|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
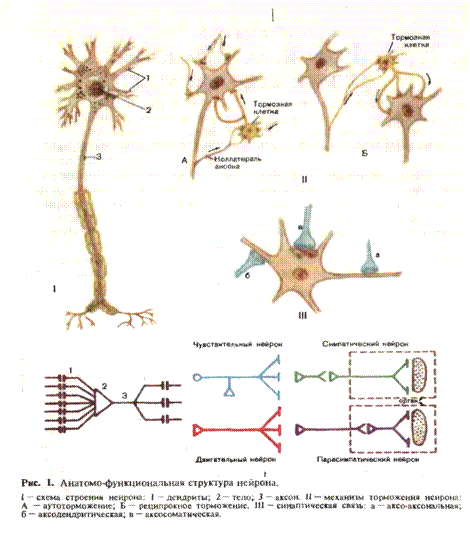
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ НЕРВНОЙ СИСТЕМЫОсновная функция нервной системы—объединение и регулирование различных физиологических процессов в соответствии с меняющимися условиями внешней и внутренней среды. Адаптация (приспособление) к окружающей среде не исключает определенной независимости организма. Чем выше уровень адаптации, тем менее однозначным становится реагирование организма на изменение условий жизни, тем большей свободой действий он располагает. Например, такой физиологический показатель, как постоянство температуры тела, независимо от температуры окружающей среды означает известную автономность по отношению к средовым влияниям. Чем выше уровень адаптации, тем сложнее и разнообразнее становятся реакции организма, дифференцированнее анализ средовых воздействий и тем большее значение в регуляции поведения приобреттет индивидуальный жизненный опыт. Нервная клетка. Основной структурно-функциональной единицей нервной системы является нервная клетка—нейрон (рис. 1), в котором различают тело клетки и отростки: дендриты и аксон. Нервный импульс распространяется всегда в одном направлении: по дендритам— ж телу клетки, по аксону—от тела клетки (закон динамической поляризации нервной клетки Рамон- и-Кахаля). Таким образом, нейрон — система, имеющая множество «входов» (дендриты) и лишь один «выход» (аксон). Такая закономерность свойственна нервной системе в целом. Количество волокон, несущих импульсы к центру, превосходит число волокон, несущих импульсы к периферии (принцип «воронки», или общего конечного пути). В функциональном отношении нейроны можно разграничить на афферентные, доставляющие импульсы к центру, эфферентные, несущие информацию от центра к периферии, и вставочные, в которых происходит предварительная промежуточная переработка импульсов и организуются коллатеральные связи. Особое место занимают так называемые рецепторные нейроны, представленные ложноуниполярными клетками, проводящими возбуждение от рецепторов в центральную нервную систему. Тела этих нейронов находятся вне центральной нервной системы, в так назы-ваемых нервных ганглиях, располагающихся по ходу корешков спи-нальньгх или черепных чувствительных нервов. В отличие от других нервных клеток рецепторный нейрон имеет два длинных отростка, один из которых является собственно аксоном, а другой—аксоно-подобно вытянутым дендритом. Рецепторные нейроны относятся к группе афферентных. Эфферентные нейроны, расположенные в центральной нервной системе, посылают импульсы к скелетным мышцам, обеспечивая регуляцию движения. Мотонейроны, непосредственно связанные с мышцами, расположены в передних рогах спинного мозга и двигательных ядрах черепных нервов. Наряду с ними в центральной нервной системе имеются эфферентные клетки, непосредственно не связанные с мышцами, а направляющие свои аксоны к мотонейронам передних рогов спинного мозга. Последние, таким образом, являются «приемщиками» многих нисходящих влияний, обработка совокупности которых обеспечивает дифференцированную регуляцию движений. Все разнообразие нисходящих эффрентных импульсаций в конечном итоге передается мышце аксонами этих мотонейронов.
Вставочные нейроны, или интернейроны, составляют самую многочисленную группу нервных клеток в центральной нервной системе и значительно различаются как по структуре, так и по выполняемой функции. Принципиально их разграничивают на возбуждающие и тормозящие. В центральной нервной системе тела нейронов сосредоточены в сером веществе — коре больших полушарий, подкорковых ядрах, ядрах ствола мозжечка, сером веществе спинного мозга Иннервацию внутренних органов обеспечивают афферентные и эфферентные нейроны, относящиеся к так называемой вегетативной, или автономной, нервной системе. Афферентная вегетативная клетка, несущая в спинной мозг информацию от интероцепторов внутренних органов, представляет собой, как и другие рецепторные нейроны, ложно униполярную клетку с аксоном и псевдоаксональным дендритом, тело которой расположено в спинальном узле. Эфферентная вегетативная клетка, находящаяся преимущественно в боковых столбах спинного мозга, не связана непосредственно с иннервируемой гладкой мышцей, чем отличается от переднерогового мотонейрона. Аксон эфферентного вегетативного нейрона заканчивается в вегетативном узле. Здесь расположен второй вегетативный эфферентный нейрон, аксон которого иннервирует соответствующий орган. Аксон первого нейрона называют преганглионарным, аксон второго — постганглионарным. Узлы симпатической нервной системы, как правило, расположены вне иннервируемого органа, поэтому постганглионарные волокна достаточно длинны. Узлы парасимпатической нервной системы находятся в самом органе, реже в непосредственной близости от него. Нервный импульс. Нервная клетка (тело и отростки) покрыта мембраной, избирательно проницаемой для ионов К+ в покое, а для ионов Na+ в состоянии возбуждения. Протоплазма нервных клеток по сравнению с внеклеточной жидкостью содержит в 30—50 раз больше ионов К+, в 8—10 раз меньше ионов Na+ и в 50 раз ионов С1-. За счет высокой концентрации ионов К+ в протоплазме нервной клетки по сравнению с внеклеточной жидкостью, избирательной проницаемости мембраны, возникает так называемый мембранный потенциал покоя, величина которого варьирует от 50 до 70 мВ. Величина потенциала покоя определяется соотношением числа положительно заряженных ионов К+, проникающих в единицу времени. из клетки во внеклеточную жидкость, к числу ионов Na+, диффундирующих через мембрану в клетку (что несколько снижает потенциал покоя, вызванный диффузией ионов К+ из клетки). Чем выше это соотношение, тем больше амплитуда мембранного потенциала покоя. Относительная стабильность концентрации ионов внутри клетки, несмотря на процесс диффузии, поддерживается благодаря существованию так называемого натри й-к алиевого насоса, обеспечивающего «удаление» из протоплазмы проникающих туда ионов Na+ и нагнетание в нее ионов К+, вследствие чего поддерживается ионная асимметрия. Энергетическое обеспечение работы насоса осуществляется при расщеплении макроэргических фосфорных соединений, в частности аденозинтрифосфата (АТФ), под воздействием фермента АТФазы. Ингибиция этого фермента приводит к «параличу» насоса. При воздействии электрического тока на нервное волокно возникает быстрое колебание величины мембранного потенциала, соответствующее образованию потенциала действия. При этом проницаемость мембраны для ионов Na+ резко повышается и становится в 20 раз больше, чем проницаемость для ионов К+: поток ионов Na+ внутрь клетки значительно преобладает над обратным потоком ионов К+. В связи с этим возникает реверсия потенциала мембраны, соответствующая восходящей части кривой потенциала действия — фазе деполяризации. Затем проницаемость мембраны для ионов Na+ понижается, а для ионов К+ повышается В итоге происходит реполяризация мембраны с появлением положительного потенциала на наружной ее поверхности и отрицательного — на внутренней поверхности. При возникновении потенциала действия (фазы деполяризации и реполяризации) протоплазма нервной клетки обогащается ионами Na+ и теряет ионы К+. При повторных стимуляциях потеря ионов К+ могла бы быть значительной, но этого не происходит вследствие действия натрий-калиевого насоса, активность которого зависит от концентрации ионов Na+ в протоплазме. Благодаря натрий-калиевому насосу восстанавливается нормальная ионная асимметрия между вне-и внутриклеточной жидкостью. Амплитуда потенциала действия варьирует от 80 до 110 мВ. Потенциал действия нейронов возникает при деполяризации мембраны до критического уровня, который неодинаков в различных участках клетки. Чем меньше необходимая степень деполяризации, тем выше возбудимость мембраны. При раздражении аксона достаточна деполяризация на 5—10 мВ, при раздражении клетки необходима деполяризация на 20—34 мВ. При возникновении потенциала действия в определенном участке нервного волокна между этим и соседним участком появляются разность потенциалов и электрический ток. Под влиянием электрического тока мембрана соседнего (невозбужденного) участка деполяризуется, повышается натриевая проницаемость и за счет последующей ионной диффузии потенциал действия как бы смещается на новый участок мембраны; возникает распространение возбуждения. После прохождения возбуждения мембрана нервного волокна вновь восстанавливает нормальную ионную асимметрию, положительный заряд на наружной и отрицательный на внутренней поверхности. Скорость проведения возбуждения (или импульса) определяется величиной омического сопротивления окружающей волокно