|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
У. НАУТА, М. ФЕЙРТАГОрганизация мозга Головной и спинной мозг млекопитающих, включая человека, состоит из нескольких миллиардов нервных клеток, причем отдельные нейроны могут быть связаны с тысячами других. Как же организована эта огромная трехмерная сеть? Мы видим два общих подхода к представлению о нейроанатомии. Первый — высокопарный: утверждается, что мозг — вместилище ума, чувств и желаний, памяти и способности учиться, и того любопытного ощущения, которое свойственно людям, — ощущения будущего. Затем мы начинаем внимательно рассматривать, как этот таинственный орган выглядит, так сказать, «во плоти». Определенные части мозга, в особенности кора больших полушарий, удивительным образом организованы; другие поражают своей кажущейся неупорядоченностью. Но даже и самые высокоупорядоченные структуры, в которых организация нейронов и разнообразных связей между ними напоминает электронные схемы, сопротивляются нашим сегодняшним попыткам понять их.
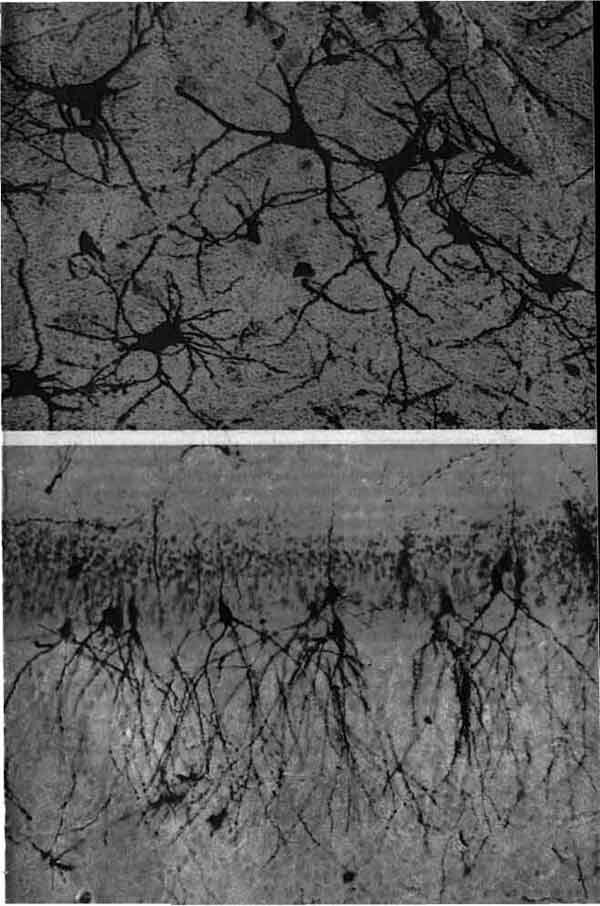
О порядке и беспорядке в клеточной организации мозговых структур можно судить по этим двум микрофотографиям. Обе они представляют тонкие срезы мозга кошки, которые подверглись двойному окрашиванию: по методу Гольджи, позволяющему выявить некоторые из нейронов со всеми их отростками в виде черных силуэтов, и по методу Ниссля, приводящему к окрашиванию всех клеточных тел в синий цвет. Метод Гольджи обеспечивает прокрашивание только 5 или менее процентов всех нейронов, по-видимому, по случайному выбору; если бы прокрашивались всенейроны, данная ткань выглядела бы однородно черной. А. Зубчатая извилина гиппокампа; тела нейронов в форме продолговатых пирамид располагаются в ряд, отсылая свои отростки почти параллельным пучком. Б. Микрофотография относится к крупноклеточной ретикулярной формации. Здесь нейроны образуют неупорядоченную сеть. Второй подход более прозаический. Мозг просто подразделяется на части, поскольку подходящие методы окрашивания выявляют в одних местах скопления нейронов, опутанных густой сетью их же собственных нитевидных отростков, в других же ткань состоит в основном из длинных Волокон нервных клеток — аксонов, которые обслуживают связи на далекие расстояния в нервной системе. Ткань первого рода — это серое вещество, второго — белое вещество. Конечно, есть искушение приписать каждой области какую-нибудь функцию, как если бы мозг в целом был чем-то вроде радиоприемника. Но существо работы центральной нервной системы - головного и спинного мозга — это проведение входящей сенсорной информации к множеству структур и конвергенция на нейронах, которые возбуждают эффекторные органы: мышцы и железы. Следовательно, система в целом обладает свойствами, выходящими за пределы тех, которые можно обнаружить у простого набора модулей. Возьмем, например, такую структуру мозга как субталамическое ядро. Его разрушение у человека ведет к моторной дисфункции, известной под названием гемибаллизма, при которой больной делает непроизвольные движения, как бы бросая мяч. Следует ли отсюда, что нормальной функцией субталамического ядра должно быть подавление движений, напоминающих бросание мяча? Конечно, нет; данное состояние больного только дает представление о работе центральной нервной системы, выведенной из равновесия отсутствием субталамического ядра. Мы упоминаем эти вещи, чтобы установить ограничения для любых объяснений анатомии мозга. Мы дадим здесь какое-то объяснение, но оно по необходимости будет несколько расплывчатым. Делать иного рода предположения было бы просто неправомерно. Полезно сделать некоторые предварительные замечания. В первые десятилетия этого века Дж. Паркер (G. Parker) из Йельского университета искал самую примитивную рефлекторную дугу. Такие дуги были идентифицированы у позвоночных животных; это состоящие из одного или нескольких нейронов пути, с помощью которых возбуждение, порождаемое сенсорным стимулом, действующим на некоторую часть тела, может быть проведено к эффекторной ткани и, соответственно, вызвать движение. Во времена Паркера рефлекторные дуги рассматривались как простейшие схемы, посредством которых природа объединяет клетки в нервную систему; в соответствии с этим было широко распространено мнение, что нервная система возникает тогда, когда некоторый организм приходит к необходимости иметь какую-нибудь клетку или цепочку клеток для посредничества между внешним стимулом и ответным движением животного. Со временем предполагалось установить, что у более развитых организмов эволюция нервной системы продвигалась вперед путем увеличения числа и сложности таких цепочек. Сначала внимание Паркера привлекли эпителиальные слои некоторых морских гидроидных полипов и актиний, потому что иногда они содержали клетки, которые выглядели (если были подходящим образом окрашены) как нейроны. У основания такой клетки Паркеру удалось разглядеть начало нити, весьма похожей на аксон, которая, подходя к мышечному волокну, разветвлялась. Паркер не был уверен в том, что тут образовывался контакт, но предположил, что между аксоном и мышечным волокном устанавливается какая-то связь. Конечно, он был прав, но это устройство весьма примитивно; его схему можно было бы назвать однонейронной нервной системой, поскольку вся линия проведения состоит всего лишь из одной клетки. Что такая нервная система будет делать в ответ на стимул, столь же предсказуемо, как и то, что будет делать дверной звонок, если нажать на кнопку. В то же время относительно нервной системы человека ясно, что поведение, на которое она делает способными человеческие существа, самое что ни на есть непредсказуемое. Очевидно, что-то должно вмешиваться в работу механизма дверного звонка, и потому Паркер исследовал ситуацию у несколько более сложных организмов. В эпителиальном слое у некоторых полипов и медуз он обнаружил нейронные структуры, сходные с найденными им ранее. Однако под эпителием он теперь нашел дополнительные нейроны, которые вместе образовали широко разветвляющуюся сеть. Таким образом, нервная система этой второй группы организмов оказалась усложненной: нейроны эпителиального слоя образовывали контакты с субэпителиальной сетью, а клетки этой сети в свою очередь образовывали контакты с сократимыми тканями в глубине организма. Следовательно, уже можно было говорить о двухнейронной нервной системе, в которой сенсорные нейроны (у этих простых созданий нейроны лежат вблизи поверхности и находятся в непосредственном контакте с окружающей средой) связаны с мотонейронами (нейронами, которые образуют контакты с эффектерными клетками, в данном случае — сократимыми клетками, и, значит, по существу, с мышечными волокнами). Остается ли эта сеть в высшей степени предсказуемой? По-видимому, нет. Предположим, что мотонейроны связаны друг с другом и на вход каждого из них поступают не только сообщения, идущие из окружающей среды через посредство сенсорных нейронов, но и сообщения от соседних мотонейронов. Вообразим далее, что некоторые из этих сообщений могут быть возбуждающими, т.