|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Тема 16. Проводящие пути ЦНС.В нервной системе импульсы, возникающие при воздействии на рецепторы, передаются по отросткам с нейрона на нейрон. Импульсы передаются только в одном направлении – от рецептора через вставочный нейрон к эфференту, что обусловлено морфофункциональными особенностями синапсов, которые проводят возбуждение только от пресинаптической мембраны к постсинаптической. Проводящие пути – это совокупность нервных волокон, проходящих в определенных зонах белого вещества головного и спинного мозга, объединенных общностью морфологического строения и функции. В спинном и головном мозге выделяют по строению и функции три группы проводящих путей.
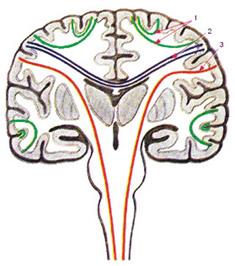
Рис. 1. Типы проводящих путей центральной нервной системы. 1 - ассоциативные проводящие пути, 2 - комиссуральные проводящие пути, 3 - проекционные проводящие пути.
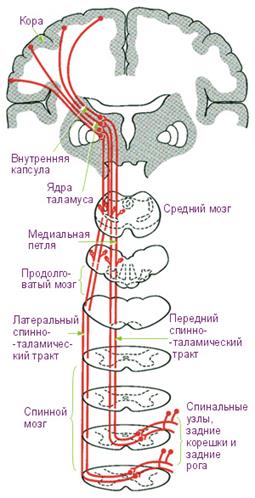
Ассоциативные пути соединяют участки серого вещества, различные функциональные центры (кора мозга, ядра) в пределах одной половины мозга. Выделяют короткие и длинные ассоциативные волокна. Короткие волокна соединяют близлежащие участки серого вещества и располагаются в пределах одной доли мозга – внутридолевые пучки волокон. Длинные ассоциативные волокна связывают участки серого вещества, расположенные на значительном расстоянии друг от друга, обычно в различных зонах. К ним относятся верхний продолговатый пучок, соединяющий кору лобной доли с теменной и затылочной, нижний продолговатый пучок, связывающий серое вещество височной доли с затылочной долей. В спинном мозге ассоциативные волокна связывают между собой нейроны, расположенные в различных сегментах. Они образуют собственные пучки спинного мозга (межсегментарные пучки), которые располагаются вблизи серого вещества. Короткие пучки перекидываются через 2-3 сегмента, а длинные пучки соединяют далеко расположенные сегменты спинного мозга. Комиссуральные (спаечные) нервные волокна связывают нервные центры правого и левого полушария большого мозга, образуют мозолистое тело, спайку свода и переднюю спайку, т.е. комиссуральные волокна проходят из одного полушария в другое. В мозолистом теле располагаются волокна, соединяющие новые, более молодые отделы мозга. В белом веществе полушарий волокна мозолистого тела расходятся веерообразно, образуя лучистость мозолистого тела. В спинном мозге комиссуральные пути образованы волокнами, переходящими с одной стороны спинного мозга на другую (волокна спиноталамического пучка и др.). Проекционные волокна соединяют нижележащие отделы с базальными ядрами и корой, и, наоборот, кору головного мозга, базальные ядра с ядрами мозгового ствола и со спинным мозгом. При помощи проекционных нервных волокон, достигающих кору большого мозга, картины внешнего мира как бы проецируются на кору, как на экран, где происходит высший анализ поступивших импульсов и сознательная их оценка. Выделяют проекционные восходящие и нисходящие пути. Восходящие (афферентные, чувствительные) несут импульсы, идущие от органов чувств, опорно-двигательного аппарата, внутренних органов и сосудов в головной мозг, к его подкорковым и высшим центрам. По характеру проводимых импульсов восходящие проекционные пути делят на 3 группы: 1) экстерорецептивные пути – импульсы поступают от органов чувств (зрения, слуха, вкуса, обоняния), кожных