|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Малые системы нейроновМОЗГ Содержание Предисловие редактора перевода Д. Хьюбел. МОЗГ Ч. Стивенc НЕЙРОН Э. Кэндел МАЛЫЕ СИСТЕМЫ НЕЙРОНОВ У. Hayта, М. Фейртаг ОРГАНИЗАЦИЯ МОЗГА У. Коуэн РАЗВИТИЕ МОЗГА Л. Иверсен ХИМИЯ МОЗГА Д. Хьюбел, Т. Визель ЦЕНТРАЛЬНЫЕ МЕХАНИЗМЫ ЗРЕНИЯ Э. Эвартс МЕХАНИЗМЫ ГОЛОВНОГО МОЗГА, УПРАВЛЯЮЩИЕ ДВИЖЕНИЕМ М. Гешвинд СПЕЦИАЛИЗАЦИЯ ЧЕЛОВЕЧЕСКОГО МОЗГА С. Кити ЗАБОЛЕВАНИЯ ЧЕЛОВЕЧЕСКОГО МОЗГА Ф. Крик МЫСЛИ О МОЗГЕ ЛИТЕРАТУРА Предисловие редактора перевода В 1979 году американский междисциплинарный (это определение мне кажется более точным, чем «научно-популярный») журнал «Scientific American» почти целиком посвятил свой сентябрьский номер описанию наиболее значительных достижений в области изучения строения и деятельности мозга. В качестве авторов выступили ученые, непосредственно причастные к этим достижениям, внесшие в развитие науки о мозге свой ощутимый вклад. Такие имена, как Д. Хьюбел, Т. Визель, Э. Кэндел, Э. Эвартс, У. Наута и другие, хорошо известны специалистам всего мира, без этих имен трудно представить себе современную нейрофизиологию. В данном случае они предстали перед читателем в качестве увлекательных рассказчиков, весьма осторожных в оценке полученных результатов, зато ясно отдающих себе отчет в том, как неполны и противоречивы наши реальные знания о деятельности мозга — сложнейшего и загадочного порождения Природы. Доступная широким кругам читателей информация «из первых рук» всегда остается идеалом популяризации любой области знания. В лежащей перед нами книге это условие соблюдено максимально. Прогресс науки о мозге неоспорим и очевиден. По справедливому замечанию Д. Хьюбела, нейрофизиолог 50-х годов был бы поражен, если бы ему сказали, как много нового станет нам известно тридцать лет спустя. Непрерывное накопление информации о биологически активных веществах-передатчиках и модуляторах нервного возбуждения — чем - то напоминает ситуацию в области ядерной физики, где с каждый годом увеличивается семейство открытых элементарных частиц. Постепенно проясняющееся значение нарушений химизма мозга в генезе психических заболеваний коренным образом меняет облик описательной психиатрии. Поистине революционизирующее влияние на представления о физиологических основах человеческой психики-сознания и мышления оказало открытие функциональной специализации двух полушарий. Эти и другие успехи нейрофизиологии подробно освещены в настоящем издании. Вместе с тем содержащиеся в нем материалы отнюдь не исчерпывают того, что достигнуто современной наукой. Например, в статьях американских коллег совершенно отсутствуют результаты, полученные советскими исследователями мозга. Это касается прежде всего таких разделов, как изучение фундаментальных закономерностей физиологии нервной клетки (П. Г. Костюк, А. И. Шаповалов), физиологии зрения (А. Л. Бызов, В. Д. Глезер, И. А. Шевелев) и слуха (Г. В. Гершуни), нейрофизиологических механизмов организации движения (В. С. Гурфинкель, Г. Н. Орловский, М. Л. Шик). Весьма значителен вклад советских исследователей в анализ клеточных механизмов обучения-формирования условных рефлексов (Н.Н.Василевский, Л.Л.Воронин, У. Г. Гасанов, А. Б. Коган, М. Н. Ливанов, М. Я. Рабинович, В. С. Русинов, Е. Н. Соколов, В. Б. Швырков, Г. И. Шульгина, М. М. Хананашвили и многие другие). Принципиальное значение для понимания закономерностей высших форм интегративной деятельности мозга имеют теоретические обобщения П.К.Анохина, Э.А. Асратяна, И. С. Бериташвили, Н.А.Берн штейна, Л. А. Орбели и других отечественных ученых. Из перечня охваченных тем составителями исключены сведения, касающиеся строения и функций тех мозговых структур, которые непосредственно вовлечены в процесс актуализации важнейших потребностей организма - голода, жажды, полового влечения, в генез эмоциональных реакций — ярости, страха, удовольствия и т. п. В результате становится непонятным, как связаны столь тщательно изученные сенсорные системы (зрение, слух и т.п.) с не менее глубоко проанализированными механизмами организации движений. Иными словами, где локализованы структуры мозга, решающие вопрос о том, на какие воздействия внешней среды и каким образом должен отвечать живой организм? Здесь-то и возникает та проблема «гомункулуса», о которой пишет в своей заключительной статье Ф. Крик. С редкой проницательностью и остротой один из первооткрывателей генетического кода ставит этот вопрос перед умом естествоиспытателя. Как часто самые разные авторы, вне зависимости от того, оперируют ли они понятием «информации», «нейропсихического кода» или «субъективного образа», забывают о существовании проблемы «получателя информации, смотрящего на экран маленького телевизора в мозгу», или заменяют гомункулуса таким психологическим понятием, как «личность». Ф. Крик не предлагает своего решения проблемы, но совершенно справедливо требует раз и навсегда расстаться с идеей гомункулуса в любых ее вариантах, поскольку отказ от ложного пути сам по себе представляет немалый успех ищущей мысли. В отличие от всех других объектов естествознания изучение мозга человека отягощено возможностью интроспекции, т.е. возможностью взгляда «изнутри». Очевидность и наглядность представлений о своем собственном внутреннем мире затрудняют постижение законов деятельности мозга в несопоставимо большей степени, чем наглядность движения Солнца вокруг Земли столетиями искажала картину Солнечной системы. Отсюда и возникает вопрос, прозвучавший в самом начале: может ли мозг понять мозг, может ли мозг понять разум? Лучшим ответом на этот вопрос являются работы, помещенные в настоящем выпуске, реальные достижения современной науки. Быть может, одно из важных достоинств этих статей как раз и состоит не только в информировании читателя о том, что уже познано, но и в уточнении формулировки нерешенных проблем, в определении стратегических задач дальнейших исканий. П. В. Симонов