е. заставляют данный мотонейрон с большей готовностью генерировать и передавать свои собственные сигналы в ответ на поступление других сигналов, и что другие сообщения могут быть тормозными. В таких условиях приходится решать вот какую задачу: чтобы предсказать, что будет делать некоторый нейрон в ответ на различные входные сигналы, нужно алгебраически просуммировать возбуждающие и тормозные сообщения, которые на нем конвергируют. Возможно, что такое двухнейронное устройство позволяет облагодетельствованной природой медузе быть более непредсказуемой в своем поведении, чем актинии и другие организмы с однонейронной нервной системой. Однако затем следует дальнейшее достижение, и оно тоже обнаружено у очень примитивных организмов — некоторых других меду:». В каком-то смысле это высшее достижение, так как и нервная система таких медуз, и нервная система чело века — обе состоят по существу только из трех классов нейронов. У этих медуз, как и у человека, сенсорныенейроны, как правило, больше не связаны непосредственно с мотонейронами. Между этими двумя элементами развивается барьер из нейронов, которые имеют взаимосвязи не только с мотонейронами, но и друг с другом. Правда, этот третий и последний шаг, возможно, уже был сделан всеми организмами, которые имеют субэпителиальную сеть нервных клеток. Выше, при рассмотрении двухнейронной нервной системы, предполагалось, что все клетки, составляющие этот слой, — мотонейроны, т.е. клетки, иннервирующие эффекторные ткани. Однако в действительности такие связи могут устанавливать только некоторые из многих субэпителиальных клеток. Остальные могут располагаться в сплетении таким образом, что получают на вход сингалы от сенсорных нейронов эпителия, а сообщаются лишь с другими нейронами того же типа или с мотонейронами, но не с эффекторными структурами. Не являясь ни сенсорными, ни моторными, они служат посредниками при проведении сигнала по сенсомоторному пути. Короче, здесь тоже имеются промежуточные (вставочные) нейроны. Хотя трехнейронную организацию трудно вычленить в диффузной нейронной сети, она широко представлена на более поздних стадиях эволюции; у животных, более высоко развитых, чем медузы, диффузная субэпителиальная нервная сеть концентрируется либо в последовательность сегментарных ганглиев (скоплений нейронов), либо в единую несегментированную центральную нервную систему. Таким образом, критическим моментом явилось изобретение «великой промежуточной сети» — барьера из промежуточных нейронов, которые вклинились между сенсорными нейронами и мотонейронами, на раннем этапе эволюции животных. Насколько далеко зашел прогресс в развитии этой промежуточной сети на сегодняшний день, легче всего показать с помощью некоторых цифр. Начнем с вопроса о том, сколько нейронов в центральной нервной системе человека. В качестве ответа часто можно услышать: порядка 1010. Это результат подсчета вставочных нейронов и мотонейронов, ибо случилось так, что настоящие сенсорные нейроны лежат не в центральной нервной системе, а в ганглиях, которые расположены вне головного и спинного мозга. Это весьма привлекательная цифра, которую легко запоминать и использовать. И все же имеются еще классы нейронов, которые столь малы и столь плотно упакованы, что оценить их число трудно или невозможно. Один из таких классов — это клетки-зерна. Как раз в одной из частей головного мозга — мозжечке — находится так много клеток-зерен, что величина 1010нейронов для всей центральной нервной системы становится сомнительной. Общее число вполне может быть на порядок, а возможно, и на два порядка больше. Все же примем на минуту, что общее число действительно 1010. Сколько же из этих клеток мотонейроны? Ответ таков, что их, оказывается, не может быть больше двух или трех миллионов. Эта цифра представляется смущающе малой в свете того факта, что только через мотонейроны работа нервной системы может выразиться в движении. Более того, такой ответ заставляет предполагать, что на мотонейроны должно конвергировать неправдоподобно большое число влияний; другими словами, это заставляет предполагать, что типичный мотонейрон должен образовывать синаптические связи с огромным числом аксонов, исходящих от равно огромного числа нейронов великой промежуточной сети. Полагают, что на поверхности типичного мотонейрона в спинном мозгу человека имеется что-то около 10000 синаптических контактов, из которых около 2000 - на клеточном теле и 8000 - надендритах - отростках, ветвящихся локально, в отличие от единственного аксона. Это не означает, что на данный нейрон посягают 10000 вставочных нейронов: когда такие нейроны устанавливают связи с какой-то клеткой, они имеют тенденцию образовывать множественные синаптические контакты. Однако все равно получается, что средний мотонейрон должен подвергаться массированному воздействию: число 1010 для нейронов центральной нервной системы подразумевает, что на каждый мотонейрон приходится от 3000 до 5000 нейронов великой промежуточной сети. Вот какое последнее заключение должно быть выведено из цифр, которые мы процитировали: весь головной и спинной мозг человека - это великая промежуточная сеть, за исключением явно немногих миллионов мотонейронов. А когда великая промежуточная сеть начинает включать 99,98 процента всех нейронов, составляющих центральную нервную систему, этот термин теряет большую часть своего смысла: он начинает отражать саму ту сложность, с которой каждый должен столкнуться при попытках постигнуть нервную систему. Этот термин остается полезным только как напоминание о том, что большая часть нейронов мозга не принадлежит ни к сенсорным, ни к моторным. Строго говоря, они вставлены между чисто сенсорной частью организма и чисто моторной его частью. Они являются компонентами вычислительной сети. Вторая группа предварительных замечаний касается общей анатомии центральной нервной системы. Отметим, в частности, что у всех видов позвоночных головной и спинной мозг сначала появляются у зародыша всего лишь в виде трубки толщиной в один клеточный слой. В передней части этой нервной трубки, которая в конечном итоге будет заключена в череп, вскоре появляются три утолщения — первичные мозговые пузыри. Это ромбэнцефалон, или задний мозг; мезэнцефалон, или средний мозг; и прозэнцефалон, или передний мозг. («Энцефалон» происходит от греческого «внутри головы».) Из этих трех первичных пузырей передний пузырь является самым продуктивным по числу образующихся отделов и дальнейшей дифференцировке. Основным событием его эмбрионального развития является образование камер с левой и правой стороны. Они превращаются в полушария большого мозга; эту часть мозга часто называют также конечным мозгом (телэнцефалон); у некоторых видов он имеет скромные размеры, у других - чрезвычайно большие. Между полушариями лежит непарный центральный отдел переднего мозга, от которого отпочковались полушария. Он имеет название промежуточный мозг, или диэнцефалон.
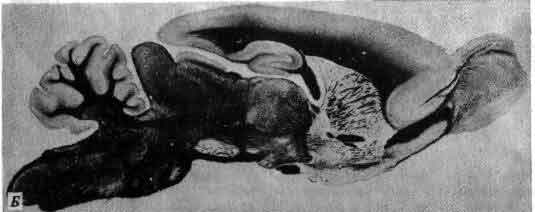
Эти два среза фиксированного препарата мозга человека иллюстрируют сложность внутреннего строения мозга. Срезы были обработаны краской, которая избирательно окрашивает в черный цвет жировую миелиновую оболочку нервных волокон; в результате белое вещество предстает как черное, а серое (состоящее преимущественно из клеточных тел) остается неокрашенным. Пустые пространства на этих срезах указывают положение желудочков—заполненных жидкостью полостей внутри мозга. Верхний рисунок. Срез проходит примерно посередине мозга и захватывает кору больших полушарий, гиппокамп и таламус. Нижний рисунок. Срез сдвинут кзади и проходит через ствол мозга. Как сделаны срезы и где располагаются упомянутые структуры, поясняет следующий рисунок. (Препарат из коллекции профессора П. Яковлева из Гарвардской медицинской школы.)