покровов (болевые, температурные, осязания, давления); 2) проприорецептивные пути – импульсы поступают от органов движения, несут информацию о положении частей тела, о размахе движений; 3) интерорецептивные пути – импульсы поступают от внутренних органов, сосудов (хемо-, баро-, механорецепторы). Экстерорецептивные проводящие пути. Все восходящие пути состоят из 3 нейронов: I нейроны начинаются в органах чувств и заканчиваются в спинном мозге или в стволовой части мозга, тела их располагаются в ганглиях. II нейроны располагаются в ядрах спинного или головного мозга и заканчиваются в ядрах таламуса, гипоталамуса. Эти нейроны образуют центростремительные восходящие пути. III нейроны лежат в ядрах промежуточного мозга, для кожной и мышечно-суставной чувствительности – в ядрах таламуса, для зрительных импульсов - в коленчатом теле, для обонятельных импульсов – в сосцевидных телах. Отростки нейронов заканчиваются на клетках соответствующих корковых центров (зрительной, слуховой, обонятельной и общей чувствительности). Проводящие пути болевой и температурной чувствительности образуют латеральный спинно-таламический путь. Рецепторы первого (чувствительного) нейрона пути болевой и температурной чувствительности, воспринимающего раздражения, располагаются в коже и на слизистой оболочке, а его тело лежит в спинномозговых узлах; центральный же отросток идет в составе заднего корешка в задний рог спинного мозга. Аксон второго нейрона, тело которого лежит в заднем роге, направляется на противоположную сторону спинного мозга. Через его переднюю серую спайку аксон входит в боковой канатик, где включается в состав латерального спинно-таламического пути, который поднимается до продолговатого мозга. Пучок располагается позади оливы, проходит в покрышку моста и покрышку среднего мозга. Аксоны заканчиваются, образуя синапсы на клетках, расположенных в таламусе (III нейрон). Аксоны III нейрона достигают коры полушария, его постцентральной извилины (IV слой коры), где находится корковый конец общей чувствительности. Импульсы от кожных рецепторов (рецепторы, воспринимающие чувство давления и осязания) поступают к клеткам коры в постцентральную извилину – место общей чувствительности.
Рис. 2. Передний и латеральный спинно-таламические пути.
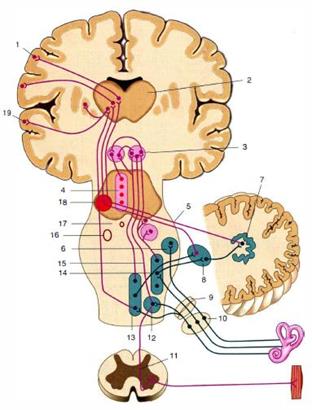
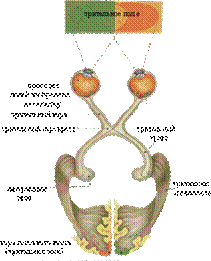
Проводящие пути специализированной чувствительности Вестибулярный (статокинетический) путь. Вместе с кожным, зрительным и двигательным анализаторами вестибулярный анализатор обеспечивает поддержание равновесия при разнообразных движениях и участвует в ориентировочных реакциях организма в пространстве (анализатор равновесия и гравитации). Путь начинается от рецепторов статокинетического анализатора, затем импульсы поступают к вестибулярному ганглию преддверно-улиткового нерва (1 нейроны); далее в составе преддверно-улиткового нерва они направляются к вестибулярным ядрам (2 нейроны); затем в составе бульбарно-таламического тракта проходят до таламуса, заканчиваясь на его центральных ядрах (3 нейроны). Аксоны центральных ядер таламуса направляются через заднюю ножку внутренней капсулы в корковую часть вестибулярного анализатора - кора полушарий средней и нижней височных извилин (4 нейроны). Следует обратить внимание, что аксоны вестибулярных ядер также формируют преддверно-спинномозговой путь ипреддверно-мозжечковый путь.