Три основных орудия нейробиолога (символическое изображение)-это микро-скоп, избирательное окрашивание нервной ткани и микроэлектрод. На рисунке приведена световая микрофотография нескольких окрашенных по Гольджи нейронов в срезе ткани мозга из зрительной коры обезьяны; длинная игла - кончик микроэлектрода, расположенного как бы для записи электрических импульсов, которые генерирует одна из клеток. Фотография получена Ф. Горо (F. Goro). Д. ХЬЮБЕЛ Мозг Вступление к выпуску журнала, посвященного нейробиологии и ее центральной проблеме: как работает человеческий мозг? Несмотря на значительные достижения, этот вопрос остается одним из самых трудных в современной науке Может ли мозг понять мозг? Может ли он понять разум? Что он такое - гигантский компьютер, или какая-либо иная гигантская машина, или же нечто большее? Эти вопросы задаются постоянно, и было бы полезно избавиться от них. Утверждение, что мозг не может быть понят мозгом, аналогично афоризму, что человек не в состоянии поднять себя сам за ушки собственных сапог. Но такая аналогия не бесспорна. Несомненно, даже беглый взгляд на то, что достигнуто на пути к пониманию мозга, убедит любого читателя этого выпуска «Scientific American», что со времен френологии мы ушли далеко вперед. Темп этого движения сейчас очень быстрый. Таким образом, с учетом всех практических задач нейробиологи работают в ожидании того, что смогут понять мозг, и в настоящее время они в этом преуспевают. Думаю, что трудности, связанные с вопросами подобного рода, носят семантический характер. Они несут на себе груз таких слов, как «понимать» и «разум» - слов для многих целей полезных, но с расплывчатыми очертаниями и неуместных в применении к этим вопросам, которые из-за них становятся бессмысленными или неразрешимыми. Мозг представляет собой ткань - запутанную, сложно сотканную, сложнее всего, что нам известно во Вселенной, однако, как любая живая ткань, она состоит из клеток. Вместе с тем, хотя это высокоспециализированные клетки, их активность подчинена законам, которые управляют любыми другими клетками. Их электрические и химические сигналы можно обнаружить, зарегистрировать и истолковать; вещества, из которых они состоят, можно определить; связи, образующие густую сеть, можно проследить. Короче говоря, мозг можно изучать совершенно так же, как изучают любой другой орган, скажем почку. Проблема возникает тогда, когда мы ставим вопрос о понимании, потому что такое слово привносит сложность внезапного откровения или прояснения, предполагает наличие такой минуты, когда можно сказать, что из темного туннеля мы выходим на свет. Но мне не ясно, может ли в данном случае наступить такая минута и узнаем ли мы о ее наступлении. Исследование мозга - это древняя область науки, а область спекуляций о мозге — еще древнее. Его изучение сильно ускорилось к концу XIX в.; новые методики, разработанные после второй мировой войны, привели к значительным успехам, и в последнее десятилетие нейробиология стала одной из самых активных отраслей науки. Следствием этого недавно явился подлинный взрыв открытий и прозрений. Но тем не менее изучение мозга только начинается. Констатация невероятной сложности мозга стала штампом, однако же это факт. Проблема понимания работы мозга в чем-то сходна с проблемой понимания структуры и функции белков. Каждый организм содержит миллионы сложных изощренных молекулярных комбинаций, причем один человек совершенно отличен от другого. Для того чтобы детально изучить структуру хотя бы одного белка, по-видимому, потребуются годы, не говоря уже о том, чтобы узнать точно, как он работает. Если понять белки — это значит узнать, как все они работают, то перспективы здесь, пожалуй, отнюдь не радужные. Точно так же мозг состоит из очень большого числа (хотя и не миллионов) функциональных подразделений, из которых каждое обладает своей особой архитектоникой и своей сетевой схемой; а дать описание одного из них вовсе не значит описать их все. Поэтому понимание пойдет медленно (хотя бы по практическим причинам — в зависимости от времени и числа занятых этим людей), неуклонно (надо надеяться) и будет развиваться по асимптоте, безусловно с прорывами, но вряд ли достигнет конечной точки. Разум — это тоже полезное слово, но (увы!) еще более расплывчатое. Поскольку оно поддается лишь нечеткому определению, говорить о его понимании (не слова, а того, к чему оно относится) - это значит говорить об упражнении в мыслительной гимнастике, которая, по-видимому, лежит за пределами естественных наук. Математик Г. Харди (G. Hardy) будто бы сказал, что математик — это тот, кто не только не знает, о чем говорит, но и не интересуется этим. Тех, кто углубляется в такие вопросы, как физиология разума, вероятно, это занимает, но я не представляю себе, как они когда-либо смогут ответить на эти вопросы. Число нервных клеток, или нейронов, из которых состоит примерно полтора килограмма человеческого мозга, достигает порядка 1011 (сто миллиардов) или же на порядок больше или меньше. Вокруг нейронов лежат, их поддерживают и питают глиальные клетки, которых тоже очень много. Типичный нейрон имеет тело (диаметром от 5 до 100 микрометров), от которого отходит одно крупное волокно, аксон, и множество мелких ветвей, дендритов. Аксон отдает веточки в начале и часто сильно ветвится в конце. Говоря в общем, дендриты и тело клетки принимают поступающие сигналы; тело клетки комбинирует и интегрирует их (грубо говоря, усредняет) и выдает выходные сигналы; оно также поддерживает общее состояние клетки; аксон проводит выходные сигналы к своим окончаниям, которые передают информацию следующей группе нейронов. Сигналы бывают двоякими - электрическими и химическими. Сигнал, генерируемый нейроном и проводимый по его аксону, представляет собой электрический импульс, но от клетки к клетке он передается молекулами передатчиков, медиаторов - веществ, которые перемещаются через особый контакт - синапс - между структурой, доставляющей информацию (окончанием аксона или, в отдельных случаях, дендритом), и структурой, воспринимающей ее (дендритом, телом клетки, или, в отдельных случаях, окончанием аксона). К одному нейрону обычно подходят отростки сотни или тысячи других нейронов, и он в свою очередь отдает веточки к сотням или тысячам других нейронов. Этого, пожалуй, достаточно, чтобы сделать возможной попытку сравнить мозг с компьютером. Большинство нейробиологов при обсуждении этого вопроса согласятся с тем, что мозг можно рассматривать как машину, которая не обладает свойствами, лежащими за пределами возможностей научного исследования. Но верно и то, что с этим согласятся не все нейробиологи. В то же время несомненно все будут согласны с тем, что компьютер — это машина и ничего больше. И таким образом, в зависимости от вкусов и убеждений, мозг и компьютер в определенном смысле или сходны по своим основным качествам, или даже совершенно различны. По моему мнению, нельзя решить этот вопрос рассуждениями.
