодотворении существует три типа скрещивания, которые происходят в популяции с частотой, соответствующей относительным пропорциям генотипов:f21=f02+1/4*f10 f11=1/2*f10 f10=f00+1/4*f01 Типы скрещиваний и частоты генотипов потомства при полном самооплодотворении P1=f12+1/2f11=(f20+1/4f10)+1/4f01=p0 т.е. самооплодотворение не приводит к изменению частоты аллелейЧастота гетерозигот при самоопылении уменьшается в 2 раза за поколение и приближается к 0, а частота гомозигот – к частотам p и q аллелей, входящих в них. Ht=(1/2)t*H0; Pt=P0+1/2[1-(1/2)t]*H0; Rt=R0+1/2*[1-(1/2)t]*H0; H=f1, P= f2, R=f0 При регулярном самоопылении в большой популяции с исходным генотипическим составом f2(например, 0,25), f1 (0,5), f0 (0,25) доли гетерозигот и гомозигот в поколениях самоопыления изменяются в соответствии с формулами, представленными в таблице. Влияние самоопыления на динамику частот генотипов для одного локуса с двумя аллелями. При самоопылении частота гетерозигот уменьшается в 2 раза за поколение и приближается к 0, а частота гомозигот – к частотам p и q аллелей, входящих в них. Сами частоты аллелей не меняются в ряду поколений самоопыления. При любой схеме скрещивания с инбридингом долю гетерозигот в поколении t популяции удобно обозначить через 2pq (1 – Ft), где Ft называется коэффициентом инбридинга. Если Ft=0, то доля гетерозигот в поколениях неизменна и составляет 2pq (равновесие по закону Харди-Вайнберга). Если в поколении t коэффициент инбридинга достиг единицы, то гетерозигот в популяции больше не осталось (f1 =0).При инбридинге снижение доли гетерозигот на 2рqFt вызывает увеличение доли каждой из двух гомозигот на pqFt. 27 Прогнозирование генотипической структуры популяции для любого поколения самоопыления При инбридинге снижение доли гетерозигот на 2pqFt вызывает увеличение доли каждой из двух гомозигот на pqFt Поэтому структура популяции может быть представлена следующим образом:
Интерес представляет величина λ, свидетельствующая об уменьшении доли гетерозигот в двух соседних поколениях t и t + 1. λ=f1(t+1)/f(t)=2pq(1-Ft+1/2pq(1-Ft)=1-Ft+1/1-Ft или f1(t+1)=λ*f1(t) В зависимости от степени инбридинга значение λ может меняться от ½ до 1. При самоопылении λ=1/2 и доля гетерозигот максимально уменьшается в каждом последующем поколении. Из последней формулы можно вывести связь Ft с Ft+1 в следующем поколении самоопыления: Ft+1=1/2(1+Ft). Дата добавления: 2015-02-02 | Просмотры: 2564 | Нарушение авторских прав |
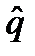 <1/3 первое слагаемое больше второго.В случае, когда гомозиготные и гетерозиготные доминантные генотипы фенотипически не различаются между собой, частота рецессивного аллеля определяется на основании частоты рецессивного генотипа, легко обнаруживающегося в популяции визуально.Пусть например доминантный аллель А определяет красную окраску лепестков, рецессивный а – белую. В равновесной панмиктической популяции на 10000 растений – только 25 с белыми цветками, т.е. доля рецессивных гомозигот аа в соответствии с законом Харди-Вайнберга равна q2 =25/10000=0,0025. Тогда доли аллелей в этой популяции равны q =0,05, p=1-q =0,95. Или q=(N22\N)1\2 = корень из 25\10000=0,05 V(q)= 1-q2\4N Частоты генотипов АА и Аа, которые по фенотипу не различаются, равны: p2 =(0,95)2=0,9025; 2pq =0,095, а частота генотипов аа равна q2 =0,0025. В случае болезней, связанных с нарушением обмена веществ у человека важно знать долю гетерозигот – носителей дефектного аллеля. Сейчас таких носителей легко идентифицировать