Рис. 3. Проводящие пути импульсов вестибулярной чувствительности. 1 - кора теменной доли мозга; 2 - таламус; 3 - медиальное ядро вестибулярного нерва; 4 - ядро глазодвигательного нерва; 5 - верхняя мозжечковая ножка; 6 - верхнее вестибулярное ядро; 7 - зубчатое ядро; 8 - ядро шатра; 9 - преддверная часть преддверно-улиткового нерва (VIII); 10 - вестибулярный узел; 11 - преддверно-спинномозговой путь (передний канатик спинного мозга); 12 - нижнее вестибулярное ядро; 13 - промежуточное и ядро медиального продольного пучка; 14 - латеральное вестибулярное ядро; 15 - медиальный продольный пучок; 16 - ядро отводящего нерва; 17 - клетки ретикулярной формации ствола мозга; 18 - красное ядро; 19 - кора височной доли мозга
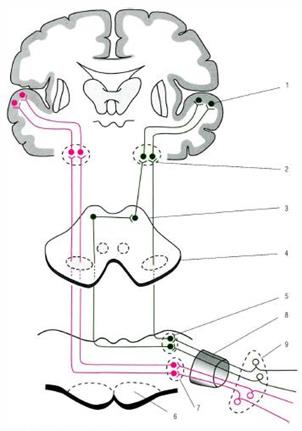
Слуховой путь обеспечивает проведение импульсов до подкорковых и корковых центров, участвуя в обеспечении ответных реакций на звуковые раздражения. От рецепторов (волосковые клетки Кортиева органа внутреннего уха) импульсы поступают к клеткам спирального ганглия(1 нейроны) преддверно-улиткового нерва, центральные отростки биполярных нейронов этого ганглияобразуют направляется к кохлеарным ядрампреддверно-улиткового нерва (2 нейроны). Аксоны вторых нейроноввыходят на дорсальную поверхность моста, образуя мозговые (слуховые) полоски четвертого желудочка, проходят в составе трапециевидного тела к его ядрампротивоположной стороны. Аксоны ядер трапециевидного тела (3 нейроны) в составе слуховой петли направляются к подкорковым центрам: нижние холмики среднего мозга, медиальные коленчатые тела, срединные ядра таламуса (4 нейроны). Далее: а) от нижних холмиков среднего мозга информация проводится к верхним холмикам, обеспечивая ответную реакцию на неожиданные слуховые раздражители; б) от срединных ядер таламуса импульсы поступают на медиальные ядра, которые являются подкорковым чувствительным центром экстрапирамидной системы; в) от ядер медиальных коленчатых тел слуховая информация проходит через заднюю ножку внутренней капсулы и затем направляется в среднюю часть верхней височной извилины - проекционный центр слуха (5 нейроны).
Рис. 4. Упрощенная схема проводящих путей слуховой сенсорной системы. 1 - верхняя височная извилина; 2 - медиальное коленчатое тело; 3 - нижний холмик пластинки крыши среднего мозга; 4 - латеральная петля; 5 - заднее ядро улиткового нерва; 6 - трапециевидное тело; 7 - переднее ядро улиткового нерва; 8 - улитковая часть преддверно-улиткового нерва; 9 - клетки спиралевидного узла.
Зрительный путь обеспечивает проведение световых импульсов от сетчатки до подкорковых и корковых центров, участвуя в обеспечении ответных реакций на зрительные раздражения. От палочек и колбочек, которые находятся в девятом слое сетчатки, импульсы поступают на периферические отростки биполярных клеток сетчатки (1 нейроны); центральные отростки биполярных клеток заканчиваются на ганглиозных нервных клетках сетчатки (2 нейроны), аксоны ганглиозных клеток формируют зрительный нерв, который направляется к зрительному перекресту, где перекрещиваются 2/3 нервных волокон, расположенных медиально (от внутренних отделов сетчатки), несущих информацию с латеральных сторон зрительного поля; неперекрещенная 1/3 волокон направляется в зрительный тракт своей стороны (от латеральных отделов сетчатки); она несет информацию от медиальных сторон зрительного поля; зрительный перекрест продолжается в зрительный тракт, который проводит импульсы к подкорковым центрам зрения (3 нейроны). От верхних холмиков импульсы поступают для обеспечения зрачкового рефлекса (изменение ширины зрачка при различном освещении); часть аксонов верхнего холмика направляется в интеграционный центр среднего мозга, который располагается также в верхнем холмике, гдеобеспечивается ответная реакция на неожиданные зрительные раздражители. От нейронов задних ядер таламуса импульсы поступают на медиальные ядра таламуса, которые являются подкорковым чувствительным центром экстрапирамидной системы, которые обеспечивают изменение тонуса мускулатуры в ответ на зрительные раздражения; От латеральных коленчатых тел зрительная информация проходит через заднюю ножку внутренней капсулы и затем в виде направляется к шпорной борозде, где располагается проекционный центр зрения (4 нейроны).
Рис. 5. Упрощенная схема зрительных проводящих путей.