Увеличение размеров большого мозга у позвоночных. В одном масштабе показан мозг некоторых различных классов. У рыб, амфибий, рептилий и птиц большой мозг невелик. У хищных млекопитающих и особенно у приматов величина и сложность мозга резко возрастают. Если считать машинами и мозг и компьютер, то в таком случае как их сравнивать? Такое упражнение представляет интерес. Компьютеры изобретены человеком и поэтому совершенно понятны, если принять, что человеческие существа вообще что-нибудь понимают; но чего люди не знают, это какими окажутся будущие компьютеры. Мозг создан эволюцией и во многих важных отношениях остается непонятным. Обе машины перерабатывают информацию и обе имеют дело с сигналами, грубо говоря, электрическими. В самых крупных вариантах и та и другая машина содержит множество элементов. Однако в этом между ними имеется интересное различие. Биологическим путем клетки производятся достаточно просто, и нейроны создаются в поистине огромных количествах. Между тем, умножить число элементов в компьютере, по-видимому, не так легко, даже несмотря на то, что это число быстро растет. Но если принять, что элементами нервной системы являются не нейроны, а синапсы, то я не могу себе представить, что компьютеры могли бы сравняться с ней. Никто не станет гадать, сколько в мозгу синапсов, но число 1014(100 триллионов) не кажется невероятным. Еще большее значение имеет одно качественное различие. Мозг не подчиняется чему-либо подобному линейной программе, во всяком случае, те его части, о которых нам хоть что-нибудь известно. Он скорее сходен с цепью в радиоприемнике или телевизоре, а может быть, с сотнями или тысячами таких цепей, соединенных последовательно и параллельно, со множеством перекрестных соединений. Мозг, по-видимому, основан на принципе относительно жестко запаянного блока из сложных цепей, элементы которого работают на низких скоростях, измеряемых тысячными долями секунды; компьютер подчиняется программам, содержит гораздо меньше элементов и работает на скоростях, при которых имеют значение миллионные доли секунды. В числе мозговых сетей должно быть много таких, которые служат для поддержания эволюции, связанной с борьбой за существование и половыми инстинктами. Компьютер пока что свободен от всего этого; он эволюционирует другими способами. Как надо изучать такой орган, как мозг? Основной подход состоит, разумеется, в том, чтобы изучить его составные части, а затем попытаться узнать, как они действуют сообща. Это осуществляется главным образом на животных, а не на человеке. Принципы нейронной функции удивительно сходны у столь далеких друг от друга животных, как улитка и человек; большая часть того, что известно о нервном импульсе, изучена на кальмаре. Даже основные структуры головного мозга так схожи. например у кошки и человека, что нередко не имеет значения, чей мозг изучать. Наряду с этим нейробиология отличается широким диапазоном привнесенных в нее подходов и методик, от физики и биохимии до психологии и психиатрии. Ни в одной другой области исследования не так важен этот широкий подход, который здесь начали применять в последние годы. Нейроанатомия и нейрофизиология — две традиционные смыкающиеся ветви нейробиологии. Анатомия стремится описать различные элементы мозга и способы их объединения; физиология занимается изучением того, как эти части работают вместе. Исследователи этих двух областей пошли было разными путями и даже обосновались на разных университетских кафедрах, но в действительности они тесно связаны друг с другом. Большинство современных нейроанатомов не довольствуются простым описанием структуры и пространственных отношений как таковых, а идут дальше и задаются вопросом, каково назначение этих структур и связей. В свою очередь физиология невозможна без анатомии. На всех этапах развития и нейроанатомия и нейрофизиология вынуждены были выжидать, пока физические науки не снабдят их необходимыми приборами и методиками. Нейрон так мал, что невооруженному глазу представляется лишь в виде точки, и также настолько мал, что его сигналы нельзя зарегистрировать простыми электродами. Для того чтобы выйти за пределы самых примитивных исследований, анатомии потребовался сначала световой, а затем электронный микроскоп, а физиологии — микроэлектрод. Для обеих наук имело значение создание специальных методик избирательного окрашивания нервной ткани. Главные достижения нейроанатомов начала нашего века состояли в признании нейрона основным элементом нервной ткани и в открытии упорядоченности и специфичности межнейронных связей. Физиологи заложили прочную основу тем, что поняли, какими электрическими и химическими способами нейрон передает свои сигналы. Эти два вида достижений ни в коей мере не раскрыли, как работает мозг, но они служат совершенно необходимым фундаментом для этого. Один из способов увидеть, как далеко вперед ушла нейробиология (и, безусловно, как невероятно далеко ей еще надо идти), состоит в том, чтобы рассмотреть некоторые исторические этапы на пути к современному пониманию мозга и дать краткий обзор теперешнего состояния исследований в некоторых разделах этой области. Почему, прежде всего, так трудно было установить, что отдельный нейрон служит основным элементом нервной ткани? Главными препятствиями являлись ничтожные размеры, фантастические формы и огромное разнообразие форм этих клеток, а также тот факт, что ветви, принадлежащие соседним клеткам, тесно переплетаются между собой. Слово «клетка» вызывает в воображении представление о кирпичике или кусочке студня; на самом же деле нейрон выглядит как миниатюрное деревце с разветвленной кроной, ствол которого имеет толщину от 10 до 20 мкм (тысячных долей миллиметра) и длину от 0,1 мм иногда до метра. Для того чтобы увидеть отдельные нейроны, требуется не только микроскоп, но и краситель, выделяющий их из окружения. Обычно нейроны упакованы так плотно, что в любом участке сотни их системами своих ветвей переплетены в густую чащу, в которой соседние веточки разделены пленками жидкости толщиной всего лишь около 0,02 мкм, и таким образом фактически все пространство занято клетками и их разнообразными отростками; а если на данном участке окрашены все клетки, то в световой микроскоп виден лишь плотный мазок. Поэтому самым важным достижением нейроанатомии (после самого микроскопа) было открытие, сделанное примерно в 1875 г. итальянским анатомом К. Гольджи. Он изобрел метод, при котором одновременно окрашивается, по-видимому, в случайном порядке, лишь очень малая доля всех клеток данного участка, но зато эти клетки окрашиваются целиком. При хорошо выполненном окрашивании по Гольджи вместо невразумительной мазни на препарате видны лишь несколько нейронов, но каждый из них полностью, со всеми своими ветвями. Просмотрев много срезов мозга, окрашенных по Гольджи, анатом может дать перечень разных клеток в этой ткани. Доныне никто не знает, как и почему срабатывает метод Гольджи, окрашивая полностью одну из 100 клеток и совершенно не затрагивая все остальные. Современник Гольджи, испанец С. Рамон-и-Кахал посвятил всю свою поразительно плодотворную жизнь приложению нового метода практически ко всем частям нервной системы. Его гигантская «Histologie du systeme nerveux de l'homme et des vertebres» («Гистология нервной системы человека и позвоночных животных»), впервые опубликованная в 1904 г. на испанском языке, до сих пор остается самой фундаментальной монографией по нейробиологии. Во времена Кахала шел спор о степени непрерывности между клетками. Отделены ли клетки одна от другой полностью, или же они соединены от аксона к дендриту в непрерывную сеть? Если бы существовала непрерывность протоплазмы, то сигналы, генерируемые одной клеткой, могли бы переходить в соседнюю, не прерываясь; если же непрерывности нет, то тогда должен существовать специальный процесс генерации сигналов заново в каждой клетке. На препаратах Кахала, окрашенных по Гольджи, выявляется множество обособленных, полностью окрашенных клеток и никогда не было видно ничего похожего на сеть. Таким образом, его первым большим вкладом явилось представление о нервной системе как о совокупности отдельных, обособленных клеток, которые сообщаются друг с другом с помощью синапсов.
К. Гольджи (1844-1926) дал ключ к микроскопическому исследованию нервной системы, разработав (1875 г.) метод избирательного окрашивания нервной ткани, при котором в данном участке одновременно окрашивается лишь небольшая доля клеток, но зато полностью. Фотография сделана в начале восьмидесятых годов, в бытность Гольджи профессором гистологии и общей патологии в Университете в Павии. С. Рамон-и-Кахал (1852-1934) посвятил свою жизнь тщательному изучению при помощи метода Гольджи буквально всех частей нервной системы множества разных животных. Признанием трудов Гольджи и Рамон-и-Кахала по изучению строения нервной системы явилось присуждение им в 1906 г. Нобелевской премии по физиологии и медицине. Рамон-и-Кахал преподавал в нескольких испанских университетах и дольше всего в Мадридском. Эта фотография сделана самим Рамон-и-Кахалом в двадцатых годах. Кахал внес второй вклад, пожалуй, еще более значительный. Он собрал множество данных о том, что невероятно сложные связи между нейронами не случайны, как иногда думали, а представляют полную противоположность случайности, т.е. высоко структурированы и специфичны. Он дал исчерпывающее описание архитектоники десятков различных структур мозга и в каждом случае идентифицировал и классифицировал разные клетки, а иногда показывал, насколько позволяли его методы, как эти клетки связаны между собой. Со времени его трудов стало ясно, что если нейробиолог хочет понять мозг, он должен не только изучить, как построены разные его части, но и раскрыть их назначение и детально исследовать их работу как отдельных структур и в совокупности. Но сначала нужно узнать, как отдельный нейрон генерирует сигналы и передает их следующей клетке. Хотя Кахал, быть может, никогда не формулировал проблемы, связанные с пониманием мозга, с полной четкостью именно в этих выражениях, но знакомясь с его трудами, нельзя не извлечь из них такого вывода. Долгое время нейроанатомам приходилось довольствоваться все более подробными описаниями, основанными на световой микроскопии с окрашиванием по Гольджи и по Нисслю (Nissl) (последнее выделяет тела отдельных клеток без дендритов и аксонов). Первым действенным орудием прослеживания связей между разными мозговыми структурами — например, между разными областями коры большого мозга или между корой и стволом мозга и мозжечком - явился метод окрашивания, который предложил в начале пятидесятых годов нашего века в Голландии У. Наута (W. Nauta), который теперь работает в Массачусетском технологическом институте. Этот метод основан на том, что при разрушении нейрона (механическим, электрическим или тепловым воздействием) отходящее от него нервное волокно дегенерирует и, пока оно еще не совсем исчезло, окрашивается иначе, чем соседние нормальные волокна. Если разрушить определенную часть мозга и через несколько дней окрасить мозг методом Науты, а затем исследовать под микроскопом, то наличие избирательно окрашенных волокон в какой-либо другой и, возможно, даже отдаленной его части будет означать, что эта часть получает волокна от разрушенного участка. Такой метод привел к необычайному расширению и детализации карты мозга. За последнее десятилетие благодаря новейшим эффективным методам нейроанатомия продвинулась вперед больше, чем за предыдущие 50 лет (см. У. Наута и М. Фейртаг «Организация мозга»). Успехи достигнуты отчасти благодаря усовершенствованным химическим методикам и лучшему пониманию того, как различные вещества воспринимаются нейронами и передаются в обоих направлениях вдоль нервных волокон. Типичным примером может служить радиоавтография. Радиоактивное вещество вводится в ту или иную структуру мозга; тела клеток поглощают его, пересылают по своим аксонам, и оно накапливается в их окончаниях. Если затем приготовить срез ткани мозга, наложить его на фотоэмульсию и исследовать под микроскопом расположение проявленных зерен серебра, удается выявить «места назначения» аксонов. Можно вводить другие вещества, которые, наоборот, воспринимаются нервными окончаниями и передаются поаксонам в обратном направлении - к телу клетки, выявляя место возникновения аксона.
Окрашенная по Гольджи нервная ткань из зрительной коры крысы зарисована Рамон-и-Кахалом в 1888 г. Цифры по правому краю обозначают слои клеток; заглавными буквами помечены отдельные нейроны. Одним из самых важных вкладов Рамон-и-Кахала в нейробиологию явилось доказательство того факта, что нейрон представляет собой отдельную, обособленную клетку, а не элемент непрерывной сети. Последним в этом ряду достижений явилась методика, разработанная несколько лет назад Л. Соколовым (L. Sokoloff) в Национальном институте охраны психического здоровья. Глюкоза служит «топливом» для нейронов и в активном состоянии клетки потребляют больше глюкозы, чем в покое. Меченая дезоксиглюкоза усваивается клетками, как если бы это была глюкоза. Она расщепляется, как глюкоза, но продукт первого этапа ее метаболизма не подвергается дальнейшим превращениям. Не имея поэтому возможности выйти из клетки, этот продукт скапливается в ней, и степень радиоактивности в определенных клетках указывает на их функциональную активность. Можно поставить, например, такой опыт: ввести это вещество внутривенно лабораторному животному, а затем предъявить звуковой раздражитель; микроскопическое исследование мозга позволит выявить те его области, которые связаны со слухом. Совсем недавно разработана новая методика-позитронно-эмиссионная томография, которая позволяет обнаруживать с помощью наружных датчиков присутствие дезоксиглюкозы или других веществ, меченных радиоактивными изотопами, испускающими позитроны. Эта перспективная методика делает возможным картирование активных структур мозга in vivo у лабораторного животного или у человека. Применение всех существующих методик для выявления в первом приближении, без деталей, связей в одной только структуре — скажем, в части коры больших полушарий или в мозжечке — может занять у одного-двух анатомов пять или десять лет. Законченных нейроанатомов, людей особой породы, часто одержимых, а порой даже полупараноиков, во всем мире насчитывается всего несколько десятков. А поскольку мозг состоит из сотен разных структур, становится ясно, что одного только понимания связей в головном мозгу придется ждать еще много лет. Кроме того, изучить связи данной структуры в мозгу — это еще не значит понять ее физиологию. Для этого надо прежде всего узнать, как работают отдельные нейроны. За последние три-четыре десятилетия стало более или менее понятно, как один нейрон генерирует электрические сигналы и передает информацию другим клеткам. Над выяснением этого механизма трудились многие ученые, в их числе: Г. Дэйл (Н. Dale), О. Леви (О. Loewi), А. Ходжкин (A. Hodgkin), А. Хаксли (A. Huxley), Б. Катц (В. Katz), Дж. Экклс (J. Eccles) и С. Куффлер (St. Kuffler). Одно из удивительных открытий состояло в том, что все нейроны, несмотря на различия по величине и форме, используют два одинаковых вида электрических сигналов: градуальные потенциалы и потенциалы действия. Весь нейрон — его тело, длинный аксон и ветвящиеся дендриты — поляризован таким образом, что внутри он заряжен отрицательно приблизительно на 70 мВ по отношению к наружной поверхности. Этот «потенциал покоя» создается двумя свойствами клеточной мембраны. Во-первых, мембрана активно переносит ионы, выводя из клетки положительно заряженные ионы натрия и пропуская внутрь положительно заряженные ионы калия, вследствие чего концентрации этих двух видов ионов внутри клетки и снаружи совершенно различны. Во-вторых, способность проникать через мембрану у натрия и калия также различна. Изменения разности потенциалов, которая создается таким образом между наружной и внутренней сторонами клетки в состоянии покоя, -это электрические сигналы нервов. Изменение трансмембранного напряжения на любом участке клетки или ее отростков быстро распространяется по мембране во всех направлениях, постепенно затухая; уже на расстоянии нескольких миллиметров сигнал вряд ли удается обнаружить. Это электрический сигнал первого рода, т.е. градуальный потенциал. Его основная функция состоит в передаче сигналов на очень короткие расстояния. Сигнал второго типа, потенциал действия, передает информацию на большие расстояния. Если мембрана деполяризована (ее потенциал снижен) до критического уровня — от уровня покоя, равного 70 мВ, до приблизительно 50 мВ, - то наступает внезапное, резкое изменение: на время снимаются существующие препятствия току ионов калия и натрия и возникает локальный поток ионов, достаточный для того, чтобы изменить знак мембранного потенциала, который становится положительным внутри и достигает 50 мВ, а затем полярность снова меняется и восстанавливается нормальный потенциал покоя. Все это занимает около тысячной доли секунды. За это время первое изменение знака (на положительный внутри) создает мощный градуальный потенциал, который распространяется и доводит соседний участок мембраны до критического уровня; это вызывает изменение полярности в следующем участке мембраны, что в свою очередь создает изменение знака в соседнем участке. В результате временное изменение полярности быстро распространяется по нервному волокну. Этот распространяющийся потенциал действия, «бегущий» по всей длине волокна, не ослабевая, и является нервным импульсом. Любая сигнализация в нервной системе на расстоянии миллиметра и более происходит в форме импульсов. Независимо от типа волокна и от того, связано ли оно с движением, зрением или мышлением, сигналы везде практически одинаковы. Меняется же в данном нервном волокне при определенных условиях просто число импульсов в секунду. Приход импульса к окончанию аксона сказывается на следующем нейроне в цепи: изменяется его склонность генерировать импульсы. На пресинаптической мембране окончания выделяется медиатор - вещество, которое диффундирует через щель между двумя клетками и оказывает на постсинаптическую мембрану по ту сторону щели одно из двух воздействий. В возбудительномсинапсе медиатор вызывает понижение постсинаптического мембранного потенциала, и в результате постсинаптическая клетка генерирует импульсы с большей частотой. В тормозном синапсе эффект медиатора состоит в стабилизации постсинаптического мембранного потенциала, из-за чего возбудительным синапсам труднее деполяризовать постсинаптическую клетку, и вследствие этого генерация импульсов либо прекращается, либо идет с меньшей частотой. Является ли данный синапс возбудительным или тормозным, зависит от того, какой медиатор выделяет пресинаптическая клетка, и от химизма мембраны постсинаптической клетки. Почти к каждому нейрону приходят входы от множества окончаний, обычно от многих сотен, а иногда и тысяч, причем одни из них возбудительные, а другие тормозные. В каждый данный момент одни входы активны, а другие бездействуют, и от суммы возбудительных и тормозных эффектов зависит, будет ли клетка генерировать импульсы, и если будет, то с какой частотой. Иными словами,нейрон — это нечто гораздо большее, чем простой прибор для пересылки импульсов с одного места на другое. Каждый нейрон непрерывно оценивает все сигналы, приходящие к нему от других клеток, и выражает результаты этой оценки частотой своих собственных сигналов. Таким образом, распространение обоих типов сигналов по мембране нейрона и химические процессы на уровне синапсов в общих чертах изучены. Однако далеко не ясна связь между формойнейрона (форма дерева с разветвленной кроной) и способом суммирования и оценки получаемых сигналов. Два входных сигнала, из которых каждый может быть возбудительным или тормозным, несомненно суммируются совсем по-разному, в зависимости от того, расположены ли синапсы по соседству (например, на одной и той же дендритной веточке) или один синапснаходится на одной веточке, а другой далеко от него (возможно, на веточке другого дендрита) или же один синапс образуется на веточке, а другой на теле клетки. Форма, весьма различная у разных классов нейронов, должна играть важную роль в их функции, но пока это все, что можно сказать с уверенностью. Близкая к этому группа вопросов касается значения некоторых синапсов (обычного вида синапсов с пресинаптическим и постсинаптическим компонентами), которые соединяют два дендрита или два аксона, а не как обычно — аксон с дендритом или с телом клетки. Мягко выражаясь, никто толком не знает, что о них думать. Наконец, положение еще больше осложняется тем, что некоторые синапсы совершенно отличны от обычного химического типа, так как в них не происходит диффузия медиатора, а течет электрический ток. Их открыли в 50-х годах Э. Фершпен (Е. Furshpan) и Д. Поттер (D. Potter) из Лондонского университетского колледжа. Почему в одних синапсах природа применяет передачу химическую, а в других - электрическую, пока что остается загадкой. Если глубже рассматривать многие еще не получившие ответа вопросы относительно нервных сигналов, то они оказываются связанными с тонкой структурой и функцией нейронной мембраны, потому что на молекулярном уровне точно еще не известно, как ионы проходят через мембраны или как влияют изменения потенциала и медиаторы на проницаемость для отдельных ионов (см. Ч. Стивене «Нейрон»). Особый интерес представляет химизм синаптической передачи, при которой обнаружено уже больше 20 медиаторных веществ, а также те способы, какими нейроны синтезируют, выделяют, поглощают и разрушают эти уже достаточно хорошо известные вещества (см. Л. Иверсен «Химия мозга»). Еще не полное, но в значительной мере уже достигнутое понимание химии синапсов нашло свое отражение в психиатрии и фармакологии. Многие заболевания - от паркинсонизма до депрессии, - по-видимому, связаны с нарушением синаптической передачи, и действие многих лекарств сводится к усилению или ослаблению этой передачи (см. С. Кити «Заболевания человеческого мозга»). Лет через десять основные черты активности отдельных нейронов, надо полагать, будут детально изучены. Теперь же, когда положено известное начало пониманию структуры и работы отдельных клеток, нейробиологи находятся в положении человека, который, немного разбираясь в физических свойствах сопротивлений, конденсаторов и транзисторов, пытается заглянуть в телевизор. Он не поймет, как работает этот прибор в целом, пока не узнает, как соединены его элементы, и пока не получит хоть некоторое представление о назначении прибора, о его блоках и их взаимодействии. В изучении мозга первую ступень после нейрона и его работы составляет знакомство с тем, как взаимосвязаны более крупные части мозга и как устроена каждая из них. Следующей ступенью явится попытка установить, как нейроны взаимодействуют, и выяснить значение посылаемых ими сигналов. Один из способов разобраться в общей организации мозга состоит в том, чтобы рассмотреть ее в форме приблизительной схемы. На входе лежат группы рецепторов — видоизмененные нервные клетки, специализированные для преобразования в электрические сигналы разных форм информации, которая приходит к ним из внешнего мира. Одни рецепторы реагируют на свет (зрение), другие на химические воздействия (вкус и обоняние), третьи на механическую деформацию (осязание и слух). Рецепторы образуют контакты с первой группой нейронов, те в свою очередь со следующими, и так далее. На каждом этапе аксоны ветвятся и подходят к следующему нейрону цепи, на котором конвергирует множество аксонов. Каждая воспринимающая клетка интегрирует возбудительные и тормозные импульсы, конвергирующие на ней от клеток низшего порядка. Раньше или позже, после ряда этапов нервные аксоны оканчиваются на клетках железы или мышцы — выходах нервной системы. Коротко говоря, имеется вход — единственный для человека способ узнавать о внешнем мире. Имеется выход — единственный для человека способ реагировать на внешний мир и воздействовать на него. А между входом и выходом находится все остальное, что должно включать восприятие, эмоции, память, мысль и все прочее, что делает человека человеком. Вряд ли нужно говорить, что описанный путь от входа к выходу очень упрощен. Хотя главное движение происходит на схеме слева направо, но на любом этапе между клетками часто существуют боковые связи; нередко имеются также связи, идущие в обратном направлении — от выхода к входу, совершенно так же, как имеется обратная связь во многих электронных цепях. От входа к выходу ведет не один только путь; множество разных наборов рецепторов специализированы для разных видов чувствительности и для определенных форм энергии, действующих на каждый орган чувств, и при этом имеются бесчисленные шунты, переключения и обходы. Синапсов между рецепторами и мышцами может быть или очень много, или всего лишь два или три. (Когда число их невелико, цепь обычно называют рефлексом; сужение зрачка в ответ на освещение сетчатки представляет собой рефлекс с участием четырех или пяти синапсов.) При этом следует снова указать, что синапс бывает или возбудительный или тормозный; если в определенный момент клетка испытывает оба этих воздействия, в результате они могут полностью погасить друг друга.
Общая организация головного мозга показана в виде примерной схемы, на которой изображен поток информации от сенсорных сигналов на входе рецепторных клеток (А) до конечного выхода мотонейронов (Z) на мышечные клетки. На своем выходе рецепторы и нейроны обычно ветвятся и посылают к следующему уровню дивергирующие сигналы. Большинство нейронов получает на входе от нижних уровней конвергирующие сигналы как возбудительные, так и тормозные. Кое-что известно о значении связей близ входа (В, С) и близ выхода (X, Y) головного мозга. Гораздо меньше известно о работе областей лежащих между ними, которые составляют большую часть мозга. Теперь физиологи имеют некоторое представление о том, какого рода операции нервная система производит на входе и на выходе. На входе система, очевидно, занята главным образом извлечением из внешнего мира информации, которая представляет биологический интерес. Обычно рецепторы реагируют лучше всего на начало или на конец стимула, например давления на кожу. Нам важнее всего узнавать об изменениях; никому не хочется и не нужно, чтобы 16 часов в день ему напоминали, что на ногах у него обувь. Если в качестве примера взять зрительную систему, то здесь важны контрасты и движения, и на первых двух-трех этапах значительная часть цепей занята усилением эффектов контраста и движения. На последующих уровнях зрительной системы поведение клеток становится сложнее, но оно всегда упорядочено и, к счастью, имеет смысл в применении к восприятию. В настоящее время путь информации, идущей от светочувствительных клеток сетчатки, прослежен в головном мозгу до шестого-седьмого этапа, до роли коры больших полушарий, связанной со зрением (см.статью Д. Хьюбела и Т. Визелл). Хотя в настоящее время зрительная система является одной из лучше всего изученных частей мозга, нейробиологам еще далеко до понимания того, как происходит восприятие или опознание предметов. Правда, объем сведений, приобретенных за несколько лет, прошедших с момента создания микроэлектродов, позволяет думать, что такой отдел головного мозга, как кора больших полушарий, со временем будет доступен — во всяком случае в принципе — сравнительно простому объяснению. На выходе нервной системы механизм, каким нейрон посылает импульс к мышечному волокну, в последнее время стал в основном понятен, и уже более 50 лет нам известны те части головного мозга, которые управляют движением. Однако, что именно делают в действительности эти структуры, когда человек движется или собирается двигаться, все еще во многом неизвестно. Изучение идет медленно главным образом потому, что исследовать произвольные движения надо у бодрствующих и хорошо обученных животных, тогда как изучать сенсорные системы можно у наркотизированных животных. Одно из основных усилий направлено сейчас на то, чтобы проследить за ходом моторного импульса от мотонейрона обратно к таким структурам, как моторная кора и мозжечок, с целью узнать, как на решение совершить движение влияют разные сигналы, идущие от входа нервной системы (см. статью Э. Эвартса). На выходе важно не сокращение отдельного волокна, а координированное сокращение и одновременное расслабление многих мышц; например, при сжимании кисти в кулак или схватывании предмета недостаточно просто согнуть пальцы, сократив сгибатели в предплечье, - надо также сократить разгибатели, чтобы мышцы-сгибатели пальцев не согнули кисть. Это противодействующее разгибание кисти происходит автоматически, без размышлений (это можно проверить, сжав руку в кулак и почувствовав при этом, как сокращаются мышцы-разгибатели на наружной стороне предплечья). Очевидно, что команда сжать кисть в кулак, идущая от головного мозга, включает в себя импульсацию клеток, удаленных на несколько переключений от выхода нервной системы, — клеток,аксоны которых распределяются по разным мотонейронам и тормозным нейронам, снабжающим в свою очередь все мышцы, участвующие в выполнении команды. При других движениях действуют другие цепи, в которые могут быть включены те же мышцы, но в других комбинациях. Таким образом, не трудно представить себе ряд такого рода функций, осуществляемых нервной системой, особенно близ ее сенсорного и моторного кондов. Недостает сведений главным образом относительно обширной промежуточной области — лобная и теменная доли, лимбическая система, мозжечок и так далее, и так далее, — хотя об их анатомии известно немало. В некоторых случаях известна своего рода физиология основной схемы связей, например установлено, какими нейронами возбуждаются или тормозятся другие нейроны. Для мозжечка не только описана в деталях схема связей, но и ясно, какие синапсы возбудительные, а какие тормозные; для некоторых видов синапсов с достаточной степенью надежности химически идентифицированы медиаторы. Но как работает мозжечок, известно только самым неопределенным образом. Он несомненно участвует в регуляции движений, мышечного тонуса, равновесия, но как такие функции выполняются этой великолепно построенной, упорядоченной и фантастически сложной машиной, совершенно неизвестно.
На выходе нервная система активирует мышечное волокно по концевым аксонным веточкам мотонейрона. Эта микрофотография получена с помощью сканирующего электронного микроскопа Б. Риз и Т. Ризом из Национального института инсульта, неврологических и коммуникативных нарушений. Одна концевая веточка лежит (в некоторых местах она отодвинута) в бороздке мышечного волокна. Нервное волокно в значительной части окружено оболочкой, образуемой шванновской клеткой, тело которой видно слева внизу. Бороздку пересекают узкие складки постсинаптической мышечной мембраны. Каждая складка примыкает к «активной зоне» нервного волокна; когда к синапсу приходит импульс из этой зоны через «окна» в оболочке, образованной шванновской клеткой, выделяется химический медиатор ацетилхолин. Молекулы ацетилхолина активируют рецепторы на мышечной мембране, вызывая таким образом мышечное сокращение. По общему признанию, работать с мозжечком трудно; он образует водораздел между сенсорными и моторными процессами в совершенно незаполненной части приведенной на рисунке схемы. Виды сигналов, поступающих на его входы — специфическое значение импульсов, приходящих в мозжечок из коры больших полушарий, спинного мозга и так далее, — известны недостаточно;нейронные структуры, к которым направлены его выходы и которые в свою очередь связаны с мышцами, тоже мало изучены. По тем же причинам и другие части мозга в большинстве своем все еще мало понятны. Несмотря на методические успехи последних лет, крайне необходимы новые революционные методы. В качестве одного лишь примера можно указать, что сейчас не существует способа изучать сигналы отдельных клеток у человека, не вскрывая его черепа на операционном столе, последнее же совершенно недопустимо. Тем не менее достигнуты большие успехи в понимании некоторых высших функций человеческого мозга (см. Н. Гешвинд «Специализация человеческого мозга»), но чтобы действительно понять такую специально человеческую функцию, как речь, нужно разработать способы экстракраниальной регистрации активности отдельных нейронов. Указать, как соединены между собой нейроны и как они работают в каждый данный момент — это лишь одна из конечных целей нейрофизиологии. Некоторые важные аспекты деятельности мозга лежат за этой гранью. Например, память и обучение безусловно представляют собой кумулятивные процессы, связанные с изменением во времени, а о механизмах, лежащих в их основе, известно еще очень мало. Пожалуй, нейробиология особенно склонна к увлечениям, из-за которых порой она чуть не сходит с рельсов. Несколько лет тому назад возникла мысль, что следы памяти могут откладываться в форме крупных молекул, причем информация кодируется последовательностью мелких молекул подобно тому, как генетическая информация закодирована в ДНК. Мало кто, знакомый с высоко упорядоченной специфичностью связей в головном мозгу, принял эту идею всерьез, и тем не менее немало времени было потрачено во многих лабораториях на то, что животных обучали выполнению какой-либо задачи, затем растирали их мозг, а далее находили или химические отличия в их мозге, или «статистически значимое» усиление способности обучаться тем же задачам у животных, которым был введен экстракт мозга обученных животных. Увлечение это угасло, но факт тот, что не всегда нейробиология двигалась вперед или даже стояла на месте — временами она даже откатывалась назад. В конечном счете изучение памяти, вероятно, будет касаться двух совсем разных ее компонентов. Первый компонент составляют изменения, которые вероятнее всего происходят в синапсах в результате многократного использования нейронных цепей. Например, возможно повышение эффективности одного синапса за счет других на той же клетке. Определенные комбинации стимулов, повторяясь, могут таким образом усиливать один из многих проводящих путей данной нейронной структуры. Исследования такого плана трудно осуществимы на высших животных; их гораздо легче производить на малых системах нейронов, которые составляют полностью или частично нервную систему некоторых низших животных. В отдельные клетки этих животных удается легко ввести микроэлектрод, и, что еще важнее, такие клетки часто обладают индивидуальностью; говоря, например, о клетке №56 в определенном ганглии у рака, можно быть уверенным, что у всех других раков она занимает буквально то же положение и обладает теми же связями. (В этом глубокое различие между мозгом многих беспозвоночных и мозгом человека. Дать номер какому-нибудь нейрону в человеческом мозгу невозможно так же, как невозможно дать номер волосу на голове или поре на коже.) Изящные опыты проводились на уровне отдельных нейронов у беспозвоночных при их обучении (см. Э. Кэндел «Малые системы нейронов»). Например, показано даже, что при выработке реакции или ее забывании у животного происходят доступные определению изменения в передаче сигналов через те или иные синапсы. Разумеется, обучение здесь простое, но, по-видимому, истинное. Снова и снова на беспозвоночных делались открытия, которые потом были распространены высшие формы. Поэтому маловероятно, что исследователи откажутся работать на таких животных, убоявшись насмешек начетчиков над невероятным звучанием таких тем, как «Решение задачи пиявкой». Второй компонент, которым придется овладевать при изучении памяти, будет гораздо труднее. Все то, что человек помнит (в любом смысле этого слова, кроме самого рудиментарного), связано с восприятием или с движениями, или с жизненным опытом. Для проникновения в механизм памяти в любом истинном смысле нужно знать, что происходит, когда человек воспринимает, действует, думает и испытывает, для того чтобы понять, что из всего этого воспроизводится, когда он вспоминает что-то или чему-то научается. Из этих двух компонентов первый — синаптический — представляется мне сравнительно легким, а второй неимоверно трудным. Понять работу мозга у взрослого животного — задача достаточно трудная; понять, как мозг становится мозгом, пожалуй, по меньшей мере так же трудно. Как развивается нервная система до и после рождения? Центральная задача состоит в том, чтобы раскрыть, как информация, закодированная в молекулах ДНК, трансформируется в связи между клетками внутри структур, в пространственные соотношения этих структур и связи между ними. Зрительный нерв, например, содержит около миллиона волокон, каждое из которых начинается в определенной точке маленького участка сетчатки. В свою очередь каждое волокно соединено в определенном порядке с плоским латеральным коленчатым телом в головном мозгу, и таким образом на коленчатом теле создается в известном смысле карта сетчатки. Каким образом во время развития волокна выходят из сетчатки, доходят до коленчатого тела и распределяются там с абсолютной топографической точностью? Такие же наборы связей, подобных проводам, смонтированные по своим особым схемам и соединяющие между собой топографически организованные области, имеются по всей нервной системе, а как собираются эти точные схемы, остается одной из важных неразрешенных проблем (см. У. Коуэн «Развитие мозга»). Изучение развития мозга потенциально важно не только потому, что оно проливает свет на работу мозга, но и потому, что о многих нервных болезнях известно точно, а о многих предположительно, что они возникают в период развития. Сюда относится большинство врожденных аномалий, синдром Дауна, некоторые виды мышечной дистрофии, вероятно, некоторые обычные формы эпилепсии и множество более редких заболеваний. Остается только гадать о том, сколько пройдет времени, пока можно будет сказать, что мозг — или разум — в общем и целом понят (снова эти расплывчатые слова). Еще недавно, в 1950 г., того, кто предсказал бы, что через 10 лет процессы, лежащие в основе жизни, будут разгаданы, — сочли бы оптимистом, если не чудаком, а между тем, так и случилось. Думаю, что понадобится гораздо более 10 лет, чтобы понять мозг, просто потому, что он так многогранен, — это шкатулка, до краев наполненная остроумными решениями огромного множества задач. Весьма возможно, что человеческие существа никогда не разгадают всех тех отдельных задач, которые задает им мозг. Но можно надеяться, что по мере того, как будут рассмотрены одна за другой все области мозга, станет все яснее и яснее, что функции мозга упорядочены и доступны уразумению в понятиях физики и химии, без привлечения непознаваемых, сверхестественных процессов (см. Ф. Крик «Мысли о мозге»). На этом пути будут отдельные главные вехи. Например, может быть раскрыт некий единый механизм, посредством которого работает память (ее синаптический компонент), или некий единый процесс, который объясняет, как в процессе развития нервные волокна находят места свoero назначения. Однако это не значит, что в будущем в какой-то момент будет сделано открытие или ряд открытий, в результате которых мозг станет совершенно понятным. Исследования мозга прогрессируют медленно. Методические усовершенствования нескольких последних десятилетий заметно ускорили этот процесс, но, безусловно, не привели К каким-либо внезапным переворотам, подобно тем, какие были совершены Коперником, Ньютоном, Дарвином, Эйнштейном или Уотсоном и Криком. Каждая из таких революций отличалась тем, что переводила какой-нибудь очень важный раздел проводимых человеком исследований природы в область рационального и экспериментального анализа, уводя его от сверхестественного. Если Коперник показал, что Земля не является центром мироздания, а Галилей увидел в небе не ангелов, а звезды и планеты; если Дарвин установил, что человек состоит в родстве со всеми остальными живыми существами; если Эйнштейн ввел новые представления о времени и пространстве, о массе и энергии; если Уотсон и Крик показали, что биологическая наследственность объяснима в физических и химических понятиях, то наряду с этими открытиями, сужающими границы сверхъестественного, главное, что еще остается науке решать, — это, очевидно, проблема мозга и то, представляет ли он собой нечто большее, чем чрезвычайно сложную великолепную машину. Этот вопрос затрагивает само существо человека, и поэтому принципиальные изменения в наших взглядах на человеческий мозг не могут не оказывать глубокого влияния на наши взгляды на нас самих и на окружающий мир. Разумеется, такие достижения произведут значительное воздействие и на другие области исследования. Будут до некоторой степени захвачены те разделы философии, которые занимаются природой мышления и восприятия, а также, я полагаю, некоторые части психологии, которые пытаются получить ответы на подобные вопросы косвенным путем. Если будут открыты механизмы обучения и памяти, это отразится на всей области воспитания. В нейробиологии революция истинно коперниковских или дарвиновских масштабов во всяком случае не совершится одним ударом, возможно, никогда не совершится, а если произойдет, то постепенно, в течение многих десятилетий. С каждым ее этапом человеческие существа, несомненно, будут все ближе к пониманию самих себя. Ч. СТИВЕНС Нейрон Это отдельная нервная клетка, строительный блок мозга. Она передает нервные импульсы по единственному длинному волокну (аксону) и получает их по многочисленным коротким волокнам (дендритам) Нейроны, или нервные клетки, являются строительными блоками мозга. Хотя они имеют те же самые гены, то же самое общее строение и тот же самый биохимический аппарат, что и другие клетки, они обладают и уникальными особенностями, которые делают функцию мозга совершенно отличной от функции, скажем печени. Важными особенностями нейронов являются характерная форма, способность наружной мембраны генерировать нервные импульсы и наличие уникальной структуры, синапса, служащего для передачи информации от одного нейрона другому.
