|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Скэвенджер-рецептор для модифицированных ЛПНПРецептор для b-ЛПОНП Рецептор для комплексов ЛПНП-Ig Рецептор для других атерогенных ЛП, включая ЛП(а) Эндосома с ЛПНП-частицей Пул ХС и его эфиров в лизосомах Пул ХС в цитоплазме (липидные капли) Мультимеллярные везикулы с ЭХС 10. рецептор для ЛПВП3 11. ретроэндоцитоз ЛПВП2 –частиц, обогащенных ЭХС АХАТ - ацил-КоА-холестерин-ацилтрансфераза ГЭХ – гидролаза эфиров ХС
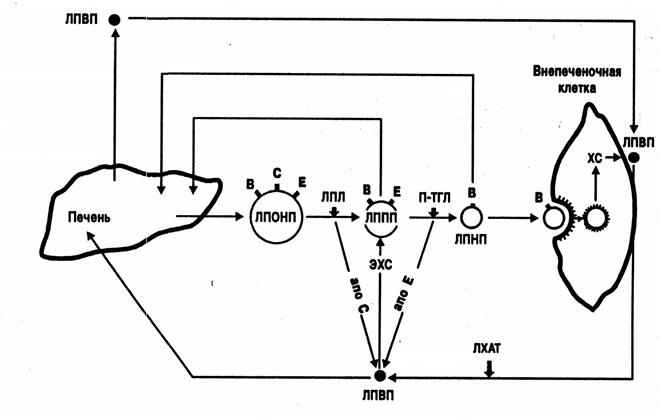
ЛПВП-частицы, обогащенные ХС и апо-Е, направляются в печень, где взаимодействуют с апо-В, Е -рецепторами, интернализуются и разрушаются. Таким образом, апо-Е выступает в роли вектора, направляющего частицы ЛПВП в печень для окисления содержащегося в них ХС. Доказано, что ХС ЛПВП является предпочтительным субстратом для образования в гепатоцитах желчных кислот. Взаимодействие ЛПВП с гепатоцитами, по существу, завершает процесс «обратного» транспорта ХС в организме. Таким образом, ЛПВП активно забирают ХС из ГМК, фибробластов, макрофагов, эндотелиальных клеток и направляют его в печень для окисления и удаления из организма. Подводя итоги, можно констатировать, что существующие в норме механизмы регуляции доставки и удаления ХС достаточно эффективно поддерживают ХС гомеостаз отдельной клетки и целостного организма(рис. 6).
Рис. 6. Транспорт ХС липопротеинами различных классов из печени в периферическую клетку и обратно. В, С, Е – соответствующие апопротеины (по А.Н.Климову, Н.Г.Никульчевой, 1995).
Генетические дефекты в данных системах могут быть причиной наследственных гиперхолестеринемий или дислипопротеинемий (ДЛП), ведущих к развитию атеросклероза. Однако частота встречаемости наследственных дефектов транспорта ХС не очень высока, в то время как сам атеросклероз распространен практически среди всей человеческой популяции в развитых странах мира. Каковы могут быть причины развития атеросклероза у лиц, не имеющих каких-либо генетических аномалий? Для того чтобы попытаться ответить на этот вопрос, необходимо уточнить, какие именно ЛП могут быть отнесены к т.н. «атерогенным», способным вызывать ДЛП и атеросклероз. 7.Патологические ЛП
1. b-ЛПОНП. При некоторых нарушениях ЛП обмена в крови у человека и некоторых животных обнаруживаются b-ЛПОНП или флотирующие b-ЛП. В частности, b-ЛПОНП появляются у экспериментальных кроликов, содержащихся на высокохолестериновой диете, а также у человека при III-ем типе ДЛП. Это – самостоятельная фракция ЛП, близкая по свойствам к ЛП промежуточной плотности, но отличающаяся от последних большей насыщенностью ХС, апо-Е-1 и путями катаболизма. Эти ЛП не превращаются в ЛПНП под действием печеночной липазы, как нормальные ЛП промежуточной плотности. Предполагается, что в результате структурных изменений они утрачивают сродство к этому ферменту и поэтому длительно циркулируют в крови. На макрофагах имеются специфические рецепторы к b-ЛПОНП, в результате взаимодействия с которыми происходит их проникновение в клетку, однако этот процесс не регулируется по механизму обратной связи, поэтому макрофаги трансформируются в пенистые клетки, характерные для атеросклеротического процесса.
2. ЛПВПХС. Эта фракция ЛПВП появляется у экспериментальных кроликов, находящихся на ХС диете, а также у людей, потребляющих пищу, богатую ХС. Это – фракция ЛПВП, перегруженных ХС, в результате чего они утрачивают способность к акцепции и обратному транспорту ХС от мембран периферических клеток в печень. Данные ЛП, таким образом, лишены антиатерогенных свойств.
3. Разновидность ЛП, обозначаемая как ЛП-Х. Эти ЛП появляются в крови при заболеваниях печени, сопровождающихся застоем желчи и желтухой. ЛП-Х имеют плотность, близкую к ЛПНП. Эти частицы имеют дисковидную форму, легко слипаются, образуя «монетные столбики». Благодаря своей высокой жесткости они способствуют повышению вязкости крови. Атерогенность этих ЛП в настоящее время является предметом обсуждения.