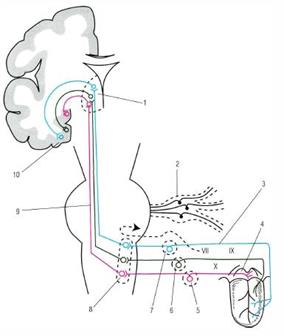
Вкусовой путь обеспечивает проведение импульсов от вкусовых сосочков до подкорковых и корковых центров, участвуя в обеспечении ответных реакций на вкусовые раздражения. От вкусовых клеток импульсы поступают к чувствительным ганглиям черепных нервов (1 нейроны)лицевого нерва от передних 2/3 языка; языкоглоточного нерва от задней 1/3 языка; блуждающего нерва от вкусовых рецепторов глотки, надгортанника, неба. Импульсы от указанных узлов следуют к ядру одиночного пути(2 нейроны). Аксоны клеток ядра одиночного пути переходят на противоположную сторону и следуют к ядрам таламуса (3 нейроны), аксоны третьих направляются в крючок и парагиппокампальную извилину - проекционный центр вкуса. Часть аксонов базальных ядер таламуса направляется в медиальные ядра таламуса - чувствительный интеграционный центр экстрапирамидной системы, который в ответ на вкусовые раздражения обеспечивает изменение тонуса мышц.
Рис. 6. Проводящие пути вкусового анализатора: 1 - таламус; 2 - ганглий тройничного нерва; 3 - промежуточный нерв; 4 - надгортанник; 5 - ганглий блуждающего нерва; 6 - ганглий языкоглоточного нерва; 7 - клетка узла коленца; 8 - вкусовое ядро (ядро одиночного пути); 9 - бульботаламический тракт; 10 - парагиппокампова извилина и крючок.
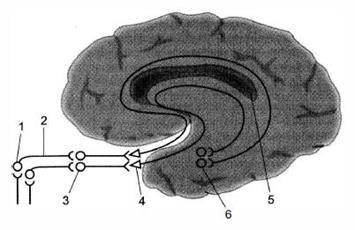
Обонятельный путь служит для проведения импульсов от обонятельных луковиц полости носа до подкорковых и корковых центров, участвуя в обеспечении ответных реакций на различные запахи. Рецепторами обоняния являются разветвления периферических отростков биполярных клеток (1 нейроны) слизистой оболочки полости носа. Центральные отростки рецепторных клеток образуют обонятельные нервы, которые проникают в полость черепа к митральным клеткам обонятельных луковиц(2 нейроны). Аксоны митральных клеток проходят в составе обонятельного трактаи вблизи обонятельного треугольника распадаются на три пучка: а) волокна медиального пучка направляются через переднюю спайку мозга в обонятельный тракт противоположной стороны к митральным клеткам обонятельной луковицы; б) волокна промежуточного пучка направляются к нейронам обонятельного треугольника, переднего продырявленного вещества и нейронам ядер прозрачной перегородки своей и, частично, противоположной сторон; в) волокна латерального пучка направляются к крючку и парагиппокампальной извилине - в проекционный центр обоняния. В подкорковые центры обоняния нервные импульсы поступают из проекционного центра обоняния. Важной особенностью обонятельного пути является то, что нервные импульсы первоначально поступают в кору полушарий большого мозга, и только затем - в подкорковые центры обоняния. Рис. 7. Схема проводящего пути обонятельного анализатора: 1 - обонятельные клетки; 2 - обонятельные нити; 3 - обонятельная луковица; 4 - обонятельный треугольник; 5 - мозолистое тело; 6 - клетки коры парагиппокампальной извилины
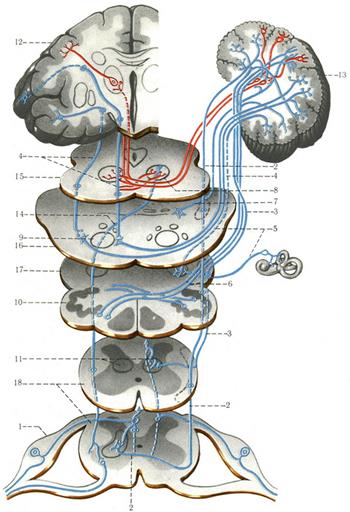
Проприорецептивные проводящие пути. Рецепторы I нейрона располагаются в мышцах, сухожилиях, связках, суставных капсулах. Тело I нейрона – в спинномозговом узле, их аксоны в составе заднего корешка, не входя в задний рог, направляются в задний канатик, а затем уходят в продолговатый мозг к тонкому и клиновидному ядрам, где заканчиваются синапсами на телах II нейронов. Аксоны II нейронов, выходящие из этих ядер, переходят на противоположную сторону, образуя медиальную петлю, проходят через покрышку моста и покрышку среднего мозга и заканчиваются в таламусе на телах третьих нейронов. Аксоны III нейронов направляются в кору постцентральной извилины, где заканчиваются в IVслое коры. Другая часть волокон III нейронов на выходе из тонкого и клиновидного ядер направляется в нижнюю мозжечковую ножку и заканчивается в коре червя. Третья часть волокон переходит на противоположную сторону, направляется через нижнюю мозжечковую ножку к коре червя противоположной стороны.
