|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология
|
II. Физиологические расы
Расы паразитов поражают определенные сорта растений-хозяев. Расы ржавчинного гриба Puccinia graminis были открыты американским ученым Стэкменом, расы Phytophthora infestans — шотландским селеционером Блэком в 1950-1960-е годы.
Понятие «физиологические» связано с тем, что способность рас преодолевать устойчивость одних сортов и неспособность преодолевать устойчивость других связана с различными физиологическими механизмами. Эти различия связаны с продуктами обмена веществ паразита и в клетках растения-хозяина.
Физиологические расы известны для патогенных бактерий, грибов и цветковых растений-паразитов. У вирусов различают штаммы, отличающиеся друг от друга по структуре ДНК и патогенности по отношению к растениям-хозяевам.
Для определения рас используют растения-дифференциаторы. Они относятся к сортам и гибридам, характеризующимся определенной вертикальной устойчивостью по отношению к возбудителю болезни. В расах присутствуют факторы вирулентности, в дифференциаторах — факторы вертикальной устойчивости. Между расой паразита и сортом растения-хозяина происходит дифференциальное (избирательное) взаимодействие. Оно заключается в следующем.
Если та или иная раса паразита преодолевает устойчивость сорта, то наблюдается реакция совместимости (рис.9). На пораженных органах проявляются основные симптомы (например, спороношение Phytophthora infestans или пустулы Puccinia graminis). Если раса паразита сталкивается с устойчивостью сорта, то на органах растения наблюдается реакция сверхчувствительности (рис.9) — основной механизм активного иммунитета растений. Она проявляется в виде мелких некрозов в местах внедрения спор возбудителя в растение-хозяина.
 Рис.9 Реакция совместимости Рис.9 Реакция совместимости
 Рис.10Реакциясверхчувствительности Рис.10Реакциясверхчувствительности
Авирулентная раса паразита (без факторов вирулентности) способна поражать только восприимчивые сорта (без факторов вертикальной устойчивости). Устойчивые сорта авирулентная раса, как правило, не поражает.
Вирулентная раса паразита (с фактором или факторами вирулентности) способна поразить восприимчивые сорта. Преодолеть устойчивость сорта с фактором или факторами вертикальной устойчивости вирулентная раса сможет в том случае, если они будут полностью соответствовать фактору или факторам вертикальной устойчивости. Если этого соответствия не будет хотя бы по одному из факторов, то раса паразита не сможет преодолеть устойчивость сорта.
Генетически факторы вирулентности определяются ѵіr-генами (virulence — вирулентность), а факторы вертикальной устойчивости— R-генами (resistance — устойчивость). В ряде исследований на основе генетического анализа было показано, что один фактор (вирулентности или вертикальной устойчивости) определяется одним геном (ѵіr- или R-геном соответственно) с полным доминированием. Вирулентность — признак рецессивный, а вертикальная устойчивость — доминантный. В последние годы посредством ПЦР-анализа ведется уточнение локусов (последовательностей ДНК), соответствующих тому или иному vir-гену паразита или R-гену растения-хозяина.
Для выявления рас Phytophthora infestans существует набор растений-дифференциаторов Блэка. Он состоит из сортов и гибридов с определенными R-генами: R1...R11. Набор Блэка признан селекционерами всего мира. Он хранится и поддерживается в Международном центре по картофелю (ІРС) в Перу. При определении рас растения-дифференциаторы (или листья с них, разложенные во влажные камеры) заражают суспензией конидий изолятов Ph. infestans. Заражение проводят в помещении с низкой освещенностью и высокой относительной влажностью, то есть в контролируемых условиях.
Дифференциатор R1 способен выявить две расы Ph. infestans, R1 и R2 — четыре, R1, R2 и R3 — восемь рас и т. д. (рис.11).

Рис. 11 Выявление рас Ph.infestans с помощью сортов дифференциаторов картофеля с одним фактором устойчивости
III.Изменчивость бактерий
Она происходит путем:
- наследуемых изменений, приводящих к появлению новых признаков за счет изменения последовательности ДНК — мутации;
- обмена участками ДНК — рекомбинации внутри генома;
- обмена генетической информацией между бактериями - трансдукции, конъюгации и трансформации (рис. 12).