среды, а также сопротивлением аксоплазмы на единицу длины, которая тем меньше, чем больше диаметр волокна. Скорость выше в волокнах большего диаметра, но при одном и том же диаметре скорость зависит от величины так называемого фактора надежности— отношения амплитуды потенциала действия к порогу деиннервации. При повышении фактора надежности скорость проведения импульса увеличивается. Аксоны многих нервных клеток покрыты миелиновой оболочкой, которая прерывается через каждые 2—3 мм, оставляя узкий участок неприкрытой миелином мембраны аксона (перехват Ранвье). Скорость проведения импульса в миелинизированных волокнах значительно выше, чем в немиелинизированных, и составляет до 120 м/с. Такая высокая скорость обеспечивается за счет особого по сравнению с немиелинизированными волокнами способа распространения возбуждения: не последовательно по ходу мембраны аксона, а сальтаторно с одного перехвата Ранвье на другой. Нервные волокна в центральной и периферической нервной системе тесно прилегают друг к другу, выполняя вместе с тем различные функции и передавая импульсы в разном направлении к разным структурам нервной системы. В связи с этим они должны быть изолированы друг от друга. Эта изоляция обеспечивается как миели-новыми оболочками, так и нейроглией, которой принадлежит важная роль в обеспечении нормального функционирования нервных клеток. Клетки нейроглии не только осуществляют изоляцию нейронов друг от друга, но и выполняют механическую опорную функцию и оказывают регулирующее влияние на ионный состав и метаболизм нервных клеток. Межнейрональные связи и нервно-мышечная передача. Функциональная общность нервной системы обеспечивается благодаря наличию тесной связи между отдельными элементами. У низших беспозвоночных животных имеется протоплазматическая непрерывность между отростками нервных клеток. На более поздних стадиях филогенеза и прежде всего у человека связь между нервными клетками осуществляется специальными образованиями—синапсами. Синапс состоит из трех основных элементов: пресинаптической мембраны, синаптической щели и постсинаптической мембраны. Пресинаптическая мембрана представляет собой нейросекреторный аппарат, в котором синтезируется и выделяется медиатор, оказывающий тормозящее или возбуждающее действие на постсинаптическую мембрану иннервируемой клетки. Постсинаптическая мембрана обладает избирательной чувствительность к химическому агенту — медиатору и практически нечувствительна к раздражителю электрическим током. Наличие синапсов определяет одностороннее проведение нервного импульса (обратная передача возбуждения с постсинаптической на пресинаптическую мембрану невозможна), поскольку в нервных волокнах возбуждение может распростра- няться в обе стороны от стимула. Вместе с тем в синапсе замедляется скорость проведения. Длительность синаптической задержки варьирует в значительных пределах в зависимости от функционального назначения синапса н составляет 0,2—0,5 мс в межнейрональных и нервно-мышечных синапсах, тогда как в нервных окончаниях гладкой мускулатуры достигает 5—10 мс. Терминали аксона, разветвляясь, могут оканчиваться на аксоне, дендритах или на теле следующей нервной клетки. В зависимости от этого различают аксо-аксональные, аксодендритические и аксосома-тические межнейрональные синапсы. Кроме того, имеются нервно-мышечные синапсы, обеспечивающие связь аксона мотонейрона с волокном скелетной мышцы или аксона эфферентной вегетативной клетки с волокнами гладкой мышцы. Нервный центр. Функциональная система. Существование поли- синаптической нервной сети создает возможность формирования сложных структур, способных относительно автономно регулировать те или иные функции. Комплекс нейронов, участвующих в регуляции какой-либо функции, обозначается как нервный центр. Понятие «нервный центр» применимо больше в физиологическом смысле, поскольку объединение нейронов в единую функциональную группу нередко распространяется на нервные клетки, расположенные в различных и далеко отстоящих друг от друга отделах нервной системы. Хотя в неврологии и употребляются такие термины, как «корковый центр иннервации взора», «дыхательный центр продолговатого мозга», «спи-нальный центр мочеиспускания», следует иметь в виду, что регуляция названных функций осуществляется при одновременном участии многих отделов нервной системы. Собственно в этом и заключается смысл ее интегративной деятельности. На ранних этапах эмбриогенеза между различными отделами нервной системы формируются четко дифференцированные, «жесткие» связи, создающие основу для жизненно необходимых врожденных реакций. Набор этих реакций обеспечивает первичную адаптацию после рождения (например, пищевые, дыхательные, защитные реакции). Совокупность нейронных групп, обеспечивающих ту или иную реакцию либо комплекс реакций, носит название функциональной системы. Понятие «функциональная система», предложенное акад. П. К. Анохиным, позволяет объяснить некоторые закономерности становления нервно-психических функций в онтогенезе. Важное значение имеет тот факт, что отдельные компоненты функциональной системы формируются примерно в одно и то же время, хотя могут принадлежать к филогенетически разным уровням. Вследствие этого в процессе эмбрионального развития наряду с общей последовательностью образования различных отделов нервной системы (по принципу: сначала эволюционно более древние, а затем более молодые) наблюдаются и отклонения от этой последовательности. Функциональные системы, имеющие первостепенное жизненное значение, формируются в первую очередь. Поскольку в функциональную систему объединяются разные в эволюционном плане уровни, то в пределах одного и того же уровня можно наблюдать разные степени созревания отдельных элементов в зависимости от их вовлеченности в функциональную систему. Принцип системогенеза важен для понимания не только особенностей развития нервной системы, но и тех нарушений, которые могут возникнуть в эмбриональном периоде. Чем в более раннем периоде внутриутробного развития отмечалось действие вредностей, тем грубее и диффузнее нарушения, тем на большее число органов и систем они распространяются. Благодаря взаимодействию центральных и периферических отделов нервой системы осуществляются регуляция отдельных физиологических функций, поддержание параметров внутренней среды на определенном уровне, выработка «решений» для реализации определенных программ действия в соответствии с ситуацией и потребностями организма. Рефлекторный принцип функционирования нервной системы. Основу деятельности нервной системы составляет рефлекторный принцип. В рефлекторной дуге различают афферентную часть (прием информации), центральную (переработка информации) и эфферентную часть (организация ответа). Для приема информации, ее обработки и регламентации силы и качества ответа необходимо наличие рецепторов, систем, обеспечивающих анализ информации и выработку решения, а также исполнительных центров. Для контроля за деятельностью исполнительных центров нужна информация о том, как выполняются приказы, насколько соответствует полученный результат запланированному, запрограммированному. Этот процесс контроля осуществляется при помощи «обратной» связи исполнительного аппарата с программирующим центром. В итоге формируется кольцевая структура: датчик первичной информации (рецептор)—анализирующий и программирующий центры — исполнительный аппарат—датчик информации к центру. Принцип кольцевой регуляции в нейрофизиологии является одним из существеннейших дополнений рефлекторного принципа. Введение понятия обратной связи позволило установить, каким образом осуществляется саморегуляция в живых системах. Поддержание какого-либо физиологического параметра на определенном уровне — активный, динамичный процесс, который осуществляется при помощи системы положительной и отрицательной обратной связи. Положительная обратная связь способствует усилению эфферентных влияний, отрицательная обратная связь приводит к ослаблению этих влияний. Если значение параметра опускается ниже заданного уровня, то в действие вступает положительная обратная связь, усиливаются эфферентные влияния и повышается значение параметра. Если это значение превышает заданный уровень, то по системе отрицательной обратной связи поступают сигналы, приводящие к ослаблению эфферентных влияний. В итоге при автоматическом регулировании все время происходит колебание значений параметра около заданного уровня. Чем меньше размах этих колебаний, тем более совершенно регулирование. Нарушение регуляторного механизма приводит к увеличению амплитуды колебаний. Свидетельством подобных расстройств являются тремор, дрожание конечностей, неустойчивость артериального давления.