Рис. 8. Схема спинно-мозжечкового переднего и заднего, оливоспинномозгового, преддверно-мозжечкового, височно-мостового, мостомозжечкового, ретикуло-мозжечкового, мозжечково-красноядерного, красноядерно-спинномозгового, таламокоркового проводящих путей (по Сентаготаи). 1 — спинно-мозговой ганглий; 2 — передний спинно-мозжечковый путь; 3 — задний спинно-мозжечковый путь; 4 — кортико-мостовой и мосто-мозжечковый пути; 5 — вестибуломозжечковый путь; 6 — оливомозжечковый путь; 7 — ретикуломозжечковый путь; 8 — красное ядро; 9 — пирамидный путь; 10 — олива; 11 — n. dorsalis; 12 — парацентральная долька; 13 — мозжечок; 14 — руброспинальный путь; 15 — средний мозг; 16 — мост; 17 — продолговатый мозг; 18 — спинной мозг.
Кора (при участии сознания) управляет двигательными функциями организма непосредственно через пирамидные пути (произвольные движения). Пирамидным путь называется потому, что свое начало он берет от больших пирамидных клеток коры. Нисходящий пирамидный путь является эфферентным. Пирамидные пути имеют двухнейронное строение. Первые нейроны – это большие пирамидные клетки, расположенные в двигательной зоне коры. Вторые нейроны входят в состав двигательных ядер черепных нервов в стволе головного мозга и двигательных ядер передних рогов спинного мозга. Они называются периферическими мотонейронами. К пирамидным путям относятся главный двигательный, пирамидный, и корково-ядерный. Главный двигательный путь проводит двигательные волевые импульсы к мышцам туловища и конечностей. Начавшись от пирамидных клеток коры средней и верхней частей предцентральной извилины, волокна пирамидного пути идут в составе лучистого венца, а затем проходят через внутреннюю капсулу, занимая передние две трети ее задней ножки, причем волокна для верхней конечности идут спереди волокон для нижней конечности. Далее они проходят через ножку мозга, а оттуда через мост в продолговатый мозг и затем к двигательным ядрам передних рогов спинного мозга, а от них к скелетным мышцам. В зависимости от направления и расположения волокон пирамидный путь делится латеральный и передний корково-спинномозговой. Корково-ядерный путь начинается от клеток, залегающих в нижней трети предцентральной извилины, и проходит через колено внутренней капсулы. Волокна этого пути переходят на противоположную сторону, к двигательным ядрам черепных нервов: III и IV пар – в среднем мозге, V, VI, VII – в мосту, IХ, Х, ХI, ХII – в продолговатом мозге, где заканчиваются синапсами на их нейронах. Аксоны двигательных нейронов указанных ядер выходят из мозга в составе соответствующих черепных нервов и направляются к скелетным мышцам человека. Так как все двигательные волокна собраны на небольшом пространстве во внутренней капсуле (колено и передние две трети задней ножки ее), то при повреждении их в этом месте наблюдается односторонний паралич противоположной стороны тела. Экстрапирамидные проводящие пути образованы нисходящими проекционными нервными волокнами, по происхождению не относящимися к гигантским пирамидным клеткам коры больших полушарий мозга. Однако кора контролирует и управляет экстрапирамидной системой и является началом экстрапирамидных путей. Первичными центрами экстрапирамидной системы являются хвостатое и чечевицеобразное ядра полосатого тела, субталамическое ядро, красное ядро и черное вещество среднего мозга. Следующее звенья экстрапирамидных путей составляют ретикулярно-спинномозговой, красноядерно-спинномозговой, преддверно-спинномозговой и оливоспинномозговой пути, оканчивающиеся в передних столбах и промежуточном сером веществе спинного мозга. Экстрапирамидная система осуществляет непроизвольную регуляции и координацию движений, регуляцию мышечного тонуса, поддержание позы, организацию двигательных проявлений эмоций (смех, плач). Обеспечивает плавность движений, устанавливает исходную позу для их выполнения. При поражении экстрапирамидной системы нарушаются двигательные функции (например, могут возникнуть гиперкинезы, паркинсонизм), снижается мышечный тонус. Таким образом, проводящие пути головного и спинного мозга объединяют организм в одно целое, обеспечивают согласованность его действий.
Дата добавления: 2015-12-15 | Просмотры: 920 | Нарушение авторских прав |