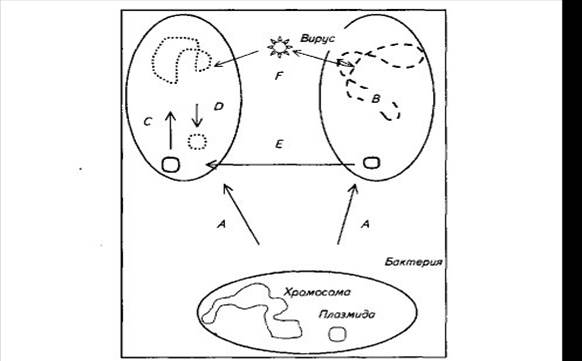
Рис.12 Схема основных процессов изменчивостей бактерий
Мутации. Скачкообразные наследуемые изменения, приводящие к появлению новых признаков, называются мутациями. В большинстве своем они приводят к исчезновению признака и снижению жизнеспособности, реже — к появлению нового признака, придающего организму новые свойства. Наиболее важные мутации у фитопатогенных бактерий происходят в генах авирулентности (аѵr) и генах группы сверхчувствительности (hrp), приводя к изменению расовой принадлежности и снижению или усилению агрессивности патогена. Например, у возбудителя черной бактериальной пятнистости томата и перца (Xanthomonas campestris pv. vesicatoria) и возбудителя сосудистого бактериоза капусты (X. campestris pv. campestris) замена нуклеотидов или удлинение пятинуклеотидного повтора в гене avrBs2 приводит к появлению новых рас, поражающих растения с генами устойчивости Bs2 (перец) или Rxc1 (абиссинская капуста). Изменение числа повторов нуклеотидов в середине гена avrBs3/Pth у Х.с. pv. vesicatoria и возбудителя бактериального рака цитрусовых X. citri также приводит к смене расы патогена.
Выявлено, что высокая нестабильность геномов фитопатогенных бактерий зачастую связана с активностью мобильных генетических элементов (транспозонов), фланкирующих гены, которые участвуют в патогенезе. Так, у Pseudomonas syringae гены авирулентности инактивируются вставкой мобильного элемента при заражении устойчивого растения, что также приводит к восстановлению вирулентности бактерии.
Различия в агрессивности бактерий и симптомах вызываемых ими болезней иногда определяются мутациями отдельных генов или изменением их регуляции. Например, генетические различия между ксантомонадами, вызывающими листовую пятнистость и сосудистый бактериоз у крестоцветных, заключаются в активизации разных протеаз и различной подвижности бактерий, которая зависит от температуры, pH и других факторов внешней среды.
Рекомбинации. Результатом рекомбинации внутри генома бактерии могут быть инверсия фрагментов ДНК, делеция (удаление) или перенос их в новое место и включение в хромосому или выделение из нее плазмид (кольцевых молекул ДНК с независимой репликацией). Большая часть изменений бактериального генома на уровне одного вида или близких видов бактерий связана именно с рекомбинацией и делецией или вставкой больших фрагментов генома. Рекомбинации чаще всего происходят между участками, имеющими сходные последовательности ДНК — нуклеотидные повторы. Именно рекомбинации оказывают наибольшее влияние на приспособленность бактерии к растению-хозяину за счет возникновения стабильных групп (блоков) генов — оперонов, вовлеченных в одну и ту же биохимическую или физиологическую реакцию.
Известно, что многие гены бактерий связаны в опероны или кластеры, вовлеченные в экспрессию одного признака.
Обмен информативной ДНК/РНК между геномами происходит путем:
- конъюгации — переноса ДНК между бактериями через особый секреторный аппарат — пиль (цилиндр, состоящий из белковых мономеров и соединяющий цитоплазму двух бактериальных клеток).
Дата добавления: 2016-03-26 | Просмотры: 839 | Нарушение авторских прав
1 | 2 |
|