с помощью молекулярных методов. Если это невозможно, то частота гетерозигот как «носителей» каких-либо рецессивных признаков может быть вычислена, если известна частота гена. Если равновесие Харди-Вайнберга справедливо, то частота гетерозигот среди всех особей, включая гомозиготы, равна H=2pq или 2q(1-q), где q- средняя из выражения q=(N22\N)1\2 и p=1-q. Частоты генов и генотипов для двух аллелей одного локуса при полном доминировании определяют по формулам Харди-Вайнберга q=корень из f0…..p=1-q……f1=2pq…..f2=p2 Если в задаче дана численность рецессивного генотипа среди всех особей, то сначала нужно определить частоту рецессивного генотипа по формуле:f0=n0\Nобщ
<1/3 первое слагаемое больше второго.В случае, когда гомозиготные и гетерозиготные доминантные генотипы фенотипически не различаются между собой, частота рецессивного аллеля определяется на основании частоты рецессивного генотипа, легко обнаруживающегося в популяции визуально.Пусть например доминантный аллель А определяет красную окраску лепестков, рецессивный а – белую. В равновесной панмиктической популяции на 10000 растений – только 25 с белыми цветками, т.е. доля рецессивных гомозигот аа в соответствии с законом Харди-Вайнберга равна q2 =25/10000=0,0025. Тогда доли аллелей в этой популяции равны q =0,05, p=1-q =0,95. Или q=(N22\N)1\2 = корень из 25\10000=0,05 V(q)= 1-q2\4N Частоты генотипов АА и Аа, которые по фенотипу не различаются, равны: p2 =(0,95)2=0,9025; 2pq =0,095, а частота генотипов аа равна q2 =0,0025. В случае болезней, связанных с нарушением обмена веществ у человека важно знать долю гетерозигот – носителей дефектного аллеля. Сейчас таких носителей легко идентифицировать с помощью молекулярных методов. Если это невозможно, то частота гетерозигот как «носителей» каких-либо рецессивных признаков может быть вычислена, если известна частота гена. Если равновесие Харди-Вайнберга справедливо, то частота гетерозигот среди всех особей, включая гомозиготы, равна H=2pq или 2q(1-q), где q- средняя из выражения q=(N22\N)1\2 и p=1-q. Частоты генов и генотипов для двух аллелей одного локуса при полном доминировании определяют по формулам Харди-Вайнберга q=корень из f0…..p=1-q……f1=2pq…..f2=p2 Если в задаче дана численность рецессивного генотипа среди всех особей, то сначала нужно определить частоту рецессивного генотипа по формуле:f0=n0\Nобщ
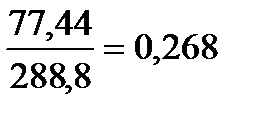


 р0♂) частях популяции следующим образом: р=
р0♂) частях популяции следующим образом: р=  р0♀+
р0♀+  р0♂.Это связано с тем, что ген, сцепленный с полом, в женской части популяции содержится в двойной дозе (ХХ), а в мужской – только в одной хромосоме Х. Например, популяция насекомых состоит из мужских и женских особей с контрастными частотами аллеля А: р0♀=1; р0♂=0. В результате первого свободного скрещивания А//А х а/→А//а + А/ генетическая структура женской части популяции потомства определяется в равной степени частотами аллелей в обеих исходных частях р♀=
р0♂.Это связано с тем, что ген, сцепленный с полом, в женской части популяции содержится в двойной дозе (ХХ), а в мужской – только в одной хромосоме Х. Например, популяция насекомых состоит из мужских и женских особей с контрастными частотами аллеля А: р0♀=1; р0♂=0. В результате первого свободного скрещивания А//А х а/→А//а + А/ генетическая структура женской части популяции потомства определяется в равной степени частотами аллелей в обеих исходных частях р♀=  р0♀ +
р0♀ +  р0♂=
р0♂=  и следующими равновесными частотами генотипов.
