|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Транспорт газов кровьюДЫХАТЕЛЬНАЯ СИСТЕМА
Дыхание – совокупность процессов, обеспечивающих потребление организмом кислорода и выделение двуокиси углерода. Основные функции системы: 1. Дыхательная – снабжение органов кислородом для обеспечения биологического окисления органических веществ, в результате которого выделяется энергия, нужная для жизни организма. 2. Выделительная – выделение углекислого газа, образующегося при окислении. 3. Гомеостатическая – поддерживает в норме физико-химические показатели крови и тканей. 4. Терморегуляторная – поддерживает температуру тела за счёт выведения паров воды. Дыхание включает в себя следующие процессы:
Внешнее дыхание
Вентиляция лёгких осуществляется в ходе дыхательного цикла: вдоха (инспирации) и выдоха (экспирации). В процессе вдоха и выдоха изменяется объём лёгких и это изменение связано с работой мышц. Мышцы, при сокращении которых объём увеличивается и совершается вдох, называют инспираторными, антагонисты – экспираторными. В норме вдох – это активный процесс, выдох – пассивный. При глубоком выдохе включаются в работу экспираторные мышцы, и он становится активным. К инспираторным мышцам относятся – диафрагма, наружные косые межрёберные мышцы (основные), лестничные, большая и малая грудные, передняя зубчатая (вспомогательные). К экспираторным мышцам – косые, поперечная и прямая мышцы живота, внутренние косые межрёберные. Лёгкие и стенки грудной полости покрыты серозной оболочкой – плеврой, между листками плевры располагается щель, которая заполнена серозной (плевральной) жидкостью. Механизм вдоха и выдоха. Во время сокращения инспираторных мышц объём грудной полости увеличивается, давление в щели становится еще более отрицательным (-6 мм.рт.ст.), лёгкие при этом расправляются, увеличивается их объём и воздух засасывается внутрь. Когда мышцы расслабляются, лёгкие в силу своей эластичности начинают спадаться – наступает выдох. Давление в плевральной щели отрицательное (-3 мм.рт.ст.), оно противостоит эластической тяге лёгких – упругим силам, которые направлены на спадение легких. Эластическая тяга лёгких обусловлена тремя факторами: 1. Поверхностным натяжением плёнки жидкости, покрывающей внутреннюю поверхность альвеол. Внутренняя поверхность альвеол покрыта специальной жидкостью – сурфактантом, в состав которой входят белки и липиды. Сурфактант при вдохе не даёт полностью спадаться одним альвеолам и перерастягиваться другим, вырабатывается пневмоцитами II типа. 2. Упругостью ткани стенок альвеол – содержат большое количество эластических волокон. 3. Тонусом бронхиальных мышц. Пневматоракс. В норме в плевральной щели газы отсутствуют, если же небольшое количество газов попадает в щель, то они быстро рассасываются, так как напряжение газов в крови лёгочных капилляров ниже, чем в атмосферном воздухе. Также в щели в норме не накапливается излишняя жидкость, так как онкотическое давление жидкости ниже онкотического давления плазмы крови. Если в щель попадает небольшое количество воздуха, то лёгкое частично спадается, но вентиляция его продолжается – это закрытый пневматоракс. Через некоторое время воздух всасывается и лёгкое расправляется. При ранениях лёгкое спадается полностью, его вентиляция прекращается – это открытый пневматоракс. Лёгочные объёмы. При спокойном дыхании человек вдыхает и выдыхает около 500 мл воздуха – это дыхательный объём. Резервный объём вдоха – количество воздуха, которое человек может вдохнуть после спокойного вдоха (3000 мл). Резервный объём выдоха – способен выдохнуть после спокойного выдоха (1300 мл). Жизненная ёмкость лёгких – количество воздуха, которое можно выдохнуть после максимального вдоха, равна сумме ДО+РОВ+РОВыд. У мужчин – 4,5 л, у женщин 3-4,5 л. В положении стоя выше. Остаточный объём – объём воздуха, который остаётся в лёгких после максимального выдоха (1200 мл). Минимальный объём – объём воздуха, находящийся в спавшихся лёгких. Этот воздух задерживается в «ловушках», которые образуется потому, что часть бронхиол спадается раньше альвеол. Общая ёмкость лёгких – максимальное количество воздуха, которое может находиться в лёгких: ОО+ЖЕЛ. Функциональная остаточная ёмкость – объём воздуха в конце спокойного выдоха: ОО+РОВыд. Минутный объём дыхания – объём вдыхаемого или выдыхаемого воздуха в единицу времени (6-9 л). ДО×ЧСС. Воздухоносные пути. Воздух находится не только в альвеолах, но и на протяжении всех воздухоносных путей. Этот воздух не участвует в газообмене. Поэтому просвет воздухоносных путей называют анатомическим мёртвым пространством. Значение воздухоносных путей: - увлажнение, согревание, очищение вдыхаемого воздуха; - осуществление защитных дыхательных рефлексов – при раздражении пылевыми частицами рецепторов носоглотки, гортани и трахеи возникает кашель, рецепторов полости носа – чихание. Центры кашля и чихания находятся в продолговатом мозге.
