|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Неспецифические факторы защиты организмаМеханические факторы. Кожа и слизистые оболочки механически препятствуют проникновению микроорганизмов и других антигенов в организм. Последние все же могут попадать в организм при заболеваниях и повреждениях кожи (травмы, ожоги, воспалительные заболевания, укусы насекомых, животных и т. д.), а в некоторых случаях и через нормальную кожу и слизистую оболочку, проникая между клетками или через клетки эпителия (например, вирусы). Механическую защиту осуществляет также реснитчатый эпителий верхних дыхательных путей, так как движение ресничек постоянно удаляет слизь вместе с попавшими в дыхательные пути инородными частицами и микроорганизмами. Физико-химические факторы. Антимикробными свойствами обладают уксусная, молочная, муравьиная и другие кислоты, выделяемые потовыми и сальными железами кожи; соляная кислота желудочного сока, а также протеолитические и другие ферменты, имеющиеся в жидкостях и тканях организма. Особая роль в антимикробном действии принадлежит ферменту лизоциму. Этот протеолитический фермент получил название «мурамидаза», так как разрушает клеточную стенку бактерий и других клеток, вызывая их гибель и способствуя фагоцитозу. Лизоцим вырабатывают макрофаги и нейтрофилы. Содержится он в больших количествах во всех секретах, жидкостях и тканях организма (кровь, слюна, слезы, молоко, кишечная слизь, мозг и т. д.). Снижение уровня фермента приводит к возникновению инфекционных и других воспалительных заболеваний. В настоящее время осуществлен химический синтез лизоцима, и он используется как медицинский препарат для лечения воспалительных заболеваний. Иммунобиологические факторы. В процессе эволюции сформировался комплекс гуморальных и клеточных факторов неспецифической резистентности, направленных на устранение чужеродных веществ и частиц, попавших в организм. Гуморальные факторы неспецифической резистентности состоят из разнообразных белков, содержащихся в крови и жидкостях организма. К ним относятся белки системы комплемента, интерферон, трансферрин, β-лизины, белок пропердин, фибронектин и др. Белки системы комплемента обычно неактивны, но приобретают активность в результате последовательной активации и взаимодействия компонентов комплемента. Интерферон оказывает иммуномодулирующий, пролиферативный эффект и вызывает в клетке, инфицированной вирусом, состояние противовирусной резистентности. β -Лизины вырабатываются тромбоцитами и обладают бактерицидным действием. Трансферрин конкурирует с микроорганизмами за необходимые для них метаболиты, без которых возбудители не могут размножаться. Белок про-пердин участвует в активации комплемента и других реакциях. Сывороточные ингибиторы крови, например р-ингибиторы (р-липопротеины), инактивируют многие вирусы в результате неспецифической блокады их поверхности. Отдельные гуморальные факторы (некоторые компоненты комплемента, фибронектин и др.) вместе с антителами взаимодействуют с поверхностью микроорганизмов, способствуя их фагоцитозу, играя роль опсонинов. Большое значение в неспецифической резистентности имеют клетки, способные к фагоцитозу, а также клетки с цитотоксической активностью, называемые естественными киллерами, или NK-клетками. NK-клетки представляют собой особую популяцию лимфоцитоподобных клеток (большие гранулосодержащие лимфоциты), обладающих цитотоксическим действием против чужеродных клеток (раковых, клеток простейших и клеток, пораженных вирусом). Видимо, NK-клетки осуществляют в организме противоопухолевый надзор. В поддержании резистентности организма имеет большое значение и нормальная микрофлора организма.
№ 53 Комплемент, его структура, функции, пути активации, роль в иммунитете.
Природа и характеристика комплемента. Комплемент является одним из важных факторов гуморального иммунитета, играющим роль в защите организма от антигенов. Комплемент представляет собой сложный комплекс белков сыворотки крови, находящийся обычно в неактивном состоянии и активирующийся при соединении антигена с антителом или при агрегации антигена. В состав комплемента входят 20 взаимодействующих между собой белков, девять из которых являются основными компонентами комплемента; их обозначают цифрами: С1, С2, СЗ, С4... С9. Важную роль играют также факторы В, D и Р (пропердин). Белки комплемента относятся к глобулинам и отличаются между собой по ряду физико-химических свойств. В частности, они существенно различаются по молекулярной массе, а также имеют сложный субъединичный состав: Cl-Clq, Clr, Cls; СЗ-СЗа, СЗЬ; С5-С5а, С5b и т. д. Компоненты комплемента синтезируются в большом количестве (составляют 5—10% от всех белков крови), часть из них образуют фагоциты. Функции комплемента многообразны: а) участвует в лизисе микробных и других клеток (цитотоксическое действие); б) обладает хемотаксической активностью; в) принимает участие в анафилаксии; г) участвует в фагоцитозе. Следовательно, комплемент является компонентом многих иммунологических реакций, направленных на освобождение организма от микробов и других чужеродных клеток и антигенов (например, опухолевых клеток, трансплантата). Механизм активации комплемента очень сложен и представляет собой каскад ферментативных протеолитических реакций, в результате которого образуется активный цитолитический комплекс, разрушающий стенку бактерии и других клеток. Известны три пути активации комплемента: классический, альтернативный и лектиновый. По классическому пути комплемент активируется комплексом антиген-антитело. Для этого достаточно участия в связывании антигена одной молекулы IgM или двух молекул IgG. Процесс начинается с присоединения к комплексу АГ+АТ компонента С1, который распадается на субъединицы Clq, Clr и С Is. Далее в реакции участвуют последовательно активированные «ранние» компоненты комплемента в такой последовательности: С4, С2, СЗ. Эта реакция имеет характер усиливающегося каскада, т. е. когда одна молекула предыдущего компонента активирует несколько молекул последующего. «Ранний» компонент комплемента С3 активирует компонент С5, который обладает свойством прикрепляться к мембране клетки. На компоненте С5 путем последовательного присоединения «поздних» компонентов С6, С7, С8, С9 образуется литический или мембраноатакующий комплекс который нарушает целостность мембраны (образует в ней отверстие), и клетка погибает в результате осмотического лизиса. Альтернативный путь активации комплемента проходит без участия антител. Этот путь характерен для защиты от грамотрицательных микробов. Каскадная цепная реакция при альтернативном пути начинается с взаимодействия антигена (например, полисахарида) с протеинами В, D и пропердином (Р) с последующей активацией компонента СЗ. Далее реакция идет так же, как и при классическом пути — образуется мембраноатакующий комплекс. Лектиновыи путь активации комплемента также происходит без участия антител. Он инициируется особым маннозосвязывающим белком сыворотки крови, который после взаимодействия с остатками маннозы на поверхности микробных клеток катализирует С4. Дальнейший каскад реакций сходен с классическим путем. В процессе активации комплемента образуются продукты протеолиза его компонентов — субъединицы СЗа и СЗb, С5а и С5b и другие, которые обладают высокой биологической активностью. Например, СЗа и С5а принимают участие в анафилактических реакциях, являются хемоаттрактантами, СЗb — играет роль в опсонизации объектов фагоцитоза, и т. д. Сложная каскадная реакция комплемента происходит с участием ионов Са2+ и Mg2+.