и следующими равновесными частотами генотипов. Гель содержит образцы тканей от 12 самок дрозофилы.Генотипы будут следующие:Pqm100/100 Pqm100/108 и тд.Пример определения частот электрофоретических аллелей (кодоминирование)
Гель содержит образцы тканей от 12 самок дрозофилы.Генотипы будут следующие:Pqm100/100 Pqm100/108 и тд.Пример определения частот электрофоретических аллелей (кодоминирование) j.Это можно показать с помощью решётки – таблицы частот встречаемости аллелей, например для случая n=3.Например, пусть частоты шести генотипов в исходной популяции равны: f(A1A1)=0,15; f(A1A2)=0,1; f(A1A3)=0,1; f(A2A2)=0,35. Тогда частоты гамет с тремя аллелями перед скрещиванием составят:p1(A1)=f(A1A1)+1/2f(A1A2)+1/2f(A1A3)=0,25;p2(A2)=f(A2A2)+1/2f(A1A2)+1/2f(A2A3)=0,30;p3(A3)=f(A3A3)+1/2f(A1A3)+1/2f(A2A3)=0,45.Если построить решётку частот объединения этих гамет, то можно установить, что равновесные частотыp, составят:f'(A1 A1)=p f'(A1 A2)=2p1*p2 Можно доказать, что максимум суммы равновесных долей всевозможных гетерозиготных генотипов достигается при равных долях всех аллелей, т.е. при pi=1/n. Этот максимум равен (n-1)/n и увеличивается, приближаясь к единице по мере возрастания n – числа аллелей в локусе, т.е. доля различных гетерозигот приближается к единице. Например, при n=5, pi = 0,2, а суммарная доля равновесных частот всех гетерозигот в популяции составит 0,8, а гомозигот только 0,2.
j.Это можно показать с помощью решётки – таблицы частот встречаемости аллелей, например для случая n=3.Например, пусть частоты шести генотипов в исходной популяции равны: f(A1A1)=0,15; f(A1A2)=0,1; f(A1A3)=0,1; f(A2A2)=0,35. Тогда частоты гамет с тремя аллелями перед скрещиванием составят:p1(A1)=f(A1A1)+1/2f(A1A2)+1/2f(A1A3)=0,25;p2(A2)=f(A2A2)+1/2f(A1A2)+1/2f(A2A3)=0,30;p3(A3)=f(A3A3)+1/2f(A1A3)+1/2f(A2A3)=0,45.Если построить решётку частот объединения этих гамет, то можно установить, что равновесные частотыp, составят:f'(A1 A1)=p f'(A1 A2)=2p1*p2 Можно доказать, что максимум суммы равновесных долей всевозможных гетерозиготных генотипов достигается при равных долях всех аллелей, т.е. при pi=1/n. Этот максимум равен (n-1)/n и увеличивается, приближаясь к единице по мере возрастания n – числа аллелей в локусе, т.е. доля различных гетерозигот приближается к единице. Например, при n=5, pi = 0,2, а суммарная доля равновесных частот всех гетерозигот в популяции составит 0,8, а гомозигот только 0,2.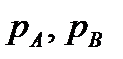 При полном сцеплении генов А и В, четыре комбинации аллелей АВ, Аb, аВ, ab можно рассматривать как четыре аллеля одного локуса. Равновесные частоты при не полном сцеплении зависят только от долей аллелей Обозначим частоты четырёх гамет, образующихся в гаметогенезе исходной популяции, следующим образом:
При полном сцеплении генов А и В, четыре комбинации аллелей АВ, Аb, аВ, ab можно рассматривать как четыре аллеля одного локуса. Равновесные частоты при не полном сцеплении зависят только от долей аллелей Обозначим частоты четырёх гамет, образующихся в гаметогенезе исходной популяции, следующим образом: fffff
fffff
 fff
fff