Лёгочное дыхание Газообмен между воздухом и кровью осуществляется в альвеолах. Альвеолы представляют собой полушаровидные выпячивания стенок альвеолярных ходов и дыхательных бронхиол. Большая часть наружной поверхности альвеол соприкасается с капиллярами малого круга кровообращения. Суммарная площадь этих контактов велика – около 90 м2. Вентиляция легких определяется объемом воздуха, вдыхаемого или выдыхаемого в единицу времени. Обычно ее оценивают через минутный объем дыхания (МОД). МОД=ЧД×ДО. Газообмен между воздухом и кровью осуществляется в альвеолах. Поэтому важна не общая величина вентиляции легких, а величина вентиляции именно альвеол – альвеолярная вентиляция (АВ). Она меньше вентиляции легких на величину вентиляции мертвого пространства. Объем анатомически мертвого пространства (МП) около 150 мл, или примерно ⅓ дыхательного объема при спокойном дыхании. Вентиляция мертвого пространства (ВМП) = 150×ЧД. АВ=МОД – ВМП. Эффективность вентиляции альвеол выше при более глубоком и редком дыхании, чем при частом и поверхностном. В последнем случае большая часть воздуха идёт на вентиляцию мёртвого пространства. В обычных условиях человек дышит атмосферным воздухом, имеющим относительно постоянный состав: 21% кислорода и 0,03% углекислого газа. В выдыхаемом воздухе всегда меньше кислорода (16 %) и больше углекислого газа (4,5%). В альвеолярном воздухе – 14 % кислорода и 5,5 % углекислого газа. Различие в составе альвеолярного и выдыхаемого воздуха объясняется тем, что последний является смесью воздуха мёртвого пространства и альвеолярного воздуха. Альвеолярный воздух является внутренней газовой средой организма. От его состава зависти состав артериальной крови. При обычном дыхании состав альвеолярного воздуха почти постоянен, при глубоком дыхании зависимость состава от фаз вдоха и выдоха увеличивается. Постоянство состава альвеолярного газа обеспечивается регуляцией дыхания и является необходимым условием нормального протекания газообмена. Воздух, заполняющий мертвое пространство, играет роль буфера, который сглаживает колебания состава альвеолярного газа в ходе дыхательного цикла. Различают два способа перемещения молекул газов в воздухоносных путях: - конвективный – обусловлен движением всей смеси газов по градиенту общего давления; - диффузионный – это перемещение отдельных газов вследствие разности его парциального давления. Газообмен в альвеолах осуществляется в результате диффузии кислорода из альвеолярного воздуха в кровь (ок. 500 л/сутки) и двуокиси углерода из крови в альвеолярный воздух (ок. 430 л/сутки). В крови газы находятся в растворённом и химически связанном состоянии. В диффузии участвуют только молекулы растворённого газа. Количество газа, растворяющегося в жидкости, зависит от: - состава жидкости; - объёма и давления газа над жидкостью; - температуры жидкости; - природы газа. Чем выше давление газа и чем ниже температура жидкости, тем больше молекул газа в ней растворяется. Растворение продолжается до наступления динамического равновесия между растворяющимися и выходящими в газовую среду молекулами. Сила, с которой молекулы растворённого газа стремятся выйти в газовую среду, называется напряжением газа в жидкости. Таким образом, в состоянии равновесия напряжение газа в жидкости равно парциальному давлению газа над жидкостью. Если парциальное давление газа выше его напряжения, то газ будет растворяться. И, наоборот. Проницаемость лёгочной мембраны для газа выражают величиной диффузионной способности лёгких. Это – количество газа, проникающего через лёгочную мембрану за 1 мин на 1 мм.рт.ст. градиента давлений. Пропорциональна толщине мембраны. Для кислорода ДС в норме составляет 25 мл/мин*мм.рт.ст., для углекислого газа – в 24 раза выше.