4. Липопротеин(а), ЛП(а), в англоязычной литературе - Lp(a). ЛП(а) – своеобразный липид-белковый комплекс, относящийся к апо-В-содержащим ЛП, или к ЛП, богатым ХС. ЛП(а) представляют собой сферические частицы, флотирующие при центрифугировании между ЛПНП и ЛПВП. По физико-химическим свойствам, химическому составу и особенностям катаболизма ЛП(а) отличается от ЛПНП. В отличие от ЛПНП, ЛП(а) имеет уникальный состав белковой компоненты, в частности, он содержит апопротеин (а) – высокогликозилированный полипептид, соединенный с апо-В дисульфидным мостиком (см. рис.7).
Рис. 7. Структура ЛП(а). Апо (а) – полипептид, соединенный с апо-В дисульфидным мостиком (по A.Scanu et al., 1987). На долю апо(а) приходится 20% массы белка, на долю апо-В – около 65% и на долю необычного для ЛП белка – альбумина – около 15%. Обнаружено близкое структурное сходство апо(а) с плазминогеном – одним из факторов свертывания крови. В настоящее время точно не установлены места синтеза и катаболизма ЛП(а). Частичная деградация этих ЛП происходит путем их связывания с апо-В, Е-рецепторами. Один из наиболее важных аспектов изучения этих ЛП – их роль в развитии атеросклероза. На основании эпидемиологических исследований удалось установить четкую связь между повышенной концентрацией ЛП(а) в крови и развитием коронарного атеросклероза с инфарктом миокарда. Повышенный уровень ЛП(а) определяется более чем у половины пациентов, перенесших инфаркт миокарда. Особенно высок уровень этого ЛП у больных, перенесших инсульт. Считается, что 20-30 мг/дл является предельной концентрацией ЛП(а) для здорового человека, превышение этого уровня рассматривается как патология. Важным является то, что повышенный уровень ЛП(а) в крови практически не снижается при применении известных в настоящее время гиполипидемических препаратов. ЛП(а) обнаружены в местах поражения сосудов, причем отложения апо(а) располагаются, главным образом, внеклеточно и в участках скопления фибриногена, что подтверждает связь ЛП(а) со свертывающей системой крови. Атерогенность ЛП(а) может быть обусловлена следующими факторами: · Апо(а), соединенный с апо-В, задерживает деградацию и удаление ЛП(а) из кровотока через классический рецепторный путь, способствуя более длительной циркуляции в крови, их модификации и поступления в клетки путем нерегулируемого эндоцитоза. · Апо(а), конкурируя с плазминогеном за места связывания на фибрине, ингибирует фибринолиз и тем самым способствует тромбообразованию. · ЛП(а) или очищенный апо(а) стимулирует рост ГМК аорты человека в культуре ткани. Поскольку пролиферация ГМК является одним из важнейших клеточных механизмов развития атеросклероза, предполагается, что ЛП(а) таким образом способствуют развитию атеросклеротической бляшки. Популяционные исследования, проведенные в различных регионах мира показали, что повышение уровня ЛП(а) в крови представляет собой самостоятельный и независимый фактор риска ИБС – наиболее частого и тяжелого клинического проявления атеросклероза.
8. Модифицированные липопротеины Модифицированные ЛП образуются в организме из нормально синтезированных и секретированных в кровь ЛП. Причиной их модификации могут быть выброс клетками свободных радикалов и продуктов ПОЛ, повышенная концентрация в крови, межклеточной жидкости и сосудистой стенке некоторых метаболитов (например, глюкозы), а также ферментов различного спектра действия. Важно отметить, что модифицированные ЛП образуются также и при замедлениискорости деградации нативных ЛПНП, например: нарушение рецепторного захвата ЛПНП® снижение скорости катаболизма ЛПНП и удлинение времени их циркуляции ® воздействие метаболитов, продуктов ПОЛ и различных ферментов® модификация (= приобретение атерогенности) ЛПНП. Возникновение концепции модифицированных ЛП относится к концу 70-х – началу 80-х годов и тесно связано с открытием специфических клеточных рецепторов к ним. Эксперименты, проведенные на обезьянах и крысах, показали, что если животным ввести внутривенно ЛПНП с блокированными лизиновыми и аргининовыми остатками, которые не распознаются апо В, Е-рецепторами, то такие химически модифицированные ЛП все равно удаляются из кровотока и, во многих случаях быстрее нативных [Mahley R. Et al., 1980]. Это послужило основанием предположить, что наряду, с классическим рецепторным путем удаления ЛПНП из кровотока, должны быть и другие, одним из которых оказался так называемый скэвенджер-путь (дословно: путь уборки мусора), осуществяемый клетками РЭС. Предположение о наличии такого пути захвата и катаболизма ЛПНП было сделано M. Brown и соавт. (1979) на основании опытов, проведенных на культуре макрофагов с использованием химически модифицированных (ацетилированных) ЛПНП. Оказалось, что ацетилированные ЛПНП, почти нераспознаваемые апо-В, Е рецепторами фибробластов, активно захватывались макрофагами, причем в макрофагальных клетках происходило значительное накопление ЭХС. Последующие исследования позволили установить, что захват модифицированных ЛПВП также осуществляется при участии рецепторов, но другой природы, которые стали называть скэвенджэр-рецепторами, или ацетил-ЛПНП-рецепторами. Помимо макрофагов, скэвенджер-рецепторы имеют и другие клетки РЭС: звездчатые ретикулоэндотелтоциты (купферовские клетки печени), ретикулярные клетки селезенки и эндотелиальные клетки сосудов.