№ 54 Интерфероны, природа. Способы получения и применения. Интерферон относится к важным защитным белкам иммунной системы. Открыт при изучении интерференции вирусов, т. е. явления, когда животные или культуры клеток, инфицированные одним вирусом, становились нечувствительными к заражению другим вирусом. Оказалось, что интерференция обусловлена образующимся при этом белком, обладающим защитным противовирусным свойством. Этот белок назвали интерфероном. Интерферон представляет собой семейство белков-гликопротеидов, которые синтезируются клетками иммунной системы и соединительной ткани. В зависимости от того, какими клетками синтезируется интерферон, выделяют три типа: α, β и γ-интерфероны. Альфа-интерферон вырабатывается лейкоцитами и он получил название лейкоцитарного; бета-интерферон называют фибробластным, поскольку он синтезируется фибробластами — клетками соединительной ткани, а гамма-интерферон — иммунным, так как он вырабатывается активированными Т-лимфоцитами, макрофагами, естественными киллерами, т. е. иммунными клетками. Интерферон синтезируется в организме постоянно, и его концентрация в крови держится на уровне примерно 2 МЕ/мл (1 международная единица — ME — это количество интерферона, защищающее культуру клеток от 1 ЦПД50 вируса). Выработка интерферона резко возрастает при инфицировании вирусами, а также при воздействии индукторов интерферона, например РНК, ДНК, сложных полимеров. Такие индукторы интерферона получили название интерфероногенов. Помимо противовирусного действия интерферон обладает противоопухолевой защитой, так как задерживает пролиферацию (размножение) опухолевых клеток, а также иммуномодулирующей активностью, стимулируя фагоцитоз, естественные киллеры, регулируя антителообразование В-клетками, активируя экспрессию главного комплекса гистосовместимости. Механизм действия интерферона сложен. Интерферон непосредственно на вирус вне клетки не действует, а связывается со специальными рецепторами клеток и оказывает влияние на процесс репродукции вируса внутри клетки на стадии синтеза белков. Применение интерферона. Действие интерферона тем эффективнее, чем раньше он начинает синтезироваться или поступать в организм извне. Поэтому его используют с профилактической целью при многих вирусных инфекциях, например гриппе, а также с лечебной целью при хронических вирусных инфекциях, таких как парентеральные гепатиты (В, С, D), герпес, рассеянный склероз и др. Интерферон дает положительные результаты при лечении злокачественных опухолей и заболеваний, связанных с иммунодефицитами. Интерфероны обладают видоспецифичностью, т. е. интерферон человека менее эффективен для животных и наоборот. Однако эта видоспецифичность относительна. Получение интерферона. Получают интерферон двумя способами: а) путем инфицирования лейкоцитов или лимфоцитов крови человека безопасным вирусом, в результате чего инфицированные клетки синтезируют интерферон, который затем выделяют и конструируют из него препараты интерферона; б) генно-инженерным способом — путем выращивания в производственных условиях рекомбинантных штаммов бактерий, способных продуцировать интерферон. Обычно используют рекомбинантные штаммы псевдомонад, кишечной палочки со встроенными в их ДНК генами интерферона. Интерферон, полученный генно-инженерным способом, носит название рекомбинантного. В нашей стране рекомбинантный интерферон получил официальное название «Реаферон». Производство этого препарата во многом эффективнее и дешевле, чем лейкоцитарного. Рекомбинантный интерферон нашел широкое применение в медицине как профилактическое и лечебное средство при вирусных инфекциях, новообразованиях и при иммунодефицитах.
№ 55 Видовой (наследственный) иммунитет. Врожденный, иди видовой, иммунитет, он же наследственный, генетический, конституциональный — это выработанная в процессе филогенеза генетически закрепленная, передающаяся по наследству невосприимчивость данного вида и его индивидов к какому-либо антигену (или микроорганизму), обусловленная биологическими особенностями самого организма, свойствами данного антигена, а также особенностями их взаимодействия. Примером может служить невосприимчивость человека к некоторым возбудителям, в том числе к особо опасным для сельскохозяйственных животных (чума крупного рогатого скота, болезнь Ньюкасла, поражающая птиц, оспа лошадей и др.), нечувствительность человека к бактериофагам, поражающим клетки бактерий. К генетическому иммунитету можно также отнести отсутствие взаимных иммунных реакций на тканевые антигены у однояйцовых близнецов; различают чувствительность к одним и тем же антигенам у различных линий животных, т. е. животных с различным генотипом. Объяснить видовой иммунитет можно с разных позиций, прежде всего отсутствием у того или иного вида рецепторного аппарата, обеспечивающего первый этап взаимодействия данного антигена с клетками или молекулами-мишенями, определяющими запуск патологического процесса или активацию иммунной системы. Не исключены также возможность быстрой деструкции антигена, например, ферментами организма или же отсутствие условий для приживления и размножения микроба (бактерий, вирусов) в организме. В конечном итоге это обусловлено генетическими особенностями вида, в частности отсутствием генов иммунного ответа к данному антигену. Видовой иммунитет может быть абсолютным и относительным. Например, нечувствительные к столбнячному токсину лягушки могут реагировать на его введение, если повысить температуру их тела. Белые мыши, не чувствительные к какому-либо антигену, приобретают способность реагировать на него, если воздействовать на них иммунодепрессантами или удалить у них центральный орган иммунитета — тимус.