Мозг человека разрезан на пять частей и раскрыт как книжка, что позволяет проследить связи между наружным и внутренним строением. Два сечения, представленные на предыдущем рисунке, обозначены буквами А и Б. В процессе развития из переднего мозга выделяется еще одна пара камер-глазные пузыри. Их имеют даже незрячие животные, а у животных, способных видеть, они удлиняются в направлении к поверхности головы и в конечном счете превращаются в две сетчатки, соединенные с основанием переднего мозга своими стеблями-зрительными нервами. Наконец, из внутренней поверхности первичного переднего мозга развивается непарная срединная камера, которая дифференцируется и образует заднюю долю гипофизарного комплекса.
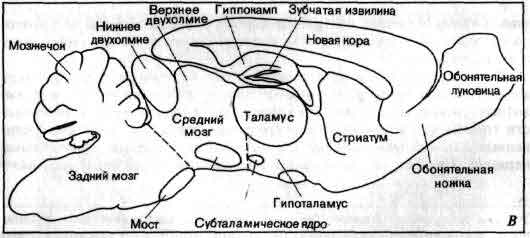
Два метода окрашивания дают возможность получить взаимно дополняющие друг друга изображения внутреннего строения мозга крысы. А. Показано сечение, проходящее чуть в стороне от срединной плоскости параллельно ей; срез был обработан по методу Ниссля, который позволяет избирательно окрашивать клеточные тела. Следовательно, каждая точка на микрофотографии соответствует какой-то отдельной клетке. Б. Срез обработан по методу Лойе (Loyez), который позволяет избирательно окрашивать миелинизированные волокна, оставляя тела клеток непрокрашенными, благодаря чему выделяются приводящие пути. В. Схема, на которой показаны различные структуры мозга. На нижнем рисунке на стр. 92 показан результат этого эмбрионального роста. Такая схема в общем выдерживается для всех млекопитающих; она изображает полностью сформированную центральную нервную систему млекопитающего, разбитую на несколько отделов. Слева - спинной мозг (на рисунке — сильно укороченный). Вправо от него, без какой-либо резкой границы, отходит задний мозг — самый нижний отдел головного мозга. Его дорсальная часть (часть, наиболее близкая к спине животного) - придаток, называемый мозжечком. За задним мозгом идет средний мозг, который у млекопитающих включает две пары структур, область из четырех бугров, известную под названием четверохолмия, крыши среднего мозга, или тектума («тектум» в переводе — «крыша»). Нижняя пара структур называется нижним двухолмием, верхняя пара — верхним двухолмием. Кроме этого, средний мозг дает мало оснований для подразделения, по крайней мере в продольном направлении. В сущности это довольно короткий участок человеческого мозга. Далее идет центральный непарный отдел переднего мозга — промежуточный мозг. Его дорсальные две трети — это таламус. Остальная часть — гипоталамус. (Несколько в стороне от гипоталамуса есть третий отдел промежуточного мозга — субталамус, самую удивительную группу клеток которого — субталамическое ядро — мы упоминали вначале. Включение его усложнило бы рисунок.) Гипоталамус характеризуется наличием железистого придатка, называемого гипофизарным комплексом. Он также продолжается в переднем направлении, переходя в перегородку — структуру, которую, несмотря на ее расположение, лучше всего классифицировать как принадлежащую к промежуточному мозгу. Остальная часть переднего мозга — конечный мозг, мозговые полушария. В мозгу млекопитающих это наибольшая часть, намного превосходящая другие, и у многих видов млекопитающих оболочка полушарий — мозговой плащ, или кора большого мозга, — испещрена извилинами (gyri) и бороздами (sulci). У основания каждого полушария имеется простирающаяся вперед структура, которая полностью состоит из серого вещества (коры), хотя это и кора с очень примитивной клеточной структурой. Ее вздутый передний конец — обонятельная луковица, а ее ножка — обонятельный стебель; только та часть коры, которая лежит непосредственно под остальной частью полушария, является настоящей обонятельной корой. Вторая большая часть коры мозга у млекопитающих находится там, где мозговой плащ подворачивается и образует сложные складки, которые в поперечном сечении напоминают орнамент в стиле рококо. Эта замечательная структура известна под названием гиппокампа.
Головной и спинной мозг человека и других млекопитающих можно подразделить на отделы, исходя из данных анатомии, эмбриологии и клеточной организации. А. Мозг человека изображен таким образом, что его внутренние структуры просматриваются сквозь «прозрачные» наружные слои мозжечка. Б. Обобщенная схема мозга млекопитающего; эта схема условно принята для всех остальных рисунков данной статьи. Соответствующие структуры на обоих рисунках окрашены одинаково. Обычно принято делить мозг на передний, средний и задний. Задний мозг включает мозжечок. К среднему мозгу относятся бугорки четверохолмия - нижнее и верхнее двухолмия. Строение переднего мозга более сложно. Его наружная часть - полушарие, поверхность которого - складчатый пласт мозговой коры, включающей гиппокамп, новую кору и обонятельные поля. Внутри полушария располагаются миндалина и полосатое тело; последнее подразделяется на бледный шар и стриатум, включающий хвостатое ядро и чечевицеобразное ядро. Остальная часть переднего мозга — промежуточный мозг: верхние его две трети составляет таламус (который имеет множество подразделении), а нижнюю треть - гипоталамус (который соединяется с гипофизарным комплексом). После выделения перечисленных частей остается еще одна часть коры большого мозга млекопитающих, которая имеет большую протяженность и чрезвычайно сложное строение; у человека и других приматов она, согласно оценкам, содержит не менее 70 процентов всех нейронов центральной нервной системы. Это новая кора — самая поздняя кора в процессе эволюции. Мы приобрели ее благодаря расхождению путей эволюции: после рептилий одна линия животных развивалась, усовершенствуя прежний тип, и дошла до птиц, а другая — более «авантюрная» - приобрела новую кору и дошла до млекопитающих. Таким образом, с чисто филогенетической точки зрения птицы воплощают в себе логическое завершение традиционного развития мозга, а млекопитающие представляют собой отклонение, поскольку они не имеют в своей родословной птиц. В результате одного из многочисленных разветвлений эволюции млекопитающих появились приматы - отряд, в котором новая кора достигает своего максимального развития. Мы, человеческие существа, пожинаем все плоды этого развития, возможно, включая и психопатологию. В глубине каждого полушария большого мозга млекопитающих имеется несколько скоплений серого вещества. Одно из них — это миндалина, которая лежит под обонятельной корой. Другое - полосатое тело, находящееся в самой середине полушария. Это тело в свою очередь подразделяется на две части, которые различаются по клеточному составу. Первая из них — это внутренняя зона, называемая палеостриатумом, или бледным шаром. Вторая зона - наружная; она известна под названием стриатум. Вернемся теперь к схеме связей в центральной нервной системе млекопитающих. Начнем с идентификации сенсорных нейронов, типа тех, которые Паркер нашел в эпителиальном слое медуз. Однако у позвоночных расположение сенсорных нейронов совсем иное. Известен всего один пример, где сенсорный нейрон является также рецептором на поверхности тела: только обонятельные эпителиальные клетки в слизистой крыши носа контактируют с внешней средой. Все другие сенсорные нейроны в теле позвоночных надежно упрятаны под поверхность и находятся в ганглиях, располагающихся вдоль спинного мозга по всей его длине, или в аналогичных ганглиях, лежащих в стороне от мозга. (У позвоночных термин «ганглий» сохранен для скоплений нейронов вне центральной нервной системы.) У каждого сенсорного нейрона есть аксон, который делится на две части: одна часть идет в центральную нервную систему, а другая иннервирует периферические структуры.
Соматосенсорная информация, например сообщения о кожных ощущениях, передается по нескольким путям спинного мозга. В левой части схемы показана пара репрезентативных сенсорныхнейронов, доставляющих сигналы от периферических сенсорных рецепторов в спинной мозг. Отсюда один путь сразу же ответвляется к мотонейронам (сплошные треугольники), волокна которых идут за пределы мозга к скелетным мышцам. Все другие пути сначала идут в скопления вторичных сенсорных нейронов, располагающиеся либо в той же самой области спинного мозга, либо в его верхней части, в ядрах дорсального столба. Путь, называемый медиальным лемниском, поднимается вверх от ядер дорсального столба к вентральному ядру таламуса, которое в свою очередь посылает волокна в соматосенсорную зону коры. Второй путь, называемый спино-таламическим трактом, поднимается к переднему мозгу из скоплений вторичных сенсорных клеток по всей длине спинного мозга, по ходу раздавая в стороны часть своих волокон. Малая доля волокон спино-таламического тракта в конечном счете достигает вентрального ядра. Скопления вторичных сенсорных клеток также посылают свои волокна в мозжечок. На приведенном здесь рисунке, одна из таких клеток — назовем ее первичным сенсорным нейроном — посылает свой аксон в спинной мозг, доставляя туда сенсорные сообщения о таких событиях, как прикосновение к коже, движение сустава или сокращение мышцы. Эти сообщения не сразу поступают на мотонейроны; свои первые синаптические контакты сенсорный нейронустанавливает с так называемыми вставочными нейронами. Однако имеется одно исключение. Это моносинаптическая рефлекторная дуга, образуемая боковой ветвью первичного сенсорного волокна, идущей «напролом» и устанавливающей прямой синаптический контакт с мотонейроном. Сначала это кажется обескураживающим: всего лишь несколькими абзацами выше мы отмечали, что после самых ранних ступеней эволюции нервной системы мотонейроны больше не утруждают себя обработкой сырых данных. Мы предположили, что вместо этого они теперь получают краткий итог обработки информации («дайджест»)нейронами великой промежуточной сети. Следовательно, моносинаптическая дуга может рассматриваться как очень примитивный тип нервной сети; с другой стороны, ее можно считать относительно новой: по-видимому, ее вновь открыли только наземные животные. В конце концов, воздух и земля — самые суровые среды; для горного козла один неверный шаг может оказаться роковым. В противоположность этому рыба может без всякого риска для себя сделать любое количество аналогичных ошибочных движений. Рыбы прекрасно уравновешены в воде, и сила тяжести для них далеко не так обременительна и враждебна. Таким образом, именно наземное, а не водное существование, по-видимому, требует наличия высоконадежных рефлекторных систем для удержания равновесия и, в частности, наличия пути, по которому мышца может сигнализировать соответствующим мотонейронам (и только им), что она чрезмерно растянута силой тяжести. Моносинаптические рефлекторные дуги никогда не обнаруживаются за пределами таксах корректирующих механизмов. Так что короткие цепи между сенсорным входом и моторным выходом оказываются представленными в ничтожно малом числе. Подавляющее большинство первичных сенсорных волокон млекопитающих входит в великую промежуточную сеть и синаптически контактирует с клетками, составляющими группу, которую мы будем называть группой вторичных сенсорных клеток: это нейроны, стоящие первыми в цепи приема первичных сенсорных сигналов. Оттуда многие пути более или менее прямо направляются к мотонейронам. Все они в совокупности могут быть названы локальным рефлекторным путем, если не упускать из виду, что слово «локальный» может ввести в заблуждение, так как есть несколько рефлексов, которые вовлекают в работу весь спинной мозг по всей его длине, но тем не менее относятся к локальным, поскольку не выходят за его пределы. Первое звено в локальном рефлекторном пути — это некоторый нейрон из группы вторичных сенсорных клеток. Многие из таких клеток сами не образуют контактов с мотонейронами; вместо этого они синаптически переключаются на некоторые другие нейроны великой промежуточной сети, и только уже эти последние нейроны, наконец, замыкают дугу. Другие пути состоят из аксонов, не идущих к мотонейронам. Возьмем мозжечковые каналы: из скоплений вторичных сенсорных нейронов заднего и спинного мозга многие аксоны восходят прямо к мозжечку. Показанный на рисунке на стр. 94 аксон, который идет таким образом, берет свое начало в группе вторичных сенсорных клеток спинного мозга и потому называется спино-мозжечковым волокном. (Слова «аксон» и «волокно» в нейроанатомии используются как синонимы.) Третий путь — это лемнисковый путь. Слово «лемниск» в переводе с латинского — лента (петля); здесь им обозначаются пучки волокон, которые берут начало в группах вторичных сенсорных клеток и поднимаются к переднему мозгу, в частности к таламусу. На рисунке (стр. 94) показано, как один такой пучок идет по центру спинного мозга. В действительности он проходитлатеральнее: упрощенную иллюстративную схему трудно сделать топографически точной. Этот пучок назван спино-таламическим трактом, хотя только одно из трех его репрезентативных волокон изображено как оканчивающееся в таламусе. Два других волокна сопровождают его на некотором расстоянии, а затем, так сказать, «совершают непредвиденную посадку»: на рисунке показано, что оба оканчиваются на нейронах заднего мозга, хотя с таким же успехом они могли бы окончиться несколько далее по ходу — в среднем мозгу. Важно то, что из волокон спино-таламического тракта лишь небольшая доля действительно достигает таламуса. Тем не менее тракт назван в честь этого преуспевшего меньшинства, которое оканчивается в специфической части таламуса — вентральном ядре. Здесь волокна образуют синапсы с таламическими нейронами, аксоны которых идут, не прерываясь, в специализированное поле новой коры, называемое соматосенсорной корой. Заметим, что на пути от первичного сенсорного нейрона до новой коры в данном случае насчитывается только два синаптических переключения. Первое — в спинном мозгу между первичным сенсорным волокном и нейроном из группы вторичных сенсорных клеток. Второе переключение — в промежуточном мозгу, между волокнами лемниска и нейронами вентрального ядра таламуса. Однако то, что происходит в новой коре, можно назвать синаптическим катаклизмом. В новой коре в реакцию на пришедший сигнал уже на первом этапе вовлекаются сотни, а то и тысячи нейронов. А действуя через синаптические связи, эти первые нейроны, возбужденные сигналом, вовлекают бесчисленное множество следующих. Двухсинаптическая сенсорная линия проведения, идущая в новую кору, может быть названа «сквозной линией», так как два синапса — это, по-видимому, минимум для такого рода систем. Она также может быть названа закрытой, или маркированной линией, поскольку, как правило, сенсорные пути с минимальным числом переключений сохраняют топографию той сенсорной периферии, от которой они идут. Так, кончик пальца способен различить два отдельных стимула, когда к нему прикасаются ножками измерительного циркуля в точках, находящихся всего лишь в двух или трех миллиметрах друг от друга. Это называется разрешающей способностью. Наличие такой способности означает, что каждая ножка циркуля стимулирует проводящие пути, которые достаточно независимы для того, чтобы обеспечить это, если можно так сказать, сенсорное разрешение. Какая-нибудь клетка соматосенсорной коры, если ее «допросить» с помощью микроэлектрода, может «признаться» в том, что ее единственный интерес составляет квадратный миллиметр кожи на указательном пальце. Одна из ее ближайших соседок может контролировать смежный квадратный миллиметр, и так далее. Благодаря этому топография поверхности тела будет верно воспроизведена. Проводящие пути, диаметрально противоположные маркированным,- это такие, по которым проводятся топографически перепуганные сообщения от данного сенсориума или даже от нескольких различных сенсориумов. Такая любопытная организация действительно существует: один из спино-таламических «отщепенцев», показанных на рисунке (стр. 94), синаптически переключается на нейрон заднего мозга, аксон которого продолжает путь в таламус. Однако на уровне этого «лишнего» переключения линия принимает сообщения не только от спино-таламического волокна, но и от слуховой системы. Как же таламус может понять, что случилось, когда импульсы поступают по этой системе путей? Нейроны заднего мозга называют мультимодальными, или неспецифическими, а рассмотренный выше проводящий путь можно назвать открытым: везде, где есть синаптические переключения, линия открыта для входа сигналов от других нейронов. Огромное большинство нейронов в глубине заднего и среднего мозга относится к нейронам именно такой любопытной неспецифической природы. Кажется, что они сидят, раскинув свои дендриты, словно длинные руки, на расстояния в несколько миллиметров, как бы стараясь поймать сигналы любого рода. Эти нейроны типичны для так называемой ретикулярной формации, в которой лишь сравнительна малое число клеточных скоплений получает гомогенные входные сигналы. Если бы эту ситуацию описали инженеру-электронщику, он, вероятно, отнесся бы к ней неодобрительно; он сказал бы, что от такой схемы нечего надеяться получить что-либо, кроме шума. Однако такая ситуация преобладает в мозгу всех позвоночных, включая человека. Так что ее существование, по-видимому, вызвано необходимостью удовлетворять какие-то специфические потребности. На данный момент можно сказать, что в функции ретикулярной формации входит обеспечение фона общей активации в центральной нервной системе и что ретикулярная формация воплощает в себе некий механизм регуляции состояний активности по всей центральной нервной системе. Некоторые из этих состояний подвержены суточным колебаниям — скажем сон и бодрствование. В каждом из этих состояний можно выделить большое количество «полутонов» возбуждения и расслабления. Все они связаны с тем или иным характером активности ретикулярной формации. Электронщика наверняка больше бы устроил второй соматосенсорный лемниск, выходящий из спинного мозга. Это медиальный лемниск. Он намного более компактно организован: почти все его волокна - маркированные линии, восходящие прямо к центральному ядру таламуса из двух скоплений вторичных сенсорных клеток на переходе между спинным и задним мозгом, которые называются ядрами дорсального столба. Наш инженер не удивился бы, узнав, что медиальный лемниск обеспечивает лучшее разрешение двух точек, чем спино-таламический тракт. А что сказать о других сенсориумах? Маленький орган, который для упрощения чертежа изображен на рисунке (стр. 99) возле мозжечка, - это орган слуха. Внутри него, в высоко специализированном эпителиальном комплексе, называемом кортиевым органом, обнаружены клетки, которые имеют по одной-единственной ресничке. Они иннервируются первичными сенсорными нейронами, у которых направляющиеся в центр отростки оканчиваются на нейронах кохлеарных ядер - скоплений вторичных сенсорных нейронов в заднем мозге, специализированных для приема и обработки сигналов исключительно от слухового сенсориума. На рисунке показаны только два нейрона; в действительности их десятки тысяч. В кохлеарных ядрах берут свое начало волокна латерального лемниска, восходящие к таламусу. Ни одно из них не выходит за пределы нижнего двухолмия. На этом пути при переключении в среднем мозгу начинаются аксоны, которые добираются до таламуса, где они заканчиваются в медиальном коленчатом теле. (На рисунке не показано несколько других слуховых «станций», по-видимому, менее обязательных и связанных с самим латеральным лемниском.) Нейроны медиального коленчатого тела в свою очередь проецируются (посылают свои аксоны) в специальную область новой коры, называемую слуховой корой. Сравним это со зрительной системой. Множество нейронов сетчатки обрабатывает выходные сигналы фоторецепторного аппарата глаза. Аксоны определенной части этих клеток сначала объединяются в зрительный нерв. Затем происходит перераспределение аксонов по двум путям, в процессе которого аксоны, вышедшие из назальной (носовой) половины сетчатки одного глаза, пересекают срединную плоскость головы, чтобы объединиться с аксонами, которые вышли из темпоральной (височной) половины сетчатки другого глаза. В результате образуются зрительные тракты. Аксоны, составляющие зрительный тракт, распределяются между двумя большими конечными областями. Одна из них — это верхнее двухолмие; однако у всех приматов более важной областью, по крайней мере если судить по количеству аксонов, является латеральное коленчатое тело таламуса. Нейроны этого ядра проецируются в свою очередь в новую кору, а именно - в определенную зону у заднего полюса полушария мозга, известную под названием зрительной коры. Отметим, что в противоположность слуховым волокнам, ни одно из которых не может достичь своего таламического ядра без синаптических переключений, большая часть зрительных волокон (у приматов) это делает. Следовало бы добавить, однако, что многие из нейронов верхнего двухолмия, получающих зрительные волокна, посылают свои собственные аксоны в таламус, но не влатеральное коленчатое тело, а в латеральное заднее ядро. Нейроны этого последнего клеточного скопления в свою очередь проецируются в новую кору, однако не в ту область, в которую идутаксоны из латерального коленчатого тела, а в соседнюю, которая отличается от первичной зрительной коры. По-видимому, зрительная система имеет два пути, восходящих к зрительной коре.
Три сенсориума (слуховой, зрительный и обонятельный) посылают свою информацию в кору различными путями. Слуховые пути, прежде чем они достигнут слуховой зоны коры больших полушарий, последовательно проходят через кохлеарные ядра заднего мозга, нижнее двухолмие среднего мозга и медиальное коленчатое тело таламуса. Зрительные пути начинаются в сетчатке (которая в действительности является частью мозга) и затем расходятся по двум различным каналам: один идет через латеральное коленчатое тело таламуса в зрительную кору, другой направляется через верхнее двухолмие среднего мозга в таламус, а затем — в область коры, расположенную около первичной зрительной зоны. В обонятельной системе рецепторные нейроны слизистой оболочки носа проецируются, минуя таламус, в обонятельную луковицу, которая является частью коры больших полушарий, а обонятельная луковица в свою очередь проецируется в обонятельную кору. Обонятельная система нарушает все законы, которым, казалось бы, подчиняется структура других сенсорных механизмов. Это, как мы отмечали, единственная из известных систем, в которой сенсорные нейроны лежат на поверхности. Там нет посредника - передающего элемента, как, скажем, в кортиевом органе; обонятельные эпителиальные клетки сами воспринимают внешние воздействия. Отходящие от таких нейронов очень тонкие аксоны проецируются в обонятельную луковицу, нейроны которой дают начало аксонам, заканчивающимся синаптическими контактами на клетках обонятельной коры. Мы только что проследили волокна четырех сенсориумов - соматического, слухового, зрительного и обонятельного. Теперь уже начинают выявляться некоторые существенные моменты. Во-первых, таламус предстает как узловая станция, конечный контрольно-пропускной пункт, дающий разрешение на вход в высшие инстанции мозга сигналам от всех сенсориумов (за исключением, по-видимому, обонятельного). Каждое такое переключение хочется сравнить с передачей эстафеты или с ретрансляционной станцией, однако то, что происходит в местах разрыванейронной цепи, может быть намного сложнее того, что происходит в эстафетном беге, где каждый бегун передает эстафетную палочку следующему и на конечный пункт маршрута эта палочка прибывает в неизменном виде. В центральной нервной системе «эстафета» совершенно иного рода. На каждом синаптическом переключении сенсорного пути входные сигналы трансформируются: код, с помощью которого записано прибывшее сообщение, коренным образом меняется. Можно предположить, что исходные данные не могут быть интерпретированы на высших уровнях; по-видимому, необходим их перевод на другой язык, и о синаптических ретрансляционных станциях лучше говорить как о станциях переработки данных. Затем можно сделать заключение, что кора является конечной станцией сенсорных проводящих путей. Нейроанатом бывает в высшей степени удовлетворен, когда ему удается проследить зрительные пути, скажем, от сетчатки до латерального коленчатого тела и оттуда — до зрительной коры. Трудность любого дальнейшего прослеживания заключается в сложности коры мозга, содержащей в себе 70 процентов всех нейронов центральной нервной системы человека. Что они делают со своими входными сигналами? Тут уместно привести следующие два наблюдения. Во-первых, таламо-кортикальные проекции имеют реципрокный характер: зрительная кора проецируется обратно на латеральное коленчатое тело, из которого в нее поступают входные сигналы; слуховая кора проецируется обратно на медиальное коленчатое тело, а соматосенсорная кора - на вентральное ядро. Такие возвратные связи, без сомнения, говорят о том, что функциональное состояние коры может влиять на ту процедуру, с помощью которой сенсорные станции таламуса «просеивают» направляемый в кору поток информации. Во-вторых, зрительная, слуховая и соматосенсорная зоны коры воплощают в себе только первые этапы обработки сенсорной информации. Из этих первичных сенсорных полей выходят волокна, синаптически воздействующие на смежные области, которые без оговорки нельзя назвать сенсорными: они представляют, так сказать, следующий этап обработки входных сигналов. А из этих областей выходят волокна, которые оканчиваются в областях, еще более далеких от первичных сенсорных полей. Области новой коры, более или менее удаленные от первичных полей, называются ассоциативными зонами, и у человека они занимают подавляющую часть поверхности коры; зрительная, слуховая и соматосенсорная зоны вместе занимают только около четверти всей ее площади. По всей вероятности, в ассоциативных зонах информация подвергается дальнейшей, более сложной переработке. Так, есть места, где слуховые и зрительные сигналы сходятся. Теперь известно, что в процесс обработки нервных сигналов в новой коре обычно последовательно вовлекается целый ряд ассоциативных областей и что местом назначения выходящей информации, по-видимому, неизменно являются либо гиппокамп, либо миндалина, либо и то и другое. В 1870 г. Г. Фрич (G. Fritsch) и Э. Гитциг (Е. Hitzig) опубликовали сообщение о том, что электрический ток минимальной силы, пропускаемый через область новой коры, расположенную фронтально по отношению к центральной борозде, может вызвать подергивание скелетных (но не висцеральных) мышц на стороне тела, контралатеральной месту стимуляции. Так, часто приходит в движение рука или нога. Это открытие, возможно, являющееся первым указанием на наличие в коре большого мозга функциональных подразделений, возбудило стойкий интерес к организации тех частей мозга, которые связаны с эффекторными (или моторными) функциями. В конце концов была открыта моторная кора — ограниченная область на самом высоком уровне мозга, которая явно причастна к движениям тела. Казалось, что теперь чисто моторную функцию можно будет, так сказать, вычленить из всего головного и спинного мозга. Отсюда начинаются поиски «моторной системы». Этот неопределенный термин включает не только мотонейроны, управляющие скелетной мускулатурой, но и нервные пути, которые сходятся на мотонейронах. Поиски эти длятся и по сей день, и резонно спросить, могут ли они когда-нибудь быть завершены? Рассмотрим поле 19 — полоску новой коры, отличающуюся по цитоархитектонике от соседних зон и расположенную недалеко от зрительной коры. Когда поле 19 стимулируют электрически у какого-нибудь экспериментального животного, оба глаза этого животного синхронно поворачиваются к противоположной стороне, т.е. взор отворачивается от раздражаемой током половины. В связи с этим появляется искушение назвать поле 19 моторной зоной. Однако сделать это — значит поступить произвольно, поскольку с другой точки зрения поле 19 — сенсорное: известно, что оно повторно обрабатывает информацию, которая прошла через зрительную кору. Сходный пример можно подобрать и для слухового сенсориума: вблизи соответствующей слуховой коры имеется область, называемая полем 22, электрическая стимуляция которой тоже заставляет животное поворачивать глаза к контралатеральной стороне. И при этом поле 22 состоит со слуховой корой в синаптических отношениях, во многом напоминающих отношения между полем 19 и зрительной корой. Урок из всего этого такой: между сенсорной и моторной структурами мозга нельзя провести разделяющую грань. Представляя ситуацию с другой стороны, можно сказать, что все нервные структуры занимаются программированием и регулированием поведения организма. По существу, это и есть функция нервной системы и эволюция разума содействовала ее развитию. Правда, некоторые структуры имеют в пределах великой промежуточной сети такое расположение, которое склоняет к определению их как сенсорных; примером тому служит латеральное коленчатое тело таламуса. А к другим структурам, расположенным на расстоянии не слишком большого числа синаптических переключений от мотонейронов, есть искушение приклеить ярлык «моторные». Это, впрочем, единственный способ разумного использования данных терминов. Соответственно при исследовании моторных функций центральной нервной системы, может быть, лучше всего начинать с уровня мотонейронов, которые однозначно являются частью моторной системы согласно любому определению, а затем пытаться проследить в мозгу те пути, которые воздействуют на них. Нужно только отдавать себе отчет в том, что применение этой стратегии означает движение вверх по течению, против превалирующего направления нервных потоков.
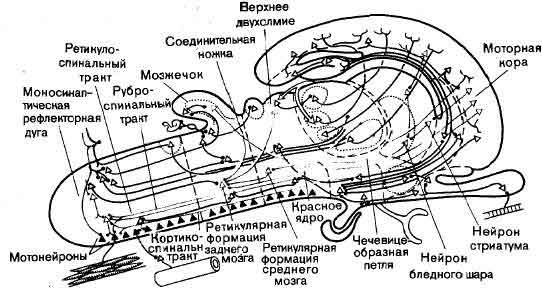
Мотонейроны головного и спинного мозга (сплошные треугольники) получают информацию из многих конвергирующих к ним каналов. Здесь показаны мотонейроны, на вход которых поступают сигналы от первичных сенсорных нейронов, из скоплений вторичных сенсорных нейронов в спинном мозгу, из ретикулярной формации ствола мозга, из красного ядра среднего мозга и из моторной коры переднего мозга. Красное ядро и ретикулярная формация сами получают сигналы из различных источников. Один из примеров конвергенции сигналов на мотонейронах является особенно впечатляющим: вся новая кора, включая слуховую, зрительную, соматосенсорную, а также моторные и другие поля, направляет пучки волокон в полосатое тело. Это ядро в свою очередь посылает массивные пучки волокон в ретикулярную формацию, которая, в конце концов, воздействует на мотонейроны. Второй путь, идущий из полосатого тела, играет роль петли обратной связи: составляющие его волокна направляются в область таламуса, которая проецируется обратно, опять же в моторную кору. Первый шаг «вверх» от мотонейронов, как правило, короток, поскольку самые сильные воздействия на типичный мотонейрон чаще всего исходят от ансамблей клеток, которые обычно меньше по размеру и располагаются поблизости. Давайте назовем совокупность всех мотонейронов и управляющих ими нейронных ансамблей «низшей моторной системой» и расчленим эту систему на функциональные субъединицы, назвав каждую «локальным моторным аппаратом», соответствующим определенной части тела: руке, ноге, глазу и т.д. Каждый локальный моторный аппарат есть, по-видимому, своего рода картотека, в которой хранятся программы, представляющие возможные движения определенной части тела. Мозг посылает сигнал в эту картотеку, используя свою систему нисходящих волокон, и выбирает подходящую программу. Но что в таком случае служит источником сигналов, посылаемых по нисходящим волокнам? Что именно оказывает влияние на локальный моторный аппарат? Мотонейроны располагаются внутри спинного, заднего и среднего мозга; в переднем мозгу нет ни одного. Здесь мы можем рассмотреть только проекции, которые сходятся на спинном мозгу. Они начинаются на всех уровнях центральной нервной системы. Внутри самого спинного мозга многие из них берут свое начало в группах вторичных сенсорных клеток, или даже, как в случае моносинаптических рефлекторных дуг, образуются коллатералями определенных первичных сенсорных волокон. В пределах заднего мозга пути исходят главным образом от внутренних двух третей заднемоэговой ретикулярной формации — области, которую называют крупноклеточной ретикулярной формацией, так как ее содержимое составляют крупные и очень крупные тела нейронов. В среднем мозгу пути исходят из верхнего двухолмия, а также из большой клеточной массы, называемой красным ядром. Вообще говоря, все эти три системы нисходящих волокон, идущих в спинной мозг (а именно: ретикуло-спинальная, текто-спинальная и рубро-спинальная), должны рассматриваться как несущие сообщения, или, если хотите, команды, которые могли зарождаться в обширных областях мозга. Верхнее двухолмие получает входные сигналы не только но зрительному нерву, но также из обширных областей коры, включая зрительную и многие другие. В красное ядро пути идут главным образом из мозжечка и моторной коры. Что же касается ретикулярной формации, то она особенно замечательна тем, что является местом, куда сходится информация самого различного происхождения. Мы уже рассуждали об этом, когда говорили о восходящих системах; при рассмотрении нисходящих систем это тоже существенно. Нейрон, представляющий данное свойство, показан на стр. 102; за основу этой модели взяты нейроны, электрическую активность которых регистрировал Дж. Моруцци (G. Moruzzi) из Пизанского университета, а также некоторые другие исследователи. Данный нейрон лежит в ретикулярной формации заднего мозга, и, по-видимому, он будет реагировать на поступление сигналов от группы вторичных сенсорных клеток спинного мозга. Однако вспышка света тоже способна активировать этот нейрон, поскольку сообщение об этом событии может успешно достичь ретикулярной формации по нисходящему пути из верхнего двухолмия. Кроме того, эта клетка реагирует и на сигналы из мозжечка, новой коры или ретикулярной формации среднего мозга. Короче, на этой клетке конвергирует большое число гетерогенных сигналов. Очевидно, ретикулярная формация должна интегрировать эти весьма разнообразные нервные сигналы, поступающие к ней по восходящим и нисходящим путям, а затем она может отправлять импульсы по ретикуло-спинальным волокнам, которые оканчиваются на промежуточных нейронах спинного мозга или даже, хотя и не часто, прямо на мотонейронах. Возможно, тут читатель опять услышит вопль электронщика: с инженерной точки зрения ретикулярная формация — бессмысленное образование. Теперь нам остается наложить на энцефало-спинальные системы заднего и среднего мозга те нисходящие системы, которые берут свое начало в переднем мозгу. Прежде всего практически все области новой коры проецируются в стриатум — наружную часть полосатого тела. В общей схеме связей сохраняется топографическое соответствие: соматосенсорная кора проецируется в область стриатума, отличную от той, куда приходятся зрительная проекция, слуховая проекция или проекции ассоциативных областей и моторной коры. Из стриатума масса волокон идет в бледный шар — внутренний отдел полосатого тела. В бледном шаре намного меньше нейронов, чем в стриатуме, так что эту систему можно рассматривать как своего рода воронку. Из бледного шара путь продолжается вниз, образуя пучок волокон, называемый чечевицеобразной петлей; здесь, однако, следует учесть любопытное исключение: большая часть волокон этой петли поворачивает назад в составе того же пучка и затем входит в верхнюю часть вентрального ядра. Мы отмечали, что в это клеточное ядро таламуса входят два больших соматосенсорных пути — медиальный лемниск и спино-таламический тракт и что оно проецируется на соматосенсорную кору. Однако только задняя часть вентрального ядра является промежуточной соматосенсорной станцией. В переднюю часть этого же клеточного скопления входят две мощные системы волокон: чечевицеобразная петля и восходящая проекция мозжечка — соединительная ножка. Эта часть вентрального ядра тоже проецируется в новую кору, но не в сенсорную, а в моторную зону. Патологические процессы, сопровождающиеся разрушением этой любопытной цепи, которая характеризуется наличием петель, вызывают серьезные двигательные расстройства. Одна из таких патологий касается пути в стриатум, который идет не из коры больших полушарий. Он идет из клеточной массы среднего мозга, нейроны которой пигментированы; у человека они черные даже на неокрашенных препаратах. По этой причине данная клеточная группа стала известна с конце XVIII в. как substantia nigra, т.е. черное вещество. Существенная потеря этих пигментированныхнейронов вызывает двигательное расстройство, известное под названием паркинсонизма. Это расстройство характеризуется ригидностью мышц, которая сильно затрудняет движения, и диагностируется по ряду симптомов, в том числе по маскообразному лицу. Кроме того, наблюдается также специфический (низкочастотный и почти вращательный) тремор рук. Однако первая жалоба больного обычно состоит в том, что ему трудно начать движение, которое он хочет сделать. Скажем, он собирается поправить какую-то деталь своего костюма, но ему почему-то трудно начать. Таким образом, можно считать, что полосатое тело существенно влияет на двигательную сферу. В более широком плане его можно рассматривать как принадлежащее к большому числу тех мозговых структур, выходные пути которых, по всей видимости, направлены к мотонейронам. Любопытно, что при этом полосатое тело не может непосредственно воздействовать на такиенейроны или хотя бы прямым путем влиять на те нейронные ансамбли, которые играют при мотонейронах роль привратников. Мы только что видели, что некоторая часть его выходного тракта, чечевицеобразной петли, поворачивает вверх и входит в вентральное ядро таламуса. Оставшаяся часть этой петли продолжает после этого поворота идти вниз, но далее каудальной границы среднего мозга не идет; на приведенном рисунке (стр. 102) один нейрон символизирует группу из нескольких тысяч нейронов, составляющих ядро ножки моста. Это часть ретикулярной формации среднего мозга. Начиная отсюда, нисходящие пути становятся неопределенными. Ретикулярная формация - это область, представляющая почти непреодолимые трудности для анатомического и функционального анализа. Волокна, проецирующие новую кору на стриатум, ни в коем случае не являются единственными кортикофугальными волокнами. Как мы отмечали, некоторые из выходных путей новой коры заканчиваются и различных ядрах таламуса, которые в свою очередь проецируются обратно на новую кору. Некоторые кортикофугальные волокна проникают в средний мозг, чтобы закончиться там в верхнем двухолмии, красном ядре и ретикулярной формации среднего мозга. Еще один контингент составляют волокна, идущие от всех частей новой коры и образующие синапсы на клетках варолиева моста - отдела заднего мозга, который в свою очередь проецируется на мозжечок. Остальные кортикофугальные волокна - те, которые проходят за пределами моста,- берут свое начало главным образом в моторной коре. Некоторые из них идут не далее чем до ретикулярной формации заднего мозга; другие достигают всех уровней спинного мозга. Эти последние волокна, которые составляют кортико-спинальный тракт, заслуживают особого упоминания. Само по себе замечательно уже то, что они проделывают путь от коры до спинного мозга, поскольку волокна, нисходящие из полосатого тела, доходят лишь до среднего мозга. Замечательно также то, что, согласно оценкам, 5 процентов кортико-спинальных волокон синаптически переключается прямо на мотонейроны. Это почти невероятный обходной путь: данные волокна не только входят в спинной мозг, но избегают нейронных ансамблей локальных моторных механизмов. Оказалось, что они «предпочитают» устанавливать синаптические связи с мотонейронами, которые заведуют мышцами конечностей. Несомненное наличие кортико-спинального тракта позволяет объяснить тот факт, что из всех областей коры больших полушарий моторная кора требует самой слабой электрической стимуляции для возбуждения в эксперименте движений тела. Объяснение состоит в том, что из всех областей коры больших полушарий мозга моторная кора удалена от мотонейронов на наименьшее число синаптических переключений. Моторная кора почти не поддается исследованию с точки зрения противопоставления произвольного и непроизвольного поведения. Рассмотрим обидную ситуацию, знакомую теннисистам. Игрок блестяще отбивает мяч и чувствует себя окрыленным. Но потом он приходит к выводу, что это была просто счастливая случайность: в следующий раз, когда теннисный мяч будет лететь к нему по сходной траектории, он может отбить его плохо. Это правда, что трудное произвольное движение было выполнено успешно, но есть ли в этом заслуга лица, которое его выполнило? Несмотря на загадку произвольного контроля, субъективный опыт с волевыми усилиями дал название моторной системе, которая иннервирует скелетную мускулатуру: это произвольная, или соматическая, нервная система, в отличие от непроизвольной, или автономной, нервной системы, которая иннервирует железы и гладкие мышцы внутренних органов. Однако последняя терминология не лучше застрахована от неправильного толкования, чем первая. Она имеет дело с термином «автономный», что значит «самоуправляемый». Автономная нервная система вовсе не является самоуправляемой. Ее функции увязаны с произвольными движениями не менее, чем с побуждениями и эмоциями. Короче, ее корни находятся в мозгу: каждому удается время от времени управлять не только сокращениями скелетных мышц, но и отправлением функций некоторыми внутренними органами. Тем не менее термин «автономная» победил во всем мире, говорящем по-английски. В других языках используются другие термины. На немецком говорят о висцеральной нервной системе (das viszerale Nervensystem), на французском — о вегетативной нервной системе (le systeme nerveux vegetatif). Автономная периферия условно изображена на рисунке (стр. 107) в виде трубчатого полого органа, под которым можно подразумевать кишечный тракт, мочевой пузырь, бронх или артерию; все они, в сущности, трубчатые структуры, толщина которых определяется числом слоев гладких мышц. Моторная иннервация такой мышечной ткани (или железы) включает два нейрона. Первый расположен в центральной нервной системе. Из него выходит очень тонкий аксон, который на периферии синаптически переключается на второй нейрон, часто располагающийся в ганглии. Второй нейрон в свою очередь посылает аксон для образования конечного соединения в висцеральную эффекторную ткань. В мозгу нейроны, которые специфически воздействуют на активность автономной нервной системы, сконцентрированы в гипоталамусе.
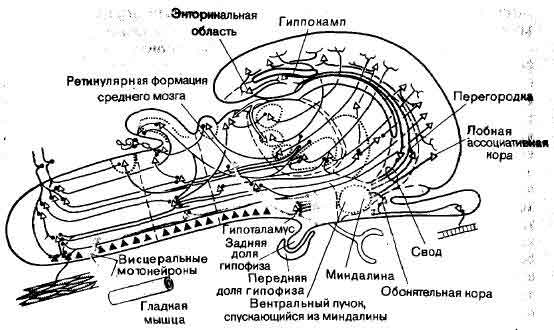
Цепи обратной связи замыкаются на гипоталамус, который регулирует активность желез и гладких мышц (таких, как не поддающиеся произвольному контролю мышцы внутренностей) через автономную нервную систему и гипофизарный комплекс. В гипоталамус приходят пути от гиппокампа и миндалины, которые являются главными компонентами так называемой лимбической системы. Кроме того, в гипоталамус поступают также волокна, исходящие из ретикупярной формации. Другим источником его входных сигналов является лобная кора, располагающаяся в самой передней части полушарий большого мозга. Лимбическая система-место назначения путей, исходящих из коры большого мозга, включая прямые проекционные волокна обонятельной коры. Доказательства этого очевидны; когда гипоталамус любого животного, а тем паче человека, внезапно разрушается, его обладатель умирает из-за сдвигов в том, что Клод Бернар назвал внутренней средой - этот термин охватывает те тканевые жидкости и функции органов, которые определяют кровяное давление, частоту пульса, частоту дыхания и т.д. В связи с этим нейрохирурги, которые вынуждены оперировать в области таламуса, всегда озабочены тем, чтобы не затронуть эту структуру. И все-таки больные умирают иногда от гипертермии (резкого повышения температуры тела) даже после успешной во всех других отношениях операции на мозге, в которой, казалось бы, были приняты все меры предосторожности, дабы не повредить гипоталамус. С другой стороны, когда массивное поражение гипоталамуса развивается постепенно, например в результате медленно растущей опухоли, острых тяжелых симптомов может вообще не быть. Создается впечатление, что в автономной системе существует командная цепь, или, как Клод Бернар определил это, автоматизм разных уровней: когда гипоталамус медленно теряет свои способности, области мозга ниже гипоталамуса сами могут обеспечить поддержание стабильности внутренней среды, хотя и в узких пределах. Все это хорошо согласуется с тем, что известно о путях автономной системы. Недавно были обнаружены волокна, идущие без переключений от гипоталамуса до мотонейронов автономной системы, расположенных в сером веществе спинного мозга, но они, по-видимому, составляют ничтожную часть выходящих из гипоталамуса волокон; для проведения нисходящих сигналов гипоталамус не имеет ничего похожего на кортико-спинальный тракт. Напротив, оказывается, что значительная часть волокон гипоталамуса идет не далее среднего мозга, где эстафету принимает ретикулярная формация. В действительности, для путей, нисходящих к автономным мотонейронам, типично прерывание на многих уровнях. При каждом таком прерывании в нисходящую линию проведения могут входить разнообразные дополнительные инструкции. Было бы разумно, если бы так было на самом деле. Жизнь зависит от иннервации внутренних органов; в каком-то смысле все остальное - биологическая роскошь. И жизненно важные системы должны быть построены по такому принципу, что никакое одиночное влияние не должно сильно сказываться на их работе. В действительности, конвергенция информации на мотонейронах для автономной нервной системы может быть столь же характерной, сколь и для соматической. Пока мы говорили о нисходящих нервных влияниях, которые в конечном счете достигают эффекторных тканей внутренних органов. А чем вызываются эти влияния? Или конкретнее: откуда и какие пути идут в гипоталамус? На рисунке (стр. 107) показан входной канал, который начинается в клетке ретикулярной формации среднего мозга — клетке, получающей свои входные сигналы от волокон спино-таламического тракта. Можно предположить, что при помощи этого пути таламус способен контролировать состояние внутренней среды. За пределами этого поиски входов в гипоталамус заводят далеко в область тканей мозга, причастных к эмоциям и побуждениям, область, в которой, скажем, эпилептические разряды могут, среди прочих симптомов, вызывать изменения в настроении, иногда до душевной боли или беспричинного страха. Это не должно нас удивлять. В конце концов эмоции и побуждения находят явное выражение в висцеральных и эндокринных изменениях. Значит, можно почти не сомневаться в том, что основные влияния на гипоталамус со стороны полушарий большого мозга исходят из гиппокампа и миндалины. Они делят свою «власть» с малым числом других частей полушарий. По этой причине общая ссылка на гиппокамп и миндалину оправдана: они являются главными компонентами того, что называют лимбической системой. Обратите внимание на показанную на рисунке двухканальную систему волокон, которая закругляется по краю новой коры и идет из гиппокампа в гипоталамус. Этот пучок образует свод. Он проходит вдоль свободного края мозгового плаща. И у кошки, и у обезьяны примерно две трети волокон этого пучка при выходе из гиппокампа направляются прямым путем в гипоталамус. Оставшаяся треть устанавливает синаптические связи в перегородке, из которой, как показано на рисунке, пути направляются также в гипоталамус. Мы уже указывали, что гиппокамп является станцией назначения для многоступенчатых проекций, которые размещены на обширной поверхности новой коры. Таким образом, при прослеживании путей управления висцеральной моторикой, как и в случае соматической моторики, оказывается, что по ходу прослеживания, если оно производится против течения, т. е. против направления передачи импульсов, приходится подключать к делу все большую часть великой промежуточной сети. Конечно, тут есть и отличие. Из данной области новой коры, скажем из первичной зрительной зоны, путь в гиппокамп может проходить с переключениями через ряд промежуточных неокортикальных полей. Концом неокортикального пути является энторинальная область коры большого мозга, смежная с гиппокампом и промежуточная по структуре между ним и новой корой. Выходящий отсюда последний отрезок завершает путь волокон в гиппокамп. В противоположность этому, путь в стриатум из любого поля новой коры является непрерывающимся. Рассмотрим далее миндалину. Хотя по цитоархитектонике она сильно отличается от гиппокампа, значительная часть ее выходных волокон тоже направляется в гипоталамус. Для гипоталамуса миндалина - это место, куда сходятся волокна из областей новой коры, синаптически удаленных от каких-либо первичных сенсорных полей. Однако она также является и адресатом волокон, которые начинаются в обонятельной коре; действительно, сюда проецируется энторинальная область. Кроме того, часть миндалины получает волокна от обонятельной луковицы. Таким образом, в случае обоняния передача сенсорных сигналов в лимбическую систему осуществляется поразительно прямым путем. Почему бы это могло быть? Почему обонятельная система находится в привилегированном положении по отношению к другим сенсорным системам? Дата добавления: 2016-06-06 | Просмотры: 929 | Нарушение авторских прав |