- богатый цистеином участок
- коллагеноподобный участок
- спиралевидный витой шнурок
- участок, соединяющий разъединенные пептидные мономеры - участок фиксации к мембране - внутриклеточный участок
Рис. 8. Схематическая модель бычьего скэвенджер-рецептора (по T.Kodama et al., 1990). Недавно, благодаря многолетним исследованиям четырех лабораторий в США, удалось выделить в чистом виде скэвенджер-рецептор и установить его первичную структуру. Скэвенджер-рецептор представляет собой полипептидный тример, каждый мономер которого состоит из 453 ак и характеризуется высокой степенью гликозилирования. Рецептор распознает как конформацию лиганда, так и его заряд (многие химически модифицированные ЛПНП имеют избыточный отрицательный заряд благодаря блокированию положительно заряженных аминогрупп лизина, либо–присоединению дополнительных карбоксильных групп). Структура скэвенджер-рецептора представлена на рис. 8. Важными участками рецептора, выполняющими функцию взаимодействия с лигандом, являются богатый цистеином (1) и коллагеноподобный (2) домены; последний по своей трехнитчатой правозакрученной спирали и аминокислотной последовательности напоминает фиброзный коллаген. Выяснилось, что и сам коллаген обладает способностью связывать перекисно-модифицированные ЛПНП. Вероятно, именно по этой причине они откладываются в богатых коллагеном тканях – сухожилиях, коже и развивающихся атеросклеротических поражениях. Особенностью скэвенджер-захвата является то, что поступление модифицированных ЛП таким путем в клетку не регулируется или слабо регулируется помеханизму отрицательной обратной связи. Содержание ХС в клетке при этом линейно нарастает в зависимости от концентрации в окружающей среде ЛП, содержащих ХС. Несмотря на то, что поступление ХС в клетку путем скэвенджер-захвата подавляет синтез собственно стерина, а также образование апо- В-Е рецепторов, это не спасает клетку от накопления в ней значительных количеств ХС, преимущественно в виде эфиров и, в меньшей степени, в виде свободного ХС и кристаллов моногидрата. С помощью скэвенджер-рецепторов осуществляется захват и других химически модифицированных ЛПНП (сукцинилированных, ацетоацетилированных, обработанных малоновым диальдегидом и др.), а также ЛПНП, подвергнутых пероксидации. Помимо скэвенджер и классических апо-В,Е и ЛПВП-рецепторов, макрофаги имеют и ряд других рецепторов (рис. 5), в частности: · Рецепторы к b-ЛПОНП; · Рецепторы к декстран-сульфату, которые взаимодействуют с комплексом ЛПНП-гликозаминогликан; · Рецепторы к Fc –фрагменту Ig и к C3-компоненту комплемента, с помощью которых макрофаги могут захватывать иммунные комплексы, включающие апо-В-содержащие ЛП (ЛПНП) в качестве антигена.
Таким образом, макрофаги, имея большой набор рецепторов, могут связывать, интернализировать и разрушать как нативные, так и различным образом модифицированные ЛП. Захват нативных ЛП происходит при участии классических апо-В,Е-рецепторов, и этот тип захвата никогда не сопровождается накоплением ЭХС в клетке. Захват модифицированных ЛП протекает по типу скэвенджер-захвата и сопровождается накоплением в клетке ЭХС и трансформацией макрофагальной клетки в «пенистую». Последняя представляет собой клетку, «нафаршированную» липидными вакуолями, содержащими ЭХС. Факт значительного накопления ЭХС в макрофагах и трансформации этих клеток в пенистые при захвате модифицированных ЛПНП позволил сделать вывод, имеющий большое теоретическое и практическое значение: ответственными за развитие атеросклеротического процесса являются не нативные, а модифицированные в организме человека ЛПНП. Именно такие ЛП должны рассматриваться как атерогенные. Нативные ЛПНП, вопреки широко распространенному заблуждению, не относятся к атерогенным ЛП.
Атерогенность ЛПНП, выделенных из крови людей (как здоровых, так и больных атеросклерозом), можно определить в лабораторных условиях. Для этого производят инкубацию ЛПНП, предварительно помеченных радиоактивным йодом, с культурами фибробластов и макрофагов. Если выделенные ЛПНП активно захватываются макрофагами и слабо – фибробластами, то они – атерогенны, если наоборот, то – нет.
Дата добавления: 2016-03-26 | Просмотры: 1053 | Нарушение авторских прав |