№ 56 Понятие об иммунитете. Виды иммунитета. Иммунитет – это способ защиты организма от генетически чужеродных веществ – антигенов экзогенного и эндогенного происхождения, направленный на поддержание и сохранение гомеостаза, структурной и функциональной целостности организма, биологической (антигенной)индивидуальности каждого организма и вида в целом. Различают несколько основных видов иммунитета. Врожденный, иди видовой, иммунитет, он же наследственный, генетический, конституциональный — это выработанная в процессе филогенеза генетически закрепленная, передающаяся по наследству невосприимчивость данного вида и его индивидов к какому-либо антигену (или микроорганизму), обусловленная биологическими особенностями самого организма, свойствами данного антигена, а также особенностями их взаимодействия. Примером может служить невосприимчивость человека к некоторым возбудителям, в том числе к особо опасным для сельскохозяйственных животных (чума крупного рогатого скота, болезнь Ньюкасла, поражающая птиц, оспа лошадей и др.), нечувствительность человека к бактериофагам, поражающим клетки бактерий. К генетическому иммунитету можно также отнести отсутствие взаимных иммунных реакций на тканевые антигены у однояйцовых близнецов; различают чувствительность к одним и тем же антигенам у различных линий животных, т. е. животных с различным генотипом. Видовой иммунитет может быть абсолютным и относительным. Например, нечувствительные к столбнячному токсину лягушки могут реагировать на его введение, если повысить температуру их тела. Белые мыши, не чувствительные к какому-либо антигену, приобретают способность реагировать на него, если воздействовать на них иммунодепрессантами или удалить у них центральный орган иммунитета — тимус. Приобретенный иммунитет — это невосприимчивость к антигену чувствительного к нему организма человека, животных и пр., приобретаемая в процессе онтогенеза в результате естественной встречи с этим антигеном организма, например, при вакцинации. Примером естественного приобретенного иммунитета у человека может служить невосприимчивость к инфекции, возникающая после перенесенного заболевания, так называемый постинфекционный иммунитет (например, после брюшного тифа, дифтерии и других инфекций), а также «проиммуниция», т. е. приобретение невосприимчивости к ряду микроорганизмов, обитающих в окружающей среде и в организме человека и постепенно воздействующих на иммунную систему своими антигенами. В отличие от приобретенного иммунитета в результате перенесенного инфекционного заболевания или «скрытной» иммунизации, на практике широко используют преднамеренную иммунизацию антигенами для создания к ним невосприимчивости организма. С этой целью применяют вакцинацию, а также введение специфических иммуноглобулинов, сывороточных препаратов или иммунокомпетентных клеток. Приобретаемый при этом иммунитет называют поствакцинальным, и служит он для защиты от возбудителей инфекционных болезней, а также других чужеродных антигенов. Приобретенный иммунитет может быть активным и пассивным. Активный иммунитет обусловлен активной реакцией, активным вовлечением в процесс иммунной системы при встрече с данным антигеном (например, поствакцинальный, постинфекционный иммунитет), а пассивный иммунитет формируется за счет введения в организм уже готовых иммунореагентов, способных обеспечить защиту от антигена. К таким иммунореагентам относятся антитела, т. е. специфические иммуноглобулины и иммунные сыворотки, а также иммунные лимфоциты. Иммуноглобулины широко используют для пассивной иммунизации, а также для специфического лечения при многих инфекциях (дифтерия, ботулизм, бешенство, корь и др.). Пассивный иммунитет у новорожденных детей создается иммуноглобулинами при плацентарной внутриутробной передаче антител от матери ребенку ииграет существенную роль в защите от многих детских инфекций в первые месяцы жизни ребенка. Поскольку в формировании иммунитета принимают участие клетки иммунной системы и гуморальные факторы, принято активный иммунитет дифференцировать в зависимости от того, какой из компонентов иммунных реакций играет ведущую роль в формировании защиты от антигена. В связи с этим различают клеточный, гуморальный, клеточно-гуморальный и гуморально-клеточ-ный иммунитет. Примером клеточного иммунитета может служить противоопухолевый, а также трансплантационный иммунитет, когда ведущую роль в иммунитете играют цитотоксические Т-лимфоциты-киллеры; иммунитет при ток-синемических инфекциях (столбняк, ботулизм, дифтерия) обусловлен в основном антителами (антитоксинами); при туберкулезе ведущую роль играют иммунокомпетентные клетки (лимфоциты, фагоциты) с участием специфических антител; при некоторых вирусных инфекциях (натуральная оспа, корь и др.) роль в защите играют специфические антитела, а также клетки иммунной системы. В инфекционной и неинфекционной патологии и иммунологии для уточнения характера иммунитета в зависимости от природы и свойств антигена пользуются также такой терминологией: антитоксический, противовирусный, противогрибковый, противобактериальный, противопротозойный, трансплантационный, противоопухолевый и другие виды иммунитета. Наконец, иммунное состояние, т. е. активный иммунитет, может поддерживаться, сохраняться либо в отсутствие, либо только в присутствии антигена в организме. В первом случае антиген играет роль пускового фактора, а иммунитет называют стерильным. Во втором случае иммунитет трактуют как нестерильный. Примером стерильного иммунитета является поствакцинальный иммунитет при введении убитых вакцин, а нестерильного— иммунитет при туберкулезе, который сохраняется только в присутствии в организме микобактерий туберкулеза. Иммунитет (резистентность к антигену) может быть системным, т. е. генерализованным, и местным, при котором наблюдается более выраженная резистентность отдельных органов и тканей, например слизистых верхних дыхательных путей (поэтому иногда его называют мукозальным).
№ 57 Структура и функции иммунной системы. Кооперация иммунокомпетентных клеток. Структура иммунной системы. Иммунная система представлена лимфоидной тканью. Это специализированная, анатомически обособленная ткань, разбросанная по всему организму в виде различных лимфоидных образований. К лимфоидной ткани относятся вилочковая, или зобная, железа, костный мозг, селезенка, лимфатические узлы (групповые лимфатические фолликулы, или пейеровы бляшки, миндалины, подмышечные, паховые и другие лимфатические образования, разбросанные по всему организму), а также циркулирующие в крови лимфоциты. Лимфоидная ткань состоит из ретикулярных клеток, составляющих остов ткани, и лимфоцитов, находящихся между этими клетками. Основными функциональными клетками иммунной системы являются лимфоциты, подразделяющиеся на Т- и В-лимфоциты и их субпопуляции. Общее число лимфоцитов в человеческом организме достигает 1012, а общая масса лимфоидной ткани составляет примерно 1—2 % от массы тела. Лимфоидные органы делят на центральные (первичные) и периферические (вторичные). Функции иммунной системы. Иммунная система выполняет функцию специфической зашиты от антигенов, представляющую собой лимфоидную ткань, способную комплексом клеточных и гуморальных реакций, осуществляемых с помощью набора иммунореагентов, нейтрализовать, обезвредить, удалить, разрушить генетически чужеродный антиген, попавший в организм извне или образовавшийся в самом организме. Специфическая функция иммунной системы в обезвреживании антигенов дополняется комплексом механизмов и реакций неспецифического характера, направленных на обеспечение резистентности организма к воздействию любых чужеродных веществ, в том числе и антигенов. Кооперация иммунокомпетентных клеток. Иммунная реакция организма может иметь различный характер, но всегда начинается с захвата антигена макрофагами крови и тканей или же со связывания со стромой лимфоидных органов. Нередко антиген адсорбируется также на клетках паренхиматозных органов. В макрофагах он может полностью разрушаться, но чаше подвергается лишь частичной деградации. В частности, большинство антигенов в лизосомах фагоцитов в печение часа подвергается ограниченной денатурации и протеолизу. Оставшиеся от них пептиды (как правило, два-три остатка аминокислот) комплексируются с экспрессированными на внешней мембране макрофагов молекулами МНС. Макрофаги и все другие вспомогательные клетки, несущие на внешней мембране антигены, называются антигенпрезентирующими, именно благодаря им Т- и В-лимфоциты, выполняя функцию презентации, позволяют быстро распознавать антиген. Иммунный ответ в виде антителообразования происходит при распознавании