ФИЛОГЕНЕЗ НЕРВНОЙ СИСТЕМЫ Сложные морфологические и функциональные особенности нервной системы возникли в результате длительной эволюции. На начальных этапах филогенеза нервная система представлена в виде синцития. Раздражение свободно распространяется по нервным клеткам, без особой дифференциации. В последующем отмечается образование узловой системы, появляются отдельные относительно самостоятельные узлы, цепочка узлов с сегментарным распределением. При этом нервные аппараты отдельных сегментов могут функционировать самостоятельно. У позвоночных наряду с сегментарной организацией нервной системы в оральном отделе развиваются важные образования, подчиняющие себе деятельность нижележащих отделов. Эта закономерность особенно проявляется у млекопитающих. В процессе филогенеза идет постепенная надстройка высших этажей центральной нервной системы, их постепенное усложнение. У позвоночных появляются большие полушария, хорошо развитые у высших позвоночных и представляющие основную массу головного мозга у человека. И. П. Павлов указывал: «Чем совершеннее нервная система животного, тем она централизованнее, тем высший ее отдел является все в большей степени распорядителем и распределителем всей деятельности организма, несмотря на то, что это вовсе ярко и открыто не выступает. Может казаться, что многие функции у высших животных идут совершенно вне влияния больших полушарий, а на самом деле это не так. Этот высший отдел держит в своем ведении все явления, происходящие в теле». Филогенез нервной системы можно представить как путь от простейшей рефлекторной дуги к многосинаптическим рефлекторным системам, обеспечивающим наиболее дифференцированные формы реагирования. Эволюционный подход в нейрофизиологии позволил более ясно представить смысл развития нервной системы. Любая живая ткань обладает свойством раздражимости, способностью так или иначе реагировать на внешние воздействия. Возникновение нервных клеток означало появление специализированного аппарата приема, накопления и перераспределения раздражающих стимулов, сначала в масштабе отдельных зон, а затем всего организма. Образование связей между нервными клетками и формирование примитивной нервной системы привели к качественно новому уровню интеграции организма. По сравнению с гуморальной регуляцией, эволюционно более древней, нервная регуляция отличается рядом преимуществ: быстротой проведения раздражения и более «прицельным» характером поведения. Гуморальные факторы сравнивают с сигналом, который «адресован всем». Передача раздражения по нервным путям скорее напоминает информацию, направленную по определенному адресу. В примитивной нервной системе возбуждение может' распространяться в любом направлении. Хотя при такой структуре невозможна тонкая координация реакций, все же обеспечивается участие всего организма в той или иной реакции. Накопление возбуждения в такой нервной сети уже создает предпосылки «памяти», т. е. реагирование на данный раздражитель может зависеть от предшествующих раздражителей, от краткой «предыстории» организма. Дальнейшее усложнение нервной системы заключается во все большей специализации нервных клеток, в появлении афферентных и эфферентных систем. Формирование рецепторов означало дифференцированное восприятие сигналов, настройку на определенные раздражители. Специализация нервных клеток сопровождалась появлением синапсов, обеспечивающих одностороннее проведение нервных импульсов. Вероятно, на этом этапе возйикают примитивные кольцевые структуры регуляции отдельных функций. При формировании автономных нервных ганглиев становится отчетливым сегментарный принцип иннервации: каждый нервный узел соответствует определенному сегменту тела. На уровне отдельного сег- мента осуществляется весьма четкая регуляция. Благодаря ганглиозной нервной системе становятся возможными сложные формы реагирования: в ганглиях «заложены» разнообразные программы действия. Однако сегменты связаны между собой недостаточно и еще не выражено координирующее влияние какого-либо одного центра. Подобные сложные автоматизмы широко представлены в мире насекомых. В ходе дальнейшей эволюции развитие нервной системы шло по пути постепенной цефализации, т. е. преобладания головных отделов, что привело К формированию головного мозга, коры больших полушарий как наивысшего отдела центральной нервной системы. Наибольшей сложности нервная система достигает у млекопитающих, у которых значительно развиты кора больших полушарий, мозолистое тело, соединяющее оба полушария, формируется пирамидная система, имеющая значение для иннервации тонких произвольных движений. Для нервной системы человека характерно дальнейшее развитие коры больших полушарий, особенно лобных долей. Поверхность коры у человека занимает 11/12 всей поверхности мозга, причем более 20% приходится на лобные доли. Пирамидная система у человека также достигает наивысшего развития.
КРАТКИЕ СВЕДЕНИЯ ПО АНАТОМИИ НЕРВНОЙ СИСТЕМЫ Нервная система человека условно разделяется на центральную и периферическую. К центральной нервной системе относятся головной мозг и спинной мозг. Черепные и спинномозговые нервы вместе с комплексом нервных узлов и нервных сплетений составляют периферическую нервную систему. На основании функционально-морфологических особенностей выде- В центральной нервной системе различают серое и белое вещество. Серое вещество — скопление тел нейронов, белое вещество — отростки нейронов, покрытых миелйновой оболочкой. В головном мозге серое вещество представлено в коре головного мозга, в коре мозжечка и в ядрах подкорковых узлов и ствола головного мозга. В спинном мозге серое вещество концентрируется в середине его, белое — на периферии. Усложнение форм поведения тесно связано с усложнением структуры и функций нервной системы. Это усложнение касается прежде всего головного мозга, который в процессе эволюции достиг высокого совершенства. В головном мозге человека различают высший отдел— новый мозг (neoencephalon), включающий основную массу коры больших полушарий; старый мозг (paleoencephalon), состоящий из различных по филогенетическому «возрасту» отделов обонятельного мозга; подкорковые ядра, входящие в состав стриопаллидарной системы; ствол мозга, в котором выделяют межуточный, средний мозг, мост мозга
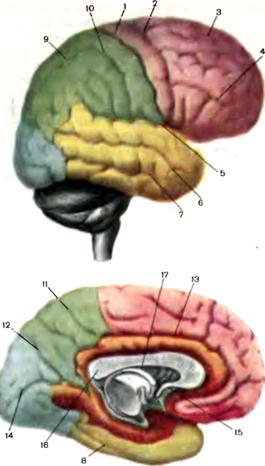
Рис. 2. Борозды и извилины головного мозга. Цветовые обозначения: розовый — лобная доля, зеленый — теменная доля, желтый — височная доля, синий — затылочная доля, оранжевый — обонятельный мозг.. Борозды: 1 — центральная; 2 —прецентральная; 3 — верхняя лобная; 4 — нижняя лобная; 5 - латеральная; 6 — верхняя височная; 7 — нижняя височная; 8 — обходная; 9 — вяутритеменная; 10 — постцентральная; 11 — краевая часть пояснон; 12 — теменно-затылочная; 13 — поясная; 14 — шпорная; 15 — передняя спайка; 16 — мозолистое тело; 17 — свод.
(варолиев мост) и продолговатый мозг. В неврологии к стволу мозга-относят также мозжечок. Большие полушария головного мозга. Большие полушария разделены по средней линии глубокой вертикальной щелью, в глубине которой правое и левое полушария соединены большой спайкой — мозолистым телом. В каждом полушарии различают лобную, теменную, височную, затылочную доли и островок (рис. 2). Доли мозговых полушарий отделяются друг от друга глубокими бороздами. Наиболее важны три глубокие борозды: центральная (ро- ландова), отделяющая лобную долю от теменной; латеральная (силь-виева), отделяющая височную долю от лобной и теменной, и теменно-за-тылочная, проходящая по внутренней поверхности полушария и отделяющая теменную долю от затылочной. Более мелкие борозды отделяют друг от друга мозговые извилины. Наличие борозд и мозговых извилин значительно увеличивает общую поверхность головного мозга. Общая поверхность площади коры полушарий составляет 2500 см2, причем 2/3 поверхности находится в глубине борозд, а 1/3 — на видимой поверхности полушарий. 1
5 Лобная доля отделяется от теменной центральной бороздой, от височной—латеральной (сильвиевой) бороздой На наружной поверхности лобной доли различают четыре извилины: вертикальную (пре-центральную) и три горизонтальные (верхнюю, среднюю и нижнюю). Вертикальная извилина заключена между центральной и прецент-ральной бороздами Верхняя лобная извилина расположена выше верхней лобной борозды, средняя лобная извилина—между верхней и нижней лобными бороздами, нижняя—между нижней лобной и сильвиевой бороздами. На нижней (базальной) поверхности лобных долей различают прямую и орбитальную извилину, которые образованы обонятельной и орбитальными бороздами. Прямая извилина залегает между внутренним краем полушария и обонятельной бороздой. В глубине обонятельной борозды лежат обонятельная луковица и обонятельный тракт. Лобная доля человека составляет 25—28% площади коры. Средняя масса лобной доли 450 г. В извилинах лобной доли сконцентрировано несколько функционально важных центров. Функция лобных долей связана с организацией произвольных движений, двигательных механизмов речи, регуляцией сложных форм поведения, процессов мышления. Теменная доля отделена от лобной центральной бороздой, от височной—сильвиевой бороздой, от затылочной—воображаемой линией от верхнего края теменно-затылочной борозды до нижнего края полушария. В теменной доле на наружной поверхности различают вертикальную постцентральную извилину и две горизонтальные дольки — верхнетеменную и нижнетеменную. Постцентральная извилина ограничена центральной и постцентральной бороздами, верхняя теменная долька расположена кверху от горизонтальной внутритеменной борозды, а нижняя — книзу от внутри-теменной борозды. Часть нижней теменной дольки, расположенную над задним отделом сильвиевой борозды, называют надкраевой (суп-рамаргинальной) извилиной, а часть, окружающую восходящий отросток верхней височной борозды, — угловой (ангулярной) извилиной. Функция теменной доли в основном связана с восприятием и анализом чувствительных раздражений, пространственной ориентации, регуляцией целенаправленных движений. Височная доля отделена от лобной и теменной долей сильвиевой бороздой. На наружной поверхности височной доли различают верхнюю, среднюю и нижнюю височные извилины. Верхняя височная извилина располагается между сильвиевой и верхней височной бороздами, средняя между верхней и нижней височными бороздами, нижняя — книзу от нижней височной борозды. На нижней базальной поверхности височной доли находится латеральная затылочно-височная извилина, граничащая с нижней височной извилиной, а более медиально — извилина гиппокампа. Функция височной доли связана с восприятием слуховых, вкусовых, обонятельных ощущений, анализом и синтезом речевых звуков, механизмами памяти. В глубине сильвиевой борозды расположена так называемая закрытая долька, или островок. Островок прикрыт участками лобной, теменной и височной долей, которые составляют покрышку, или operculum. От примыкающих соседних отделов островок отделен круговой бороздой островка. Поверхность островка разделена на две части (переднюю и заднюю) продольной центральной бороздой островка. Функция островка связана с восприятием вкуса. Затылочная доля занимает задние отделы полушарий. На наружной поверхности затылочная доля не имеет четких границ, отделяющих ее от теменной и височной долей. На внутренней поверхности полушария теменную долю от затылочной отграничивает темен-но-затылочная борозда. Борозды и извилины наружной поверхности затылочной доли непостоянны и имеют вариабельную топографию. Внутреннюю поверхность затылочной доли разделяет шпорная борозда на клин (треугольной формы долька затылочной доли) и язычную извилину. Функция затылочной доли связана с восприятием и переработкой зрительной информации. На медиальной поверхности полушарий над мозолистым телом находится поясная извилина, которая перешейком позади мозолистого тела переходит в парагиппокампову извилину. Поясная извилина вместе с парагиппокамповой извилиной составляет сводчатую извилину. На медиальной поверхности полушарий расположены участки коры, входящие в комплекс образований, относящихся к двум тесно связанным между собой функциональным системам — обонятельному мозгу и лим-бической системе. Обонятельный мозг состоит из двух отделов — периферического и центрального. Периферический отдел представлен обонятельным нервом, обонятельными луковицами, первичными обонятельными центрами. Центральный отдел включает извилину гиппокампа, зубчатую и сводчатую извилины (рис. 3). Периферический отдел обонятельного анализатора связан с корковыми областями обоих полушарий. Обонятельный мозг является одной из важнейших составных частей лимбической системы, объединяющей, кроме того, подкорковые структуры—хвостатое ядро, скорлупу, миндалевидное тело, таламус, гипоталамус, а также многочисленные пути, связывающие эти образования между собой Лимбическая система находится в тесной функциональной связи с ретикулярной формацией ствола мозга, составляет так называемый лимбико-ретикулярный комплекс.
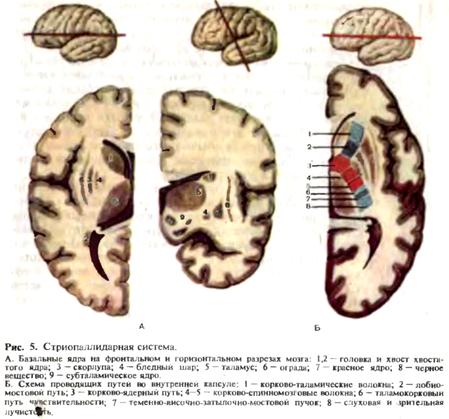
Архитектоника коры больших полушарий. Учение о структурных особенностях строения коры называется архитектоникой, об особенностях строения нервных волокон — миелоархитектоникой. Клетки коры больших полушарий менее специализированы, чем нейроны других отделов мозга, тем не менее определенные их группы анатомически и физиологически связаны с теми или иными специализированными отделами мозга. Микроскопическое строение коры головного мозга неодинаково в разных ее отделах. В коре больших полушарий предполагают наличие 10—13 млрд. нервных клеток и 100—130 млрд. клеток нейроглии. Для коры характерна шестислойность ее структуры: первый слой—зональный, второй —наружный зернистый, третий — пирамидный, четвертый — внутренний зернистый, пятый — слой больших пирамидных клеток, шестой — полиморфный. Наряду с этим в некоторых участках коры имеются отклонения от шестислойного типа строения. Эти отклонения группируют в два варианта: при первом отсутствуют наружный и внутренний зернистые слои за счет распространения в них пирамидных клеток, при втором, напротив, клетки-зерна вытесняют пирамидные клетки из третьего и пятого слоев. Крупные клетки III, V и VI слоев составляют большинство нервных клеток коры (до 8 млрд.) (рис. 4). Морфологические различия коры позволили выделить отдельные корковые цитоархитектонические поля. Имеется несколько вариантов классификаций корковых полей. Большинство исследователей в настоящее время выделяет около 50 цитоархитектонических полей. Микроскопическое строение коры довольно сложное. Установлено, что области коры, обеспечивающие регуляцию неодинаковых функций, как правило, имеют и различия в морфологии: морфологическая неоднородность и функциональная неоднородность коры больших полушарий -тесно связаны между собой. Бродман описал 52 цитоархитектонических поля в 11 областях коры больших полушарий: постцентральная область (поля 1, 2, 3, 43), прецентральная область (поля 4, 6), лобная область (поля 8—12, 44-47), островок (поля 13—16), теменная область (поля 5, 7, 39, 40), височная область (поля 20—22, 36—38, 41, 42, 52), затылочная область (поля 17—19), поясная область (поля 23—25, 31—33), ретроспленальная область (поля 26, 29, 30), область гиппокампа (поля 27, 28, 34, 35, 48) и обонятельная область (поле 51), обонятельный бугорок, периамигдалярная область. Подкорковые узлы. В толще белого вещества полушария мозга, вблизи от боковых желудочков, располагаются скопления серого вещества, называемые подкорковыми (или базальными) ядрами. К ним относятся хвостатое ядро, чечевицеобразное ядро, ограда и миндалевидное тело (рис. 5). Чечевицеобразное ядро, находящееся несколько кнаружи от хвостатого ядра, разделено на скорлупу и бледный шар. В функциональном отношении хвостатое ядро и скорлупа объединяются в стриатум, а бледный шар вместе с черным веществом и красными ядрами, расположенными в ножках мозга, а также субталамическим ядром —в паллидум. Вместе они представляют очень важное в функциональном отношении образование — стриопаллидарную систему. По морфологическим особенностям и филогенетическому происхождению паллидум является более древним, а стриатум — более молодым образованием. Паллидум содержит большое количество нервных волокон и некоторое количество крупных клеток. Хвостатое ядро и скорлупа включают в себя множество мелких и крупных клеток и небольшое количество нервных волокон. В стриарной системе имеется соматото-пическое распределение: в оральных отделах — голова, в средних — руки, в каудальных отделах—туловище и ноги. Между стриарной и паллидарной системами существует тесная связь. Стриопаллидарная система имеет многочисленные связи: пути, связывающие между собой образования стриопаллидарной системы; пути,
