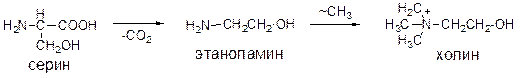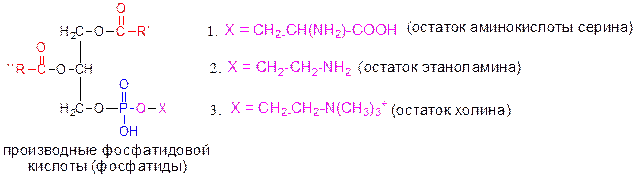|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Эти этерифицирующие агенты могут взаимно превращаться в процессе метаболизма.
1. Фосфатидилсерины (серинкефалины); 2. Фосфатидилэтаноламины (коламинкефалины); 3. Фосфатидилхолины (лецитины).
Фосфоинозитиды являются важной группой глицерофосфолипидов: a) фосфтидилинозит, b) фосфатидилинозитфосфат c) фосфатидилинозитдифосфат. Фосфоинозитиды присутствуют почти во всех животных тканях (5-10% липидного фосфора), особенно высоко их содержание в мембранах нервных клеток (рис. 7).
Рис. 7 Строение фосфоинозитидов
Плазмалогены – этолипиды с простой эфирной связью.Они являются менее распространенными, по сравнению со сложными глицерофосфолипидами. Плазмалогены содержат остаток винилового спирта, связанный простой эфирной связью с С-1 L-глицеро-3-фосфата. Двойная связь в виниловом фрагменте имеет цис -конфигурацию (рис. 8). На долю плазмалогенов приходится до 10% фосфолипидов мозга и мышечной ткани.
Рис. 8 Пример структуры плазмалогенов – L-фосфатидальэтаноламинов (n = 9-15).
Сфинголипиды – структурные аналоги глицеридов, в которых вместо глицерина используется сфингозин – длинноцепочечный двухатомный аминоспирт. Примером сфинголипидов служат церамиды – N-ацильные производные сфингозина, аминогруппа в которых ацилирована жирной кислотой. Важную группу сфинголипидов составляют сфингомиелины. В сфингомиелинах гидрокси-группа у С-1 церамида ацилирована фосфохолиновой группировкой (сфингомиелины могут быть также отнесены и
к фосфолипидам).
Сфингомиелины являются наиболее распространенными сфинголипидами, находятся в мембранах животных, особенно богата ими нервная ткань, они обнаружены также в тканях почек, печени, входят в состав липидов крови. Гликолипиды представляют собой производные сфингозинсодержащих липидов, они содержат углеводные остатки и не содержат фосфорной кислоты и азотсодержащих оснований. Типичные производные гликолипидов – цереброзиды и г англиозиды. Цереброзиды - производные церамидов, в которых остаток церамида связан с D-галактозой или D-глюкозой β-гликозидной связью. Цереброзиды входят в состав оболочек нервных клеток (рис.9) Ганглиозиды – богатые углеводами сложные липиды, были обнаружены в сером веществе головного мозга. В структурном отношении ганглиозиды сходны с цереброзидами, отличаясь лишь тем, что вместо моносахарида они содержат сложный олигосахарид, в состав которого входит по крайней мере один остаток N-ацетилнейраминовой кислоты (рис.9).
Рис.9 Строение а) цереброзидов и b) ганглиозидов
Амфипатические свойства фосфолипидов. Характерной особенностью сложных липидов является их амфипатические свойства, обусловленные наличием как неполярных гидрофобных, так и высокополярных ионизированных гидрофильных группировок. В фосфатидилхолинах, например, углеводородные радикалы жирных кислот образуют два неполярных гидрофобных «хвоста», а карбоксильная, фосфатная и холиновая группы – полярную гидрофильную «головку» (рис.10). Рис. 10 Строение лецитина (фосфатидилхолина) обусловливает его амфипатические свойства
Молекулы фосфолипидов в контакте с водой стремятся расположиться так, чтобы углеводородные цепи находились в контакте только с другими аналогичными цепями во внутренней полости, а полярные группы липидов располагаются на внешней поверхности. Таким путём образуется двойной слой толщиной в две молекулы (рис.11). Двойные слои составляют основу клеточной мембраны. В составе биомембран, ограничивающих живые клетки и их внутренние органеллы, липидные компоненты обеспечивают высокое электрическое сопротивление мембраны, ее непроницаемость для ионов и полярных молекул и проницаемость для неполярных веществ. Таким образом, мембраны сохраняют концентрационные различия между различными частями клетки или между клеткой и её водным окружением.
Рис. 11 Образование двойного слоя амфипатическими молекулами. Тонкие различия в физических свойствах клеточных мембран контролируются степенью ненасыщенности цепей жирных кислот в молекулах фосфолипидов. Известно, что глицериды, содержащие цис -двойные связи, плавятся при более низких температурах, чем насыщенные глицериды. Поэтому мембраны, состоящие из ненасыщенных фосфолипидов, являются более мягкими и позволяют молекулам проходить через них быстрее, чем через аналогичные мембраны, построенные из сложных эфиров насыщенных жирных кислот. Отмечалось также, что клетки, которые живут и размножаются при относительно низких температурах, имеют больше ненасыщенных жирных кислот в своих мембранах, чем клетки, живущие при более высоких температурах. Чтобы полярные питательные вещества и ионы могли пройти через внутреннюю часть этих липидных мембран, необходимо изменить их полярность. В мембранах имеются молекулы белков, которые окружают эти нерастворимые в углеводородах питательные вещества и ионы за счёт своей сильно полярной внутренней части и проводят их через мембрану. При соответствующей энергии проведения и благодаря разной способности белков образовывать комплексы и окружать ионы и молекулы, можно создать очень большие различия концентраций между внутренней частью клетки и окружающей её средой. Такая модель устройства биомембраны получила название жидкостно-мозаичной, рис.12.
Рис.12. Жидкостно-мозаичная модель биомембраны.
Дата добавления: 2016-06-06 | Просмотры: 1033 | Нарушение авторских прав |