Нейрон зрительной коры кошки, представленный на микрофотографии, был окрашен путем инъекции фермента — пероксидазы хрена. Тела нейронов, образующие фон, докрашены фуксином. Все волокна, отходящие от тела клетки, являются дендритами, получающими информацию от других нейронов. Передающее информацию волокно, аксон, намного тоньше и его нелегко увидеть при таком увеличении. Самое толстое волокно, направленное вертикально вверх, называют апикальным дендритом; в данный срез попала лишь небольшая его часть. При данном увеличении полная длина апикального дендрита должна составлять около 75 см. (Его ход можно проследить по соседним срезам.) При регистрации активности этой клетки у живой кошки обнаружено, что она реагирует на границу светлое-темное, идущую под углом 60° к вертикали. Благодаря его форме этот нейрон называют пирамидной клеткой. Это один из двух наиболее многочисленных типов нейронов в коре млекопитающих. Микрофотография получена Ч. Джильбертом и Т. Визелем из Гарвардской медицинской школы. Полагают, что мозг человека состоит из 1011нейронов: это приблизительно столько же, сколько звезд в нашей Галактике. Не найдется и двух нейронов, одинаковых по виду. Несмотря на это, их формы обычно укладываются в небольшое число широких категорий, и большинству нейронов присущи определенные структурные особенности, позволяющие выделить три области клетки: клеточное тело, дендриты и аксон. Тело содержит ядро и биохимический аппарат синтеза ферментов и других молекул, необходимых для жизнедеятельности клетки. Обычно тело нейрона имеет приблизительно сферическую или пирамидальную форму. Дендриты представляют собой тонкие трубчатые выросты, которые многократно делятся и образуют ветвистое дерево вокруг тела клетки. Они создают ту основную физическую поверхность, на которую поступают идущие к данному нейрону сигналы. Аксон тянется далеко от тела клетки и служит той линией связи, по которой сигналы, генерируемые в теле данной клетки, могут передаваться на большие расстояния в другие части мозга и остальной нервной системы. Аксон отличается от дендритов как по строению, так и по свойствам своей наружной мембраны. Большинство аксонов длиннее и тоньше дендритов и имеет отличный от них характер ветвления: если отростки дендритов в основном группируются вокруг клеточного тела, то отростки аксонов располагаются на конце волокна, в том месте, где аксон взаимодействует с другими нейронами. Функционирование мозга связано с движением потоков информации по сложным цепям, состоящим из нейронных сетей. Информация передается от одной клетки к другой в специализированных местах контакта — синапсах. Типичный нейрон может иметь от 1000 до 10000 синапсов и получать информацию от 1000 других нейронов. Хотя в своем большинствесинапсы образуются между аксонами одной клетки и дендритами другой, существуют и иные типы синаптических контактов: между аксоном и аксоном, между дендритом и дендритом и междуаксоном телом клетки.
Типичный нейрон позвоночного способен передавать нервные импульсы на значительные расстояния. У изображенного здесь нейрона все части увеличены пропорционально. Импульсы возникают в теле клетки и распространяются вдоль аксона, который имеет одну или несколько ветвей. Этот аксон, который для удобства изображен в виде гармошки, в действительности имеет в длину 1 см. Бывают аксоны длиной более 1 м. Конечные ветви аксона образуют синапсы более чем на 1000 других нейронов. Большинство синапсов связывает аксонные окончания одногонейрона с дендритами, образующими «дерево» вокруг клеточного тела другого нейрона. Таким образом, дендриты, окружающие нейрон на этой схеме, могли бы получать входные сигналы от десятков, сотен и даже тысяч других нейронов. Многие аксоны, такие, как этот, снабжены изолирующей миелиновой оболочкой, прерывающейся через определенные интервалы участками, называемыми перехватами Ранвье.
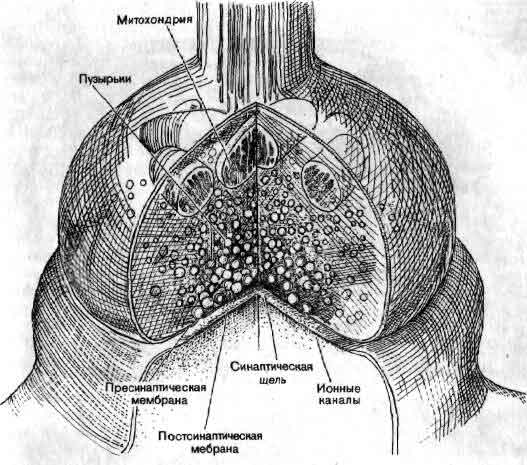
Тело нейрона содержит общий для всех клеток генетический материал и сложный метаболический аппарат. Однако в отличие от большинства других клеток нейроны после завершения эмбрионального периода не делятся; исходный их запас должен служить в течение всей жизни организма. От тела клетки отходит несколько дендритов и один аксон. Тело клетки и дендриты покрыты синапсами — бляшкообразными структурами, через которые поступает информация от других нейронов. Митохондрии снабжают клетку энергией. Белки синтезируются на эндоплазматическом ретикулуме. Транспортная система перемещает белки и другие вещества от тела клетки к тем местам, где они требуются. В области синапса аксон обычно расширяется, образуя на конце пре-синаптическую бляшку, которая является передающей информацию поверхностью контакта. Концевая бляшка содержит мелкие сферические образования, называемые синаптическими пузырьками, каждый из которых содержит несколько тысяч молекул химического медиатора. По прибытии в пресинаптическое окончание нервного импульса некоторые из пузырьков выбрасывают свое содержимое в узкую щель, отделяющую бляшку от мембраны дендрита другой клетки, предназначенного для приема таких химических сигналов. Таким образом, информация передается от одного нейрона другому с помощью некоторого посредника, или медиатора. Импульсация нейрона отражает активацию воздействующими нейронами сотен синапсов. Некоторые синапсы являются возбуждающими, т. е. они способствуют генерации импульсов, тогда как другие — тормозные — способны аннулировать действие сигналов, которые в их отсутствие могли бы возбудить разряд нейрона.
Синапс — это место переключения, в котором происходит передача информации от одного нейрона к другому с помощью химических медиаторов. Синапс состоит из двух частей: бляшкообразного утолщения, принадлежащего окончанию аксона, и рецепторной области на поверхности другого нейрона. Мембраны разделены синаптической щелью шириной около 200 нм. Молекулы химического медиатора, запасенные в пузырьках аксонного окончания, выделяются в щель под действием приходящих нервных импульсов. Медиатор изменяет электрическое состояние воспринимающего нейрона, увеличивая или уменьшая вероятность генерации этим нейроном импульса. Хотя нейроны и являются строительными блоками мозга, это не единственные клетки, которые в нем имеются. Так, кислород и питательные вещества поставляются плотной сетью кровеносных сосудов. Существует потребность и в соединительной ткани, особенно на поверхности мозга. Один из важных классов клеток центральной нервной системы составляют глиальные клетки, или глия. Глия занимает в нервной системе практически все пространство, которое не занято самими нейронами. Хотя функция глии пока не вполне изучена, по-видимому, она обеспечивает структурную и метаболическую опору для сети нейронов.
Синаптическое окончание занимает большую часть этой электронно-микроскопической фотографии, полученной Дж. Хойзером из Университета калифорнийской медицинской школы в Сан-Франциско и Т. Ризом из Национальных институтов здравоохранения. Щель, отделяющая пресинаптическую мембрану от постсинаптической, проходит вдоль нижней части фотографии. Крупные, темные структуры — это митохондрии; многочисленные округлые тела — пузырьки, содержащие медиатор; расплывчатые темные участки, расположенные вдоль щели, предположительно являются основными местами выделения медиатора. Еще одним типом клеток, повсеместно встречающихся в нервной системе, являются шванновские клетки. Оказывается, все аксоны заключены в оболочку из шванновских клеток. В некоторых случаях шванновские клетки просто окутывают аксон тонким слоем. Во многих же случаях в ходе эмбриогенеза шванновская клетка закручивается вокруг аксона, образуя несколько плотных слоев изоляции, называемой миелином. Миелиновая оболочка прерывается примерно через каждый миллиметр по длине аксона узкими щелями — так называемыми перехватами Ранвье. В аксонах, имеющих оболочку такого типа, распространение нервного импульса происходит путем его перескакивания от перехвата к перехвату, где внеклеточная жидкость оказывается в непосредственном контакте с клеточной мембраной. Эволюционный смысл миелиновой оболочки, по-видимому, состоит в экономии метаболической энергии нейрона. Как правило, миелинизированные нервные волокна проводят нервные импульсы быстрее, чем немиелинизированные. Дата добавления: 2016-06-06 | Просмотры: 2368 | Нарушение авторских прав |