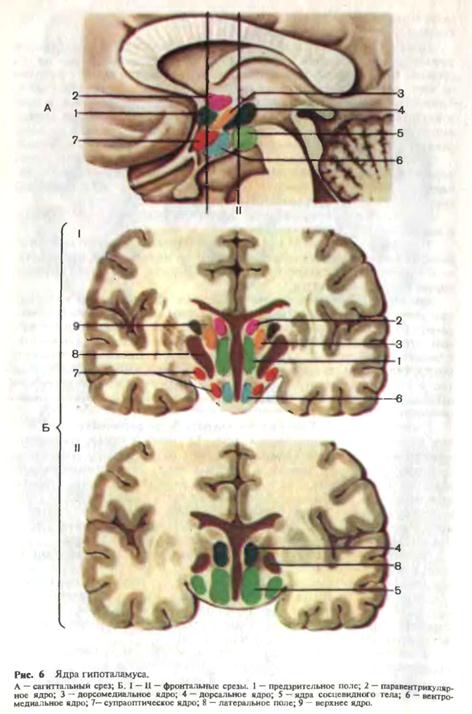
связывающие стриопаллидарную систему с конечным двигательным путем и мышцей; взаимные связи с различными отделами экстрапирамидной системы и коры больших полушарий, и, наконец, пути афферентации. Белое вещество больших полушарий состоит из нервных проводников, которые в соответствии с анатомическими и функциональными особенностями делят на ассоциативные, проекционные и комиссуральные. Ассоциативные волокна объединяют различные участки коры внутри одного полушария. Эти волокна бывают короткие и длинные. Короткие волокна обычно дугообразной формы. Они соединяют соседние извилины. Длинные волокна соединяют отдаленные участки коры. Проекционные волокна связывают полушария головного мозга с нижележащими отделами мозга—стволом и спинным мозгом. В составе проекционных волокон проходят проводящие пути, несущие афферентную (чувствительную) и эфферентную (двигательную) информацию. Комиссуральные волокна соединяют топографически идентичные участки правого и левого полушарий. Комиссуральные волокна образуют мозолистое тело, переднюю спайку, спайку свода и заднюю спайку. Основная масса комиссуральных волокон проходит через мозолистое тело, соединяя симметричные участки обоих полушарий головного мозга. Мозолистое тело—дугообразная тонкая пластинка. Удлиненная средняя часть мозолистого тела кзади переходит в утолщение, а спереди искривляется и дугообразно загибается вниз. Мозолистое тело соединяет филогенетически наиболее молодые участки полушарий и играет важную роль в обмене информацией между ними. От мозолистого тела в белое вещество полушарий мозга отходят нервные волокна. Эти волокна лучеобразно расходятся, направляясь во все доли мозга. Передняя спайка соединяет обонятельные области правого и левого полушарий. Спайка свода соединяет между собой гиппокамповы извилины правого и левого полушарий. В заднем отделе III желудочка располагаются задняя мозговая спайка и уздечковая спайка, содержащие волокна, соединяющие структуры промежуточного мозга. Внутренняя капсула. В глубине полушария, между хвостатым ядром и зрительным бугром с одной стороны и лентикулярным ядром — с другой располагается плотный слой проекционных волокон, имеющий на горизонтальном срезе вид тупого угла, открытого кнаружи. Во внутренней капсуле различаются переднюю ножку, заднюю ножку и соединяющее их колено. Все отделы капсулы представлены восходящими и нисходящими волокнами, соединяющими кору больших полушарий с нижележащими отделами головного и спинного мозга. Передняя ножка внутренней капсулы представлена эфферентными волокнами, направляющимися из коры лобной доли к зрительному бугру (корково-таламический путь) и к мозжечку через мост (корково-мостомозжеч-ковый путь). В колене и передних 2/3 задней ножки проходят нисходящие волокна от передней центральной извилины к передним рогам спинного мозга — корково-спинномозговой путь (в передних 2/3 задней ножки) и к двигательным ядрам черепных нервов — корково-ядерный путь (колено внутренней капсулы). В задней трети задней ножки проходят восходящие чувствительные волокна от зрительного бугра к задней центральной извилине (таламокорковый путь), восходящие пути зрительного и слухового анализаторов, направляющихся соответственно в затылочную и височную доли, а также нисходящие эфферентные волокна, идущие от нижних отделов затылочной и височной долей через мост к мозжечку (затылочно-височно-мостомозжечковый путь). Промежуточный мозг. К промежуточному мозгу относятся следующие образования, расположенные парамедианно вокруг III желудочка: зрительный бугор — таламус, надбугорье — эпиталамус, забугорье — ме-таталамус и подбугорье — гипоталамус. Зрительный бугор. Основная масса серого вещества промежуточного мозга относится к зрительному бугру, расположенному по обе стороны III желудочка. Прослойками белого вещества таламус разделен на ядра, которых к настоящему времени насчитывается до 150. Однако основными являются передние, вентролатеральные, медиальные, задние и внутрипластинчатые ядра. Таламус имеет яйцевидную форму, передний отдел его заострен (передний бугорок), а задний округлен и утолщен (подушка). Снаружи зрительный бугор ограничен внутренней капсулой. Кэпиталамусу относятся поводок и шишковидное тело. Шишковидное тело соединено с мозгом двумя пластинками белого вещества: верхняя переходит в поводки, соединенные между собой спайкой поводков, а нижняя направляется вниз к задней спайке мозга. Метаталам ус представлен двумя образованиями—медиальным и латеральным коленчатыми телами. Таламус и метаталамус является важнейшими афферентными центрами, коллектором восходящих афферентных импульсаций, относящихся к глубокой и поверхностной чувствительности, первичным анализатором зрения, слуха, вкуса. Зрительный бугор имеет многочисленные афферентные и эфферентные связи с корой, полосатым телом, красным ядром, верхними холмиками, гипоталамической областью, ретикулярной формацией и мозжечком, структурами лимбической системы. Шишковидное тело относится Гипоталамус. Книзу от зрительного бугра располагается скопление высокодифференцированных ядер (32 пары), которые составляют гипоталамус. Ядра гипоталамуса разделяют на три группы (переднюю, среднюю и заднюю), каждая из которых имеет свое функциональное значение. К переднему отделу относятся паравентрикулярные, супрахи-азмальные ядра, латеральные и медиальные части супраоптических ядер, к среднему отделу — задние отделы супраоптических ядер, ядер центрального серого вещества III желудочка, мамиллоинфундибулярные (передняя часть), паллидоинфундибулярные, интерфорникальное ядра. Задний отдел состоит из мамиллоинфундибулярных ядер (задняя часть), субталамического ядра (люисова тела), мамиллярного тела. Каждая группа ядер осуществляет регуляцию той или иной функции. Передние отделы гипоталамуса имеют отношение к интеграции преимущественно парасимпатической вегетативной нервной системы, задние — симпатической, средние обеспечивают регуляцию деятельности желез внутренней секреции, обмен веществ (рис. 6). В настоящее время в подбугорье различают так называемую суб-таламическую область, включающую субталамическое ядро, неопределенную зону, поля Форелля (Н1 и Н2) и некоторые другие образования. В функциональном отношении субталамическая область является частью
экстрапирамидной системы. В нижней части гипоталамуса располагаются серый бугор и воронка. Воронка заканчивается нижним придатком мозга—гипофизом. Гипофиз является одной из важнейших эндокринных желез; в филогенетическом и. функциональном отношении он тесно связан с гипоталамусом. В гипофизе различают переднюю долю (аденогипофиз), заднюю долю (нейрогипофиз), а также промежуточную часть, расположенную в виде каймы в задней части передней доли. Передняя доля развивается из эпителия глоточного кармана Ратке, а задняя—из инфун-дибудярного отростка гипоталамуса. Являясь важнейшим вегетативным центром, гипоталамус связан вегетативными волокнами с гипофизом, эпифизом и серым веществом в окружности Ш желудочка и водопровода мозга (сильвиева водопровода), вегетативными ядрами продолговатого мозга, ретикулярной формации ствола мозга, клетками боковых рогов спинного мозга. Кроме того, ядра гипоталамуса имеют многочисленные связи между собой, со зрительным бугром, стриопаллидарной системой, обонятельным мозгом, миндалевидным ядром и др. Ствол мозга. Включает средний мозг, мост мозга и продолговатый мозг (рис. 7). Сверху и кзади от ствола мозга находится мозжечок, связанный с каждым из отделов ствола мозга парой ножек: верхними — со средним мозгом, средними—с варолиевым мостом и нижними — с продолговатым мозгом. Ствол мозга во многом является аналогом спинного мозга. Двигательные ядра черепных нервов аналогичны переднему рогу спинного мозга, чувствительные — заднему рогу спинного мозга. В стволе мозга различают основание и покрышку. В основании проходят главным образом нисходящие пути, в покрышке — ядра черепных нервов и ретикулярная формация. Ретикулярная формация представляет собой комплекс нервных клеток и волокон и расположена в центральной части покрышки ствола мозга на всем его протяжении. Волокна ретикулярной формации достигают передних отделов зрительных бугров. К ретикулярной формации подходят многочисленные коллатерали от основных восходящих и нисходящих путей. Кроме того, ретикулярная формация имеет взаимные связи с гипоталамусом, зрительным бугром, образованиями стриопаллидарной системы, корой больших полушарий (особенно с корой лимбической системы), а также с мозжечком, ядрами ствола мозга и системой заднего продольного пучка. От ретикулярной формации отходят волокна к спинному мозгу (ретику-лоспинальные пути). Структуры ретикулярной формации участвуют в деятельности дыхательного и сосудодвигательного центров продолговатого мозга, центра взора моста. В стволе мозга располагаются также ядра и пути, относящиеся к стриопаллидарной системе (красное ядро, черное вещество), системе координации движений (нижняя олива), глубокой чувствительности (тонкое и клиновидное ядра), системе заднего продольного пучка (ядра Даршкевича) и др. Средний мозг. В центре среднего мозга проходит водопровод, соединяющий III и IV желудочки. Крышу среднего мозга составляет
четверохолмие, основание — ножки мозга; в средней части расположены ядра среднего мозга (рис. 7, Б). Ножки мозга острым углом сходятся к мосту мозга и, расходясь кпереди, образуют межножко-вое продырявленное пространство, сквозь которое проходят мозговые сосуды, васкуляризирующие глубинные структуры мозга. Ножки представляют собой плотные тяжи белого вещества, содержащие нисходящие пути от коры к переднему рогу спинного мозга, двигательным ядрам черепных нервов и мозжечку. Ножки мозга можно условно разделить на три части: наружную, среднюю и внутреннюю. Снаружи проходят волокна затылочно-височно-мостового пути, медиально — лобно-мос-товые пути, которые затем направляются к мозжечку. В средней части ножки мозга проходят волокна корково-спинномозгового и корково-ядерного путей таким образом, что волокна, иннервирующие мышцы лица, расположены медиально, мышцы нижних конечностей — латерально, а мышцы верхних конечностей — посредине. На границе ножек мозга с покрышкой располагается ядро черного вещества, лежащее в виде пластинки на проводящих путях. В среднем отделе между крышей (четверохолмием) и черным веществом находятся красное ядро, ядра III (глазодвигательного) и IV (блокового) нервов, задний продольный пучок, медиальная петля. Два ствола волокон задних продольных пучков расположены парамедианно у дна водопровода. Наружнее на этом же уровне лежат ядра III нерва (на уровне верхних холмиков) и IV нерва (на уровне нижних холмиков) так, что красное ядро оказывается помещенным между ними и задним продольным пучком с одной стороны и черным веществом—с другой. В латеральном отделе среднего мозга проходит мощный ствол афферентных волокон — медиальная петля, которую составляют два пути: бульботаламический, несущий импульсы глубокой чувствительности от тонкого и клиновидного ядер продолговатого мозга в зрительный бугор, и спино-таламический, являющийся проводником поверхностной чувствительности. В оральном отделе среднего мозга, кпереди от верхних холмиков, локализуются ядра заднего продольного пучка. В окружности водопровода среднего мозга находится мощный слой ретикулярной формации. Над водопроводом среднего мозга располагается пластинка крыши среднего мозга (четверохолмие), представленная волокнами и ядрами, относящимися к анализаторам зрения и слуха. Верхние холмики соединены тяжами белого вещества с наружными-коленчатыми телами, вместе с которыми они являются первичными зрительными центрами. Такая же связь имеется между нижними холмиками и внутренними коленчатыми телами (первичные слуховые центры). От ядер верхних и нижних холмиков начинаются волокна тегменто-спинального (покрышечно-спинномозгового) пути, участвующие в обеспечении старт-рефлексов. Мост головного мозга Лежит между средним мозгом и продолговатым мозгом. Вентральная часть моста образует толстый белый вал с резко выраженной поперечной волокнистостью. Посередине расположена основная борозда, в которой обычно лежит базилярная артерия. Оральная (передняя) часть моста содержит в основном продольные и поперечные волокна, в каудальной части, помимо проводящих волокон, находятся ядра черепных нервов (с V по VIII пару). Дорсальная поверхность моста представляет дно IV желудочка—ромбовидную ямку. На поперечном срезе моста границу между дорсальной и центральной его частью составляет трапециевидное тело, волокна которого относятся к системе слухового анализатора. В вентральной части располагаются продольные волокна пирамидного пути, рассеянного здесь на множество мелких пучков между собственными ядрами моста, с которыми он имеет коллатеральные связи. От собственных ядер моста берут начало поперечные волокна к мозжечку, которые составляют его средние ножки и относятся к корково-мостомозжечковому пути Поэтому существует прямая зависимость между развитием коры больших полушарий, вентральной части моста и мозжечка, и в связи с этим мост наиболее развит у человека. В дорсальной части моста мозга, находятся чувствительные пути: в латеральных отделах—спиноталамический путь, более медально—медиальная, петля, содержащая бульботаламический путь. В оральной части моста оба чувствительных пути сливаются в один плотный ствол (медиальная петля), идущий дорсолатерально в мосту и среднем мозге. Продолговатый мозг. Продолговатый мозг, являясь непосредственным продолжением спинного мозга, в своем строении во многом повторяет структуру спинного мозга. Как и другие отделы ствола мозга, продолговатый мозг представлен проводящими путями и ядрами. Оральный отдел продолговатого мозга граничит с мостом, а каудальный без четкой границы переходит в спинной мозг. Условно нижним краем продолговатого мозга считают перекрест пирамид или верхний край I шейного сегмента спинного мозга. На вентральной поверхности продолговатого мозга располагается передняя срединная щель, по обе стороны от которой лежат пирамиды. Снаружи от пирамид расположены нижние оливы, соответственно которым на поверхности продолговатого мозга определяется продольный валик. Нижние оливы отделены от пирамид боковой передней бороздой. В каудальном отделе продолговатого мозга ниже ромбовидной ямки на дорсальной поверхности различимы задние канатики, в которых проходят пучки Голля и Бурдаха (тонкие и клиновидные пучки), разде- ленные непарной задней медиальной бороздой и парными заднелате-ральными бороздами. В оральном отделе дорсальная поверхность продолговатого мозга открывается в полость IV желудочка, образуя задний угол ромбовидной ямки. Кнаружи отжраев ромбовидной ямки на боковой поверхности продолговатого мозга прослеживается продольный валик, соответствующий веревчатым телам—нижним ножкам мозжечка. На поперечном срезе продолговатого мозга в вентральном его отделе проходит корково-спинномозговой пуп, занимающий стволы пирамид. В центральной части располагаются волокна перекреста медиальной петли, несущие импульсы глубокой чувствительности от ядер Голля и Бурдаха к зрительному бугру. Вентролатеральные отделы продолговатого мозга занимают нижние оливы. Дорсальнее нижних олив проходят восходящие проводящие пути, составляющие нижние ножки мозжечка, а также чувствительный спиноталамический путь. В дорсальном отделе продолговатого мозга располагаются ядра каудальной группы черепных нервов (IX—XII пара), а также мощный слой ретикулярной формации. Ромбовидная ямка. Дно IV желудочка имеет форму ромба, как бы вдавленного в дорсальную поверхность моста и продолговатого мозга. Верхние стороны ромба ограничены верхними ножками мозжечка, нижние — нижними ножками. Верхние и нижние углы ромба соединяются продольной срединной бороздой. Кнаружи от этой борозды тянутся валики медиального возвышения, ограниченного снаружи бороздой (ограничивающая борозда). Из боковых углов ромбовидной ямки к середине тянутся парные мозговые полоски, разделяющие ромбовидную ямку на верхний и нижний треугольники (рис. 8).
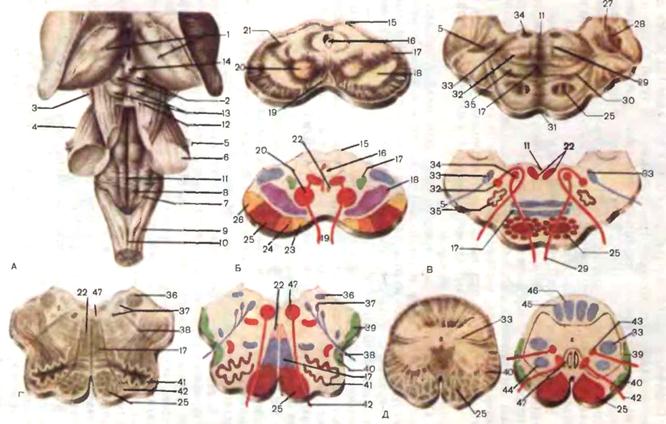
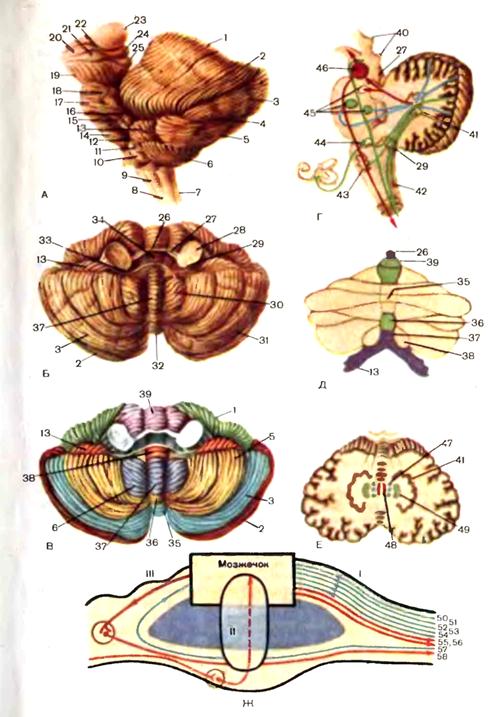
В области ромбовидной ямки лежат ядра V—XII нервов, проекция которых на дно IV желудочка имеет большое значение в топической диагностике поражения ствола мозга. Рассмотрим их расположение кнаружи от срединной борозды. Биссектриса нижнего угла определяет локализацию медиально расположенного ядра XII нерва и латерально расположенного дорсального ядра блуждающего нерва. В области верхнего угла располагаются ядра III и IV нервов. В наружном углу локализуются вестибулярные и слуховые ядра. Кнутри от них лежит параллельно срединной борозде чувствительное ядро тройничного нерва, еще более кнутри—ядро одиночного пути (вкусовое ядро), относящееся к системе IX и X нервов. Парамедианно кпереди от ядра XII нерва и дорсального ядра X нерва находятся двигательное ядро IX и X нервов и слюноотделительные ядра. В верхней части срединного возвышения располагается бугорок лицевого нерва, образованный волокнами внутреннего колена лицевого нерва, огибающими ядро отводящего нерва. Мозжечок. Расположен в задней черепной ямке под мостом мозга и продолговатым мозгом. Сверху он отделен от затылочных долей больших полушарий мозжечковым наметом. В мозжечке различают два полушария и червь, которые покрыты тонким слоем серого вещества—корой мозжечка. Поверхность коры мозжечка значительно увеличена за счет глубоких параллельных дугообразных борозд, разделяющих мозжечок на листки. В филогенетическом отношении в мозжечке различают его древнюю, старую и новую части. К древнему мозжечку относится маленькая долька — клочок, лежащая на основании полушария
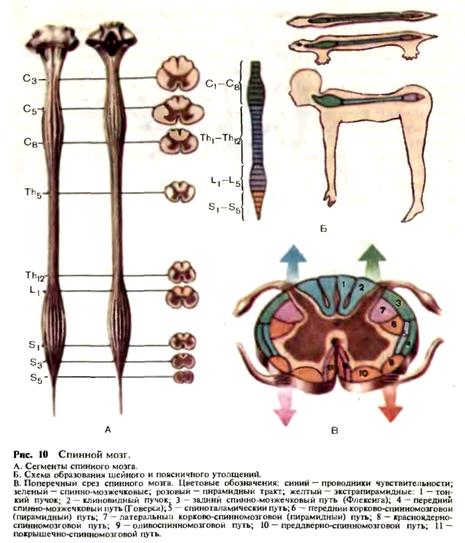
Рнс. 9. Мозжечок. А. Мозжечок и ствол мозга: 1 — четырехугольная долька; 2 — верхняя полулунная долька; 3 — нижняя полулунная долька; 4 — горизонтальная щель; 5 — двубрюшная долька; 6 — миндалина мозжечка; 7 — спинной мозг; 8 — шейные нервы; 9 — добавочный нерв; 10 — олива; 11 — блуждающий нерв; 12 — языкоглоточныи нерв; 13 — клочок; 14 — отводящий нерв; 15 — лицевой нерв; 16 — преддверно-улитковый нерв; 17 — мост; 18 — тройничный нерв; 19 — ножки мозга; 20 — зрительный тракт; 21 — медиальное коленчатое тело; 22 — латеральное коленчатое тело; 23 — подушка; 24 — верхний холмик; 25 — нижний холмик. Б, В. Общий вид мозжечка: 26 — язычок мозжечка; 27 — верхняя мозжечковая ножка; 28 — средняя мозжечковая ножка; 29 — нижняя мозжечковая ножка; 30 — миндалина; 31 — горизонтальная щель; 32 — червь; 33 — нижний мозговой парус; 34 — верхний мозговой парус; 35 — бугорок червя; 36 — пирамида червя; 37 — язычок червя; 38 — узелок; 39 — центральная долька. Г. Ножки мозжечка (сагиттальный разрез): 40 — крыша среднего мозга; 41 — зубчатое ядро; 42 — ядро задних канатиков; 43 — нижняя олива; 44 — вестибулярные ядра; 45 — ядра моста; 46 — красное ядро. Д- Филогенетическое распределение коры мозжечка. Цветовые обозначения: синий — археомозжечок, зеленый — палеомозжечок, желтый — неомозжечок. Е. Ядра мозжечка: 47 — шаровидное ядро; 48 — ядро шатра; 49 — пробковидное ядро. Ж. Ножки мозжечка: 1 — нижние ножки. Афферентные пути: 50 — задний спинно-мозжечковый (Флексига); 51 — от ядер тонкого и клиновидного; 52 — вестибуломозжечковый; 53 — ретикуломозжечковый; 54 — оливомозжечковый. Эфферентные пути; 55 — к латеральному вестибулярному ядру; 56 — к ретикулярной формации. II — средние ножки мозжечка: корково-мостомозжечковые волокна. 111 — верхние ножки мозжечка: 57 — передний спинно-мозжечковый путь (Говерса); 58 — эубчатокрасноядерно-спинно-мозговон путь. у средней ножки мозжечка, и связанная с клочком часть червя—узелок. К старому мозжечку относится червь, к новому—полушария (рис. 9). В белом веществе полушария и червя мозжечка имеется несколько ядер. Парамедиально расположено парное ядро шатра nucl. fastigii, латерально от него —мелкие островки серого вещества (шаровидное ядро — nucl. globosus), еще более латерально, вдаваясь в белое, вещество полушарий, — пробковидное ядро nucl. emboliformis. В белом веществе полушарий размещаются зубчатые ядра, nucl. dentatus. Указанные ядра, как и весь мозжечок, имеют разный филогенетический.возраст: ядро шара относится к древнему мозжечку, шаровидное и пробковидное — к старому, а зубчатое—к новому. Мозжечок имеет три пары ножек: верхние—направляются к пластинке крыши среднего мозга (четверохолмию), средние связывают мозжечок с мостом мозга, нижние — с продолговатым мозгом. Ножки мозжечка—это стволы проводящих путей, направляющихся к мозжечку или от мозжечка. В нижних ножках мозжечка проходят афферентные и эфферентные пути, в основном связанные со структурами червя мозжечка. Основные восходящие пути в нижних ножках мозжечка следующие: от задних рогов спинного мозга (задний спинно-мозжечковый путь), от вестибулярного ядра Бехтерева (вестибуломозжечковый путь), 'от ядер Голля и Бурдаха (бульбомозжечковый путь), от ретикулярной формации (ретикуломозжечковый путь), от нижней оливы (оливомозжечковый путь). Первые четыре пути заканчиваются в ядре шатра, оливомозжечковый путь —в коре мозжечка. В нижних ножках проходит несколько эфферентных путей, в конечном итоге направляющихся к переднему рогу спинного мозга: мозжечково-ретикулоспин-номозговой, мозжечково-вестибулоспинномозговой (через латеральное вестибулярное ядро Дейтерса), и мозжечково-оливоспинномозговой (см. рис. 9). В наиболее мощных средних ножках мозжечка проходят мосто-мозжечковые волокна, являющиеся частью корково-мостомозжечковых путей, идущих от верхней лобной извилины и нижних отделов затылочной и височной долей через мост к коре мозжечка. В верхних ножках мозжечка проходят афферентный путь от спинного мозга (передний спинно-мозжечковый путь) и нисходящий мозжечково-руброспинно-мозговой путь, идущий от зубчатого ядра полушария мозжечка через красное ядро к переднему рогу спинного мозга Спинной мозг. Представляет собой цилиндрический тяж, расположенный в позвоночном канале. Длина спинного мозга у взрослого человека составляет 42—46 см. По длиннику спинного мозга имеются два утолщения—шейное и поясничное. Образование этих утолщений тесно связано с сегментарным принципом строения спинного мозга (рис. 10). В спинном мозге насчитывается в общей сложности 31—32 сегмента: 8 шейных (С1—С8), 12 грудных (Th1—Th12), 5 поясничных (L1-L5), 5 крестцовых (S1-S5) и 1—2 копчиковых (Со1-Со2). Шейное утолщение располагается на уровне сегментов С5—Th1, и обеспечивает сегментарную иннервацию верхних конечностей. Поясничное утолщение соответствует сегментам L1-2-S1-2 осуществляет сегментарную
иннервацию нижних конечностей. Ниже поясничного утолщения спинной мозг заострен и образует мозговой конус, который заканчивается концевой нитью, достигающей копчиковых позвонков. На уровне каждого сегмента от спинного мозга отходят две пары передних и задних корешков. На небольшом расстоянии от спинного мозга задние корешки имеют выраженное утолщение — спинальный узел, содержащий чувствительные нервные клетки. На каждой стороне передний и задний корешки сливаются в единый ствол, образуя спинномозговой канатик. На поперечном срезе спинной мозг представлен серым и белым веществом. Серое вещество по форме напоминает крылья бабочки и имеет задние рога, содержащие чувствительные клетки; передние рога, содержащие двигательные клетки, и боковые рога, в которых преимущественно расположены вегетативные симпатические или парасимпатические нейроны, иннервирующие внутренние органы. Белое вещество состоит из покрытых миелиновыми оболочками нервных волокон, связывающих сегменты спинного мозга с центрами головного мозга. Используя контуры «бабочки» серого вещества, белое вещество спинного мозга условно разделяют на три канатика (столба): задний (кнутри от заднего рога), боковой (между задним и передним рогами) и передний (кнутри от переднего рога). Два задних канатика плотно прилегают друг к другу, а передние разделяются передней срединной щелью. В центре спинного мозга правая и левая половины серого вещества соединяются тонким перешейком (срединное промежуточное вещество), в центре которого располагается отверстие центрального канала. Задние канатики образованы восходящими проводниками глубокой чувствительности. Медиально располагаются проводники глубокой чувствительности от нижних конечностей (тонкий пучок Голля), латерально — от верхних конечностей (клиновидный пучок Бурдаха). Кроме того, в задних канатиках представлены проводники тактильной чувствительности. В боковых канатиках спинного мозга располагаются нисхр-дящие и восходящие проводники. К нисходящим прежде всего относятся волокна пирамидного (латерального корково-спинномозгового) пути, расположенного в задних отделах боковых канатиков у средней части заднего рога. Кпереди от пирамидного пути проходит красноядер-но-спинномозговой путь, а также ретикулоспинномозговой путь. Все нисходящие пути заканчиваются у клеток переднего рога спинного мозга. Вдоль всего латерального края бокового столба идут волокна восходящих спинно-мозжечковых путей: переднего спинно-мозжечково-го пути Говерса и заднего спинно-мозжечкового пути Флексига. Кнутри от пути Говерса проходят восходящие волокна поверхностной чувствительности (латеральный спиноталамический путь). Помимо этого, в боковых канатиках проходит восходящий спинопокрышечный путь, несущий проприоцептивную информацию к ядрам четверохолмия. Передние канатики спинного мозга составлены преимущественно нисходящими путями от передней центральной извилины, стволовых и подкорковых образований к переднему рогу спинного мозга (передний неперекрещенный пирамидный путь, вестибулоспинальныи путь, оливпспинномозговой путь и покрышечно-спинномозговой путь). Кроме этого, в передних канатиках проходит тонкий чувствительный пучок—передний спиноталамический путь. Состав клеток, находящихся в задних и передних рогах спинного мозга, неоднороден. В задних рогах располагаются чувствительные клетки, отростки которых переходят через среднюю линию спинного мозга в боковой столб противоположной стороны и составляют путь поверхностной чувствительности. В основании заднего рога выделяется обособленная группа клеток, относящихся к системе мозжечковой проприоцепции. Отростки этих клеток направляются в боковые столбы
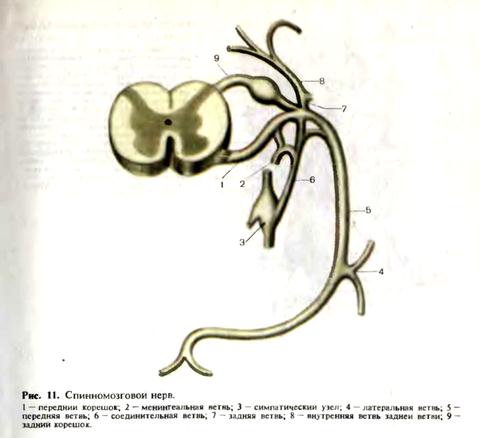
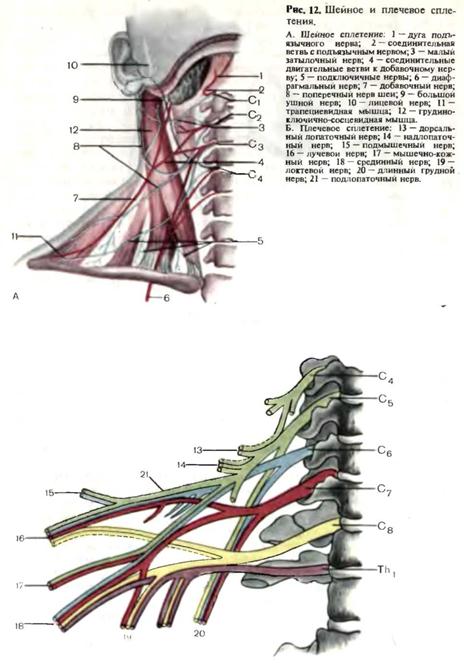
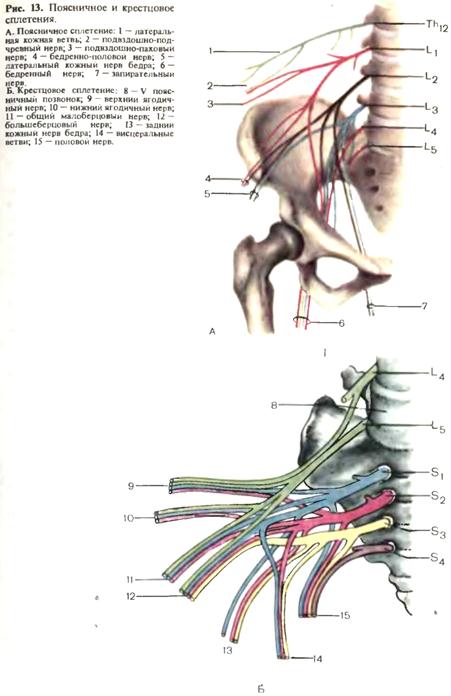
спинного мозга и в составе спиноцеребеллярных путей доходят до ядер червя мозжечка. В передних рогах спинного мозга различают три типа двигательных клеток: альфа-большие, альфа-малые и гамма-нейроны. В боковых рогах спинного мозга располагаются сегментарные вегетативные эфферентные клетки. На уровне сегментов C8-L3 располагаются эфферентные клетки симпатической нервной системы, на уровне C8-Th1 образован симпатический цилиоспинальный центр. На уровне крестовых сегментов S2-S4 находится спинальный парасимпатический центр регуляции функции тазовых органов. Периферическая нервная система. К периферической нервной системе относятся черепные и спинномозговые нервы, связанные со стволом головного мозга и спинным мозгом, их корешки и чувствительные узлы, а также нервные сплетения. Корешки спинномозговых нервов имеют строго сегментарное распределение и разделяются на задние, представленные чувствительными волокнами, и передние, представленные двигательными волокнами. Чувствительный межпозвоночный узел расположен по ходу заднего корешка вблизи от спинного мозга. В нем
Дата добавления: 2015-02-06 | Просмотры: 1729 | Нарушение авторских прав |