Транспорт газов кровью
Переносчиком дыхательных газов является кровь. В свободном (растворённом) виде переносится лишь очень небольшое количество газов. Основное количество газов переносится в связанном виде.
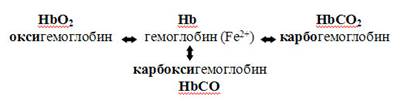
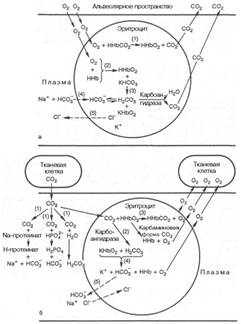
Транспорт кислорода. В растворенном виде переносится всего около 2 % кислорода. Основное его количество транспортируется в виде оксигемоглобина. Превращение гемоглобина в оксигемоглобин определяется напряжением растворённого кислорода. Графически эта зависимость выражается кривой диссоциации оксигемоглобина. Когда напряжение кислорода равно нулю, в крови находится только восстановленный гемоглобин (дезоксигемоглобин). Повышение напряжения кислорода сопровождается увеличением количества оксигемоглобина. Но данная зависимость существенно отличается от линейной. При 60 мм.рт.ст. насыщение гемоглобина кислородом достигает 90%, а при дальнейшем повышении напряжения кислорода приближается к полному насыщению очень медленно. Сродство гемоглобина к кислороду непостоянно и зависит от следующих факторов: - концентрации ионов водорода и углекислого газа; - концентрации 2,3-дифосфоглицерата (его количество увеличивается при снижении напряжения кислорода в крови, глицерат связывается с гемоглобином и снижает его способность связываться с кислородом, оксигемоглобин при этом отдает кислород); - температуры. Увеличение концентрации данных веществ и повышение температуры приводят к смещению кривой диссоциации вправо – это указывает на снижение сродства гемоглобина к кислороду. Оксигемоглобин легче диссоциирует (разлагается) на гемоглобин и кислород, кислород переходит в ткани. Такое положение кривой диссоциации характерно для тканей с интенсивным обменом веществ (в них концентрация углекислого газа и кислых продуктов увеличивается, а температура повышается). В волокнах скелетной мускулатуры содержится близкий к гемоглобину белок миоглобин. Он обладает очень высоким сродством к кислороду. Максимальное количество кислорода, которое может связать кровь при полном насыщении гемоглобина кислородом, называется кислородной ёмкостью кро ви. Кислородная ёмкость крови зависти от содержания в ней гемоглобина. В 1 л артериальной крови содержится около 180-200 мл кислорода, в венозной – 120 мл. Таким образом, протекая по тканевым капиллярам кровь, отдаёт не весь кислород. Часть кислорода, поглощаемая тканями из артериальной тканями из артериальной крови, называется коэффициентом утилизации кислорода. В покое он колеблется от 30 до 40%, при тяжёлой мышечной работе он повышается до 50-60%. Транспорт углекислого газа. Углекислый газ переносится кровью в трёх формах: - растворённой; - в виде кислых солей угольной кислоты (больший объём); - карбгемоглобина. При чём, образование солей угольной кислоты и карбгемоглобина идёт внутри эритроцитов. В эритроцитах углекислый газ соединяется с водой, образуя угольную кислоту. Этот процесс катализируется ферментов карбоангидразой. Она присутствует только в эритроцитах. Поэтому гидратация двуокиси происходит практически только в эритроцитах. В зависимости от напряжения углекислого газа карбоангидраза катализирует как образование угольной кислоты, так и расщепление ее на СО2 и воду (в капиллярах лёгких) Часть молекул двуокиси углерода соединяется в эритроцитах с гемоглобином, образуя карбгемоглобин. Благодаря связыванию напряжение двуокиси углерода в эритроцитах оказывается невысоким. Поэтому всё новые количества УГ диффундируют внутрь эритроцитов. В тканях происходит разрушение оксигемоглобина и образование карбгемоглобина. В лёгких, наоборот.
Дата добавления: 2016-03-26 | Просмотры: 1122 | Нарушение авторских прав |