В-клетками антигена, который индуцирует их пролиферацию и дифференциацию в плазмоцит. Прямое воздействие на В-клетку без участия Т-клеток могут оказать только тимуснезависимые антигены. В этом случае В-клетки кооперируются с Т-хелперами и макрофагами. Кооперация на тимусза-висимый антиген начинается с его презентации на макрофаге Т-хелперу. В механизме этого распознавания ключевую роль имеют молекулы МНС, так как рецепторы Т-хелперов распознают номинальный антиген как комплекс в целом или же как модифицированные номинальным антигеном молекулы МНС, приобретшие чужеродность. Распознав антиген, Т-хелперы секретируют γ-интерферон, который активирует макрофаги и способствует уничтожению захваченных ими микроорганизмов. Хелперный эффект на В-клетки проявляется пролиферацией и дифференциацией их в плазмоциты. В распознавании антигена при клеточном характере иммунного ответа, кроме Т-хелперов, участвуют также Т-киллеры, которые обнаруживают антиген на тех антигенпрезентирующих клетках, где он комплексируется с молекулами МНС. Более того, Т-киллеры, обусловливающие цитолиз, способны распознавать не только трансформированный, но и нативный антиген. Приобретая способность вызывать цитолиз, Т-киллеры связываются с комплексом антиген + молекулы МНС класса 1 на клетках-мишенях; привлекают к месту соприкосновения с ними цитоплазма-тические гранулы; повреждают мембраны мишеней после экзоцитоза их содержимого. В результате продуцируемые Т-киллерами лимфотоксины вызывают гибель всех трансформированных клеток организма, причем особенно чувствительны к нему клетки, зараженные вирусом. При этом наряду с лимфотоксином активированные Т-киллеры синтезируют интерферон, который препятствует проникновению вирусов в окружающие клетки и индуцирует в клетках образование рецепторов лимфотоксина, тем самым повышая их чувствительность к литическому действию Т-киллеров. Кооперируясь в распознавании и элиминации антигенов, Т-хелперы и Т-киллеры не только активируют друг друга и своих предшественников, но и макрофагов. Те же, в свою очередь, стимулируют активность различных субпопуляций лимфоцитов. Регуляция клеточного иммунного ответа, как и гуморального, осуществляется Т-супрессорами, которые воздействуют на пролиферацию цитотоксических и антигенпрезентирующих клеток. Цитокины. Все процессы кооперативных взаимодействий им-мунокомпетентных клеток, независимо от характера иммунного ответа, обусловливаются особыми веществами с медиаторными свойствами, которые секретируются Т-хелперами, Т-киллерами, мононуклеарными фагоцитами и некоторыми другими клетками, участвующими в реализации клеточного иммунитета. Все их многообразие принято называть цитокинами. По структуре цитокины являются протеинами, а по эффекту действия — медиаторами. Вырабатываются они при иммунных реакциях и обладают потенциирующим и аддитивным действием; быстро синтезируясь, цитокины расходуются в короткие сроки. При угасании иммунной реакции синтез цитокинов прекращается.
№ 58 Иммунокомпетентные клетки. Т- и В-лимфоциты, макрофаги, их кооперация. Иммунокомпетентные клетки - клетки, способные специфически распознавать антиген и отвечать на него иммунной реакцией. Такими клетками являются Т- и В-лимфоциты (тимусзависимые и костномозговые лимфоциты), которые под влиянием чужеродных агентов дифференцируются в сенсибилизированный лимфоцит и плазматическую клетку. Т-лимфоциты – это сложная по составу группа клеток, которая происходит от полипотентной стволовой клетки костного мозга, а созревает и дифференцируется в тимусе из предшественников. Т-лимфоциты разделяются на две субпопуляции: иммунорегуляторы и эффекторы. Задачу регуляции иммунного ответа выполняют Т-хелперы. Эффекторную функцияю осуществляют Т-киллеры и естественные киллеры. В орагнизме Т-лимфоциты обеспечивают клеточные формы иммунного ответа, определяют силу и продолжительность иммунной реакции. B-лимфоциты – преимущественно эффекторные иммунокомпетентные клетки. Зрелые В-лимфоциты и их потомки – плазматические клетки являются антителопродуцентами. Их основными продуктами являются иммуноглобулины. В-лимфоциты участвуют в формировании гуморального иммунитета, В-клеточной иммунологической памяти и гиперчувствительности немедленного типа. Макрофаги - клетки соединительной ткани, способные к активному захвату и перевариванию бактерий, остатков клеток и других чужеродных для организма частиц. Основная функция макрофагов сводится к борьбе с теми бактериями, вирусами и простейшими, которые могут существовать внутри клетки-хозяина, при помощи мощных бактерицидных механизмов. Роль макрофагов в иммунитете исключительно важна - они обеспечивают фагоцитоз, переработку и представление антигена T-клеткам. Кооперация иммунокомпетентных клеток. Иммунная реакция организма может иметь различный характер, но всегда начинается с захвата антигена макрофагами крови и тканей или же со связывания со стромой лимфоидных органов. Нередко антиген адсорбируется также на клетках паренхиматозных органов. В макрофагах он может полностью разрушаться, но чаше подвергается лишь частичной деградации. В частности, большинство антигенов в лизосомах фагоцитов в печение часа подвергается ограниченной денатурации и протеолизу. Оставшиеся от них пептиды (как правило, два-три остатка аминокислот) комплексируются с экспрессированными на внешней мембране макрофагов молекулами МНС. Макрофаги и все другие вспомогательные клетки, несущие на внешней мембране антигены, называются антигенпрезентирующими, именно благодаря им Т- и В-лимфоциты, выполняя функцию презентации, позволяют быстро распознавать антиген. Иммунный ответ в виде антителообразования происходит при распознавании В-клетками антигена, который индуцирует их пролиферацию и дифференциацию в плазмоцит. Прямое воздействие на В-клетку без участия Т-клеток могут оказать только тимуснезависимые антигены. В этом случае В-клетки кооперируются с Т-хелперами и макрофагами. Кооперация на тимусза-висимый антиген начинается с его презентации на макрофаге Т-хелперу. В механизме этого распознавания ключевую роль имеют молекулы МНС, так как рецепторы Т-хелперов распознают номинальный антиген как комплекс в целом или же как модифицированные номинальным антигеном молекулы МНС, приобретшие чужеродность. Распознав антиген, Т-хелперы секретируют γ-интерферон, который активирует макрофаги и способствует уничтожению захваченных ими микроорганизмов. Хелперный эффект на В-клетки проявляется пролиферацией и дифференциацией их в плазмоциты. В распознавании антигена при клеточном характере иммунного ответа, кроме Т-хелперов, участвуют также Т-киллеры, которые обнаруживают антиген на тех антигенпрезентирующих клетках, где он комплексируется с молекулами МНС. Более того, Т-киллеры, обусловливающие цитолиз, способны распознавать не только трансформированный, но и нативный антиген. Приобретая способность вызывать цитолиз, Т-киллеры связываются с комплексом антиген + молекулы МНС класса 1 на клетках-мишенях; привлекают к месту соприкосновения с ними цитоплазма-тические гранулы; повреждают мембраны мишеней после экзоцитоза их содержимого. В результате продуцируемые Т-киллерами лимфотоксины вызывают гибель всех трансформированных клеток организма, причем особенно чувствительны к нему клетки, зараженные вирусом. При этом наряду с лимфотоксином активированные Т-киллеры синтезируют интерферон, который препятствует проникновению вирусов в окружающие клетки и индуцирует в клетках образование рецепторов лимфотоксина, тем самым повышая их чувствительность к литическому действию Т-киллеров. Кооперируясь в распознавании и элиминации антигенов, Т-хелперы и Т-киллеры не только активируют друг друга и своих предшественников, но и макрофагов. Те же, в свою очередь, стимулируют активность различных субпопуляций лимфоцитов. Регуляция клеточного иммунного ответа, как и гуморального, осуществляется Т-супрессорами, которые воздействуют на пролиферацию цитотоксических и антигенпрезентирующих клеток. Цитокины. Все процессы кооперативных взаимодействий им-мунокомпетентных клеток, независимо от характера иммунного ответа, обусловливаются особыми веществами с медиаторными свойствами, которые секретируются Т-хелперами, Т-киллерами, мононуклеарными фагоцитами и некоторыми другими клетками, участвующими в реализации клеточного иммунитета. Все их многообразие принято называть цитокинами. По структуре цитокины являются протеинами, а по эффекту действия — медиаторами. Вырабатываются они при иммунных реакциях и обладают потенциирующим и аддитивным действием; быстро синтезируясь, цитокины расходуются в короткие сроки. При угасании иммунной реакции синтез цитокинов прекращается.
№ 59 Иммуноглобулины, структура и функции.
Природа иммуноглобулинов. В ответ на введение антигена иммунная система вырабатывает антитела — белки, способные специфически соединяться с антигеном, вызвавшим их образование, и таким образом участвовать в иммунологических реакциях. Относятся антитела к γ-глобулинам, т. е. наименее подвижной в электрическом поле фракции белков сыворотки крови. В организме γ-глобулины вырабатываются особыми клетками — плазмоцитами. γ-глобулины, несущие функции антител, получили название иммуноглобулинов и обозначаются символом Ig. Следовательно, антитела — это иммуноглобулины, вырабатываемые в ответ на введение антигена и способные специфически взаимодействовать с этим же антигеном. Функции. Первичная функция состоит во взаимодсйствии их активных центров с комплементарными им детерминантами антигенов. Вторичная функция состоит в их способности: • связывать антиген с целью его нейтрализации и элиминации из организма, т. е. принимать участие в формировании защиты от антигена; • участвовать в распознавании «чужого» антигена; • обеспечивать кооперацию иммунокомпетентных клеток (макрофагов, Т- и В-лимфоцитов); • участвовать в различных формах иммунного ответа (фагоцитоз, киллерная функция, ГНТ, ГЗТ, иммунологическая толерантность, иммунологическая память). Структура антител. Белки иммуноглобулинов по химическому составу относятся к гликопротеидам, так как состоят из протеина и Сахаров; построены из 18 аминокислот. Имеют видовые отличия, связанные главным образом с набором аминокислот. Их молекулы имеют цилиндрическую форму, они видны в электронном микроскопе. До 80 % иммуноглобулинов имеют константу седиментации 7S; устойчивы к слабым кислотам, щелочам, нагреванию до 60 °С. Выделить иммуноглобулины из сыворотки крови можно физическими и химическими методами (электрофорез, изоэлектрическое осаждение спиртом и кислотами, высаливание, аффинная хроматография и др.). Эти методы используют в производстве при приготовлении иммунобиологических препаратов. Иммуноглобулины по структуре, антигенным и иммунобиологическим свойствам разделяются на пять классов: IgM, IgG, IgA, IgE, IgD. Иммуноглобулины М, G, А имеют подклассы. Например, IgG имеет четыре подкласса (IgG,, IgG2, IgG3, IgG4). Все классы и подклассы различаются по аминокислотной последовательности. Молекулы иммуноглобулинов всех пяти классов состоят из полипептидных цепей: двух одинаковых тяжелых цепей Н и двух одинаковых легких цепей — L, соединенных между собой дисульфидными мостиками. Соответственно каждому классу иммуноглобулинов, т.е. М, G, A, E, D, различают пять типов тяжелых цепей: μ (мю), γ (гамма), α (альфа), ε (эпсилон) и Δ (дельта), различающихся по антигенности. Легкие цепи всех пяти классов являются общими и бывают двух типов: κ (каппа) и λ (ламбда); L-цепи иммуноглобулинов различных классов могут вступать в соединение (рекомбинироваться) как с гомологичными, так и с гетерологичными Н-цепями. Однако в одной и той же молекуле могут быть только идентичные L-цепи (κ или λ). Как в Н-, так и в L-цепях имеется вариабельная — V область, в которой последовательность аминокислот непостоянна, и константная — С область с постоянным набором аминокислот. В легких и тяжелых цепях различают NH2- и СООН-концевые группы. При обработке γ -глобулина меркаптоэтанолом разрушаются дисульфидные связи и молекула иммуноглобулина распадается на отдельные цепи полипептидов. При воздействии протеолитическим ферментом папаином иммуноглобулин расщепляется на три фрагмента: два не кристаллизующихся, содержащих детерминантные группы к антигену и названных Fab-фрагментами I и II и один кристаллизующий Fc-фрагмент. FabI- и FabII-фрагменты сходны по свойствам и аминокислотному составу и отличаются от Fc-фрагмента; Fab-и Fc-фрагменты являются компактными образованиями, соединенными между собой гибкими участками Н-цепи, благодаря чему молекулы иммуноглобулина имеют гибкую структуру. Как Н-цепи, так и L-цепи имеют отдельные, линейно связанные компактные участки, названные доменами; в Н-цепи их по 4, а в L-цепи — по 2. Активные центры, или детерминанты, которые формируются в V-областях, занимают примерно 2 % поверхности молекулы иммуноглобулина. В каждой молекуле имеются две детерминанты, относящиеся к гипервариабельным участкам Н-и L-цепей, т. е. каждая молекула иммуноглобулина может связать две молекулы антигена. Поэтому антитела являются двухвалентными. Типовой структурой молекулы иммуноглобулина является IgG. Остальные классы иммуноглобулинов отличаются от IgG дополнительными элементами организации их молекулы. В ответ на введение любого антигена могут вырабатываться антитела всех пяти классов. Обычно вначале вырабатывается IgM, затем IgG, остальные — несколько позже.
№ 60 Классы иммуноглобулинов, их характеристика.
Иммуноглобулины по структуре, антигенным и иммунобиологическим свойствам разделяются на пять классов: IgM, IgG, IgA, IgE, IgD. Иммуноглобулин класса G. Изотип G составляет основную массу Ig сыворотки крови. На его долю приходится 70—80 % всех сывороточных Ig, при этом 50 % содержится в тканевой жидкости. Среднее содержание IgG в сыворотке крови здорового взрослого человека 12 г/л. Период полураспада IgG — 21 день. IgG — мономер, имеет 2 антигенсвязывающих центра (может одновременно связать 2 молекулы антигена, следовательно, его валентность равна 2), молекулярную массу около 160 кДа и константу седиментации 7S. Различают подтипы Gl, G2, G3 и G4. Синтезируется зрелыми В-лимфоцитами и плазматическими клетками. Хорошо определяется в сыворотке крови на пике первичного и при вторичном иммунном ответе. Обладает высокой аффинностью. IgGl и IgG3 связывают комплемент, причем G3 активнее, чем Gl. IgG4, подобно IgE, обладает цитофильностью (тропностью, или сродством, к тучным клеткам и базофилам) и участвует в развитии аллергической реакции I типа. В иммунодиагностических реакциях IgG может проявлять себя как неполное антитело. Легко проходит через плацентарный барьер и обеспечивает гуморальный иммунитет новорожденного в первые 3—4 месяца жизни. Способен также выделяться в секрет слизистых, в том числе в молоко путем диффузии. IgG обеспечивает нейтрализацию, опсонизацию и маркирование антигена, осуществляет запуск комплемент-опосредованного цитолиза и антителозависимой клеточно-опосредованной цитотоксичности. Иммуноглобулин класса М. Наиболее крупная молекула из всех Ig. Это пентамер, который имеет 10 антигенсвязывающих центров, т. е. его валентность равна 10. Молекулярная масса его около 900 кДа, константа седиментации 19S. Различают подтипы Ml и М2. Тяжелые цепи молекулы IgM в отличие от других изотипов построены из 5 доменов. Период полураспада IgM — 5 дней. На его долю приходится около 5—10 % всех сывороточных Ig. Среднее содержание IgM в сыворотке крови здорового взрослого человека составляет около 1 г/л. Этот уровень у человека достигается уже к 2—4-летнему возрасту. IgM филогенетически — наиболее древний иммуноглобулин. Синтезируется предшественниками и зрелыми В-лимфоцитами. Образуется в начале первичного иммунного ответа, также первым начинает синтезироваться в организме новорожденного — определяется уже на 20-й неделе внутриутробного развития. Обладает высокой авидностью, наиболее эффективный активатор комплемента по классическому пути. Участвует в формировании сывороточного и секреторного гуморального иммунитета. Являясь полимерной молекулой, содержащей J-цепь, может образовывать секреторную форму и выделяться в секрет слизистых, в том числе в молоко. Большая часть нормальных антител и изоагглютининов относится к IgM. Не проходит через плаценту. Обнаружение специфических антител изотипа М в сыворотке крови новорожденного указывает на бывшую внутриутробную инфекцию или дефект плаценты. IgM обеспечивает нейтрализацию, опсонизацию и маркирование антигена, осуществляет запуск комплемент-опосредованного цитолиза и антителозависимой клеточно-опосредованной цитотоксичности. Иммуноглобулин класса А. Существует в сывороточной и секреторной формах. Около 60 % всех IgA содержится в секретах слизистых. Сывороточный IgA: На его долю приходится около 10—15% всех сывороточных Ig. В сыворотке крови здорового взрослого человека содержится около 2,5 г/л IgA, максимум достигается к 10-летнему возрасту. Период полураспада IgA — 6 дней. IgA — мономер, имеет 2 антигенсвязывающих центра (т. е. 2-валентный), молекулярную массу около 170 кДа и константу седиментации 7S. Различают подтипы А1 и А2. Синтезируется зрелыми В-лимфоцитами и плазматическими клетками. Хорошо определяется в сыворотке крови на пике первичного и при вторичном иммунном ответе. Обладает высокой аффинностью. Может быть неполным антителом. Не связывает комплемент. Не проходит через плацентарный барьер. IgA обеспечивает нейтрализацию, опсони-зацию и маркирование антигена, осуществляет запуск антителозависимой клеточно-опос-редованной цитотоксичности. Секреторный IgA: В отличие от сывороточного, секреторный sIgA существует в полимерной форме в виде ди- или тримера (4- или 6-валентный) и содержит J- и S-пeптиды. Молекулярная масса 350 кДа и выше, константа седиментации 13S и выше. Синтезируется зрелыми В-лимфоцитами и их потомками — плазматическими клетками соответствующей специализации только в пределах слизистых и выделяется в их секреты. Объем продукции может достигать 5 г в сутки. Пул slgA считается самым многочисленным в организме — его количество превышает суммарное содержание IgM и IgG. В сыворотке крови не обнаруживается. Секреторная форма IgA — основной фактор специфического гуморального местного иммунитета слизистых оболочек желудочно-кишечного тракта, мочеполовой системы и респираторного тракта. Благодаря S-цепи он устойчив к действию протеаз. slgA не активирует комплемент, но эффективно связывается с антигенами и нейтрализует их. Он препятствует адгезии микробов на эпителиальных клетках и генерализации инфекции в пределах слизистых. Иммуноглобулин класса Е. Называют также реагином. Содержание в сыворотке крови крайне невысоко — примерно 0,00025 г/л. Обнаружение требует применения специальных высокочувствительных методов диагностики. Молекулярная масса — около 190 кДа, константа седиментации — примерно 8S, мономер. На его долю приходится около 0,002 % всех циркулирующих Ig. Этот уровень достигается к 10—15 годам жизни. Синтезируется зрелыми В-лимфоцитами и плазматическими клетками преимущественно в лимфоидной ткани бронхолегочного дерева и ЖКТ. Не связывает комплемент. Не проходит через плацентарный барьер. Обладает выраженной цитофильностью — тропностью к тучным клеткам и базофилам. Участвует в развитии гиперчувствительности немедленного типа — реакция I типа. Иммуноглобулин класса D. Сведений об Ig данного изотипа не так много. Практически полностью содержится в сыворотке крови в концентрации около 0,03 г/л (около 0,2 % от общего числа циркулирующих Ig). IgD имеет молекулярную массу 160 кДа и константу седиментации 7S, мономер. Не связывает комплемент. Не проходит через плацентарный барьер. Является рецептором предшественников В-лимфоцитов.
№ 61 Антигены: определение, основные свойства. Антигены бактериальной клетки. Антиген – это биополимер органической природы, генетически чужеродный для макроорганизма, который при попадании в последний распознаётся его иммунной системой и вызывает иммунные реакции, направленные на его устранение. Антигены обладают рядом характерных свойств: антигенностью, специфичностью и иммуногенностью. Антигенность. Под антигенностью понимают потенциальную способность молекулы антигена активировать компоненты иммунной системы и специфически взаимодействовать с факторами иммунитета (антитела, клон эффекторных лимфоцитов). Иными словами, антиген должен выступать специфическим раздражителем по отношению к иммунокомпетентным клеткам. При этом взаимодействие компоненты иммунной системы происходит не со всей молекулой одновременно, а только с ее небольшим участком, который получил название «антигенная детерминанта», или «эпитоп». Чужеродность является обязательным условием для реализации антигенности. По этому критерию система приобретенного иммунитета дифференцирует потенциально опасные объекты биологического мира, синтезированные с чужеродной генетической матрицы. Понятие «чужеродность» относительное, так как имму-нокомпетентные клетки не способны напрямую анализировать чужеродный генетический код. Они воспринимают лишь опосредованную информацию, которая, как в зеркале, отражена в молекулярной структуре вещества. Иммуногенность — потенциальная способность антигена вызывать по отношению к себе в макроорганизме специфическую защитную реакцию. Степень иммуногенности зависит от ряда факторов, которые можно объединить в три группы: 1. Молекулярные особенности антигена; 2. Клиренс антигена в организме; 3. Реактивность макроорганизма. К первой группе факторов отнесены природа, химический состав, молекулярный вес, структура и некоторые другие характеристики. Иммуногенность в значительной степени зависит от природы антигена. Важна также оптическая изомерия аминокислот, составляющих молекулу белка. Большое значение имеет размер и молекулярная масса антигена. На степень иммуногенности также оказывает влияние пространственная структура антигена. Оказалась также существенной стерическая стабильность молекулы антигена. Еще одним важным условием иммуногенности является растворимость антигена. Вторая группа факторов связана с динамикой поступления антигена в организм и его выведения. Так, хорошо известна зависимость иммуногенности антигена от способа его введения. На иммунный ответ влияет количество поступающего антигена: чем его больше, тем более выражен иммунный ответ. Третья группа объединяет факторы, определяющие зависимость иммуногенности от состояния макроорганизма. В этой связи на первый план выступают наследственные факторы. Специфичностью называют способность антигена индуцировать иммунный ответ к строго определенному эпитопу. Это свойство обусловлено особенностями формирования иммунного ответа — необходима комплементарность рецепторного аппарата иммунокомпетентных клеток к конкретной антигенной детерминанте. Поэтому специфичность антигена во многом определяется свойствами составляющих его эпитопов. Однако при этом следует учитывать условность границ эпитопов, их структурное разнообразие и гетерогенность клонов антигенреактивных лимфоцитовой специфичности. В результате этого организм на антигенное раздражение всегда отвечает поликлональными иммунным ответом. Антигены бактериальной клетки. В структуре бактериальной клетки различают жгутиковые, соматические, капсульные и некоторые другие антигены. Жгутиковые, или Н-антигены, локализуются в локомоторном аппарате бактерий — их жгутиках. Они представляют собой эпитопы сократительного белка флагеллина. При нагревании флагеллин денатурирует, и Н-антиген теряет свою специфичность. Фенол не действует на этот антиген. Соматический, или О-антиген, связан с клеточной стенкой бактерий. Его основу составляют ЛПС. О-антиген проявляет термостабильные свойства — он не разрушается при длительном кипячении. Однако соматический антиген подвержен действию альдегидов (например, формалина) и спиртов, которые нарушают его структуру. Капсулъные, или К-антигены, располагаются на поверхности клеточной стенки. Встречаются у бактерий, образующих капсулу. Как правило, К-антигены состоят из кислых полисахаридов (уроновые кислоты). В то же время у бациллы сибирской язвы этот антиген построен из полипептидных цепей. По чувствительности к нагреванию различают три типа К-антигена: А, В, и L. Наибольшая термостабильность характерна для типа А, он не денатурирует даже при длительном кипячении. Тип В выдерживает непродолжительное нагревание (около 1 часа) до 60 "С. Тип L быстро разрушается при этой температуре. Поэтому частичное удаление К-антигена возможно путем длительного кипячения бактериальной культуры. На поверхности возбудителя брюшного тифа и других энтеробактерий, которые обладают высокой вирулентностью, можно обнаружить особый вариант капсульного антигена. Он получил название антигена вирулентности, или Vi-антигена. Обнаружение этого антигена или специфичных к нему антител имеет большое диагностическое значение. Антигенными свойствами обладают также бактериальные белковые токсины, ферменты и некоторые другие белки, которые секретируются бактериями в окружающую среду (например, туберкулин). При взаимодействии со специфическими антителами токсины, ферменты и другие биологически активные молекулы бактериального происхождения теряют свою активность. Столбнячный, дифтерийный и ботулинический токсины относятся к числу сильных полноценных антигенов, поэтому их используют для получения анатоксинов для вакцинации людей. В антигенном составе некоторых бактерий выделяется группа антигенов с сильно выраженной иммуногенностью, чья биологическая активность играет ключевую роль в формировании патогенности возбудителя. Связывание таких антигенов специфическими антителами практически полностью инактивирует вирулентные свойства микроорганизма и обеспечивает иммунитет к нему. Описываемые антигены получили название протективных. Впервые протективный антиген был обнаружен в гнойном отделяемом карбункула, вызванного бациллой сибирской язвы. Это вещество является субъединицей белкового токсина, которая ответственна за активацию других, собственно вирулентных субъединиц — так называемого отечного и летального факторов.
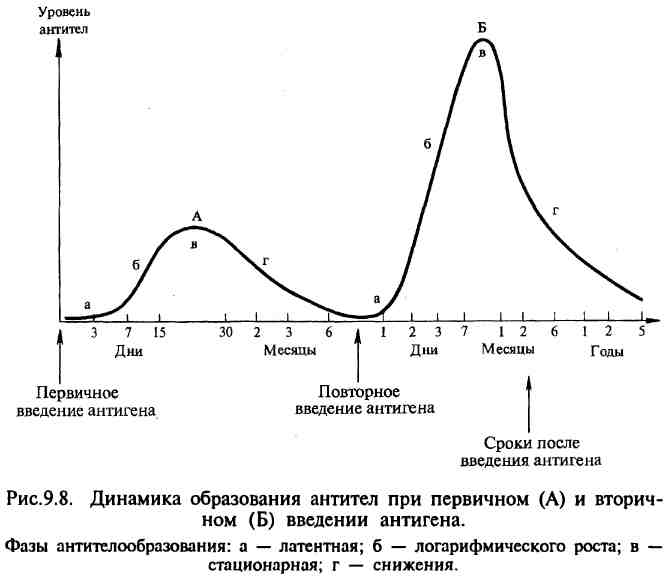
№ 62 Антителообразование: первичный и вторичный ответ.
Способность к образованию антител появляется во внутриутробном периоде у 20-недельного эмбриона; после рождения начинается собственная продукция иммуноглобулинов, которая увеличивается до наступления зрелого возраста и несколько снижается к старости. Динамика образования антител имеет различный характер в зависимости от силы антигенного воздействия (дозы антигена), частоты воздействия антигена, состояния организма и его иммунной системы. При первичном и повторном введении антигена динамика антителообразования также различна и протекает в несколько стадий. Выделяют латентную, логарифмическую, стационарную фазу и фазу снижения.
В латентной фазе происходят переработка и представление антигена иммунокомпетентным клеткам, размножение клона клеток, специализированного на выработку антител к данному антигену, начинается синтез антител. В этот период антитела в крови не обнаруживаются. Во время логарифмической фазы синтезированные антитела высвобождаются из плазмоцитов и поступают в лимфу и кровь. В стационарной фазе количество антител достигает максимума и стабилизируется, затем наступает фаза снижения уровня антител. При первичном введении антигена (первичный иммунный ответ) латентная фаза составляет 3—5 сут, логарифмическая — 7— 15 сут, стационарная — 15—30 сут и фаза снижения — 1—6 мес и более. Особенностью первичного иммунного ответа является то, что первоначально синтезируется IgM, а затем IgG. В отличие от первичного иммунного ответа при вторичном введении антигена (вторичный иммунный ответ) латентный период укорочен до нескольких часов или 1—2 сут, логарифмическая фаза характеризуется быстрым нарастанием и значительно более высоким уровнем антител, который в последующих фазах длительно удерживается и медленно, иногда в течение нескольких лет, снижается. При вторичном иммунном ответе в отличие от первичного синтезируются главным образом IgG. Такое различие динамики антителообразования при первичном и вторичном иммунном ответе объясняется тем, что после первичного введения антигена в иммунной системе формируется клон лимфоцитов, несущих иммунологическую память о данном антигене. После повторной встречи с этим же антигеном клон лимфоцитов с иммунологической памятью быстро размножается и интенсивно включает процесс антителогенеза. Очень быстрое и энергичное антителообразование при повторной встрече с антигеном используется в практических целях при необходимости получения высоких титров антител при производстве диагностических и лечебных сывороток от иммунизированных животных, а также для экстренного создания иммунитета при вакцинации.
№ 63 Иммунологическая память. Иммунологическая толерантность. Иммунологическая память. При повторной встрече с антигеном организм формирует более активную и быструю иммунную реакцию — вторичный иммунный ответ. Этот феномен получил название иммунологической памяти. Иммунологическая память имеет высокую специфичность к конкретному антигену, распространяется как на гуморальное, так и клеточное звено иммунитета и обусловлена В- и Т-лимфоцитами. Она образуется практически всегда и сохраняется годами и даже десятилетиями. Благодаря ней наш организм надежно защищен от повторных антигенных интервенций. На сегодняшний день рассматривают два наиболее вероятных механизма формирования иммунологической памяти. Один из них предполагает длительное сохранение антигена в организме. Этому имеется множество примеров: инкапсулированный возбудитель туберкулеза, персистирующие вирусы кори, полиомиелита, ветряной оспы и некоторые другие патогены длительное время, иногда всю жизнь, сохраняются в организме, поддерживая в напряжении иммунную систему. Вероятно также наличие долгоживущих дендритных АПК, способных длительно сохранять и презентировать антиген. Другой механизм предусматривает, что в процессе развития в организме продуктивного иммунного ответа часть антигенореактивных Т- или В-лимфоцитов дифференцируется в малые покоящиеся клетки, или клетки иммунологической памяти. Эти клетки отличаются высокой специфичностью к конкретной антигенной детерминанте и большой продолжительностью жизни (до 10 лет и более). Они активно рециркулируют в организме, распределяясь в тканях и органах, но постоянно возвращаются в места своего происхождения за счет хоминговых рецепторов. Это обеспечивает постоянную готовность иммунной системы реагировать на повторный контакт с антигеном по вторичному типу. Феномен иммунологической памяти широко используется в практике вакцинации людей для создания напряженного иммунитета и поддержания его длительное время на защитном уровне. Осуществляют это 2—3-кратными прививками при первичной вакцинации и периодическими повторными введениями вакцинного препарата — ревакцинациями. Однако феномен иммунологической памяти имеет и отрицательные стороны. Например, повторная попытка трансплантировать уже однажды отторгнутую ткань вызывает быструю и бурную реакцию — криз отторжения. Иммунологическая толерантность — явление, противоположное иммунному ответу и иммунологической памяти. Проявляется она отсутствием специфического продуктивного иммунного ответа организма на антиген в связи с неспособностью его распознавания. В отличие от иммуносупрессии иммунологическая толерантность предполагает изначальную ареактивность иммунокомпетентных клеток к определенному антигену. Иммунологическую толерантность вызывают антигены, которые получили название толерогены. Ими могут быть практически все вещества, однако наибольшей толерогенностью обладают полисахариды. Иммунологическая толерантность бывает врожденной и приобретенной. Примером врожденной толерантности является отсутствие реакции иммунной системы на свои собственные антигены. Приобретенную толерантность можно создать, вводя в организм вещества, подавляющие иммунитет (иммунодепрессанты), или же путем введения антигена в эмбриональном периоде или в первые дни после рождения индивидуума. Приобретенная толерантность может быть активной и пассивной. Активная толерантность создается путем введения в организм толерогена, который формирует специфическую толерантность. Пассивную толерантность можно вызвать веществами, тормозящими биосинтетическую или пролиферативную активность иммунокомпетентных клеток (антилимфоцитарная сыворотка, цитостатики и пр.). Иммунологическая толерантность отличается специфичностью — она направлена к строго определенным антигенам. По степени распространенности различают поливалентную и расщепленную толерантность. Поливалентная толерантность возникает одновременно на все антигенные детерминанты, входящие в состав конкретного антигена. Для расщепленной, или моновалентной, толерантности характерна избирательная невосприимчивость каких-то отдельных антигенных детерминант. Степень проявления иммунологической толерантности существенно зависит от ряда свойств макроорганизма и толерогена. Важное значение в индукции иммунологической толерантности имеют доза антигена и продолжительность его воздействия. Различают высокодозовую и низкодозовую толерантность. Высокодозовую толерантность вызывают введением больших количеств высококонцентрированного антигена. Низкодозовая толерантность, наоборот, вызывается очень малым количеством вы-сокогомогенного молекулярного антигена. Механизмы толерантности многообразны и до конца не расшифрованы. Известно, что ее основу составляют нормальные процессы регуляции иммунной системы. Выделяют три наиболее вероятные причины развития иммунологической толерантности: 1. Элиминация из организма антигенспецифических клонов лимфоцитов. 2. Блокада биологической активности им-мунокомпетентных клеток. 3. Быстрая нейтрализация антигена антителами. Феномен иммунологической толерантности имеет большое практическое значение. Он используется для решения многих важных проблем медицины, таких как пересадка органов и тканей, подавление аутоиммунных реакций, лечение аллергий и других патологических состояний, связанных с агрессивным поведением иммунной системы.
№ 64 Классификация гиперчувствительности по Джейлу и Кумбсу.
Изучение молекулярных механизмов аллергии привело к созданию Джеллом и Кумбсом в 1968 г. новой классификации. В соответствии с ней различают четыре основных типа аллергии: анафилактический (I тип), цитотоксический (II тип), иммунокомплексный (III тип) и опосредованный клетками (IV тип). Первые три типа относятся к ГНТ, четвертый — к ГЗТ. Ведущая роль в запуске ГНТ играют антитела (IgE, G и М), а ГЗТ — лимфоидно-макрофагальная реакция. Аллергическая реакция I типа связана с биологическими эффектами IgE и G4, названных реагинами, которые обладают цитофильностью — сродством к тучным клеткам и базофилам. Эти клетки несут на поверхности высокоаффинный FcR, связывающий IgE и G4 и использующий их как ко-рецепторный фактор специфического взаимодействия с эпитопом аллергена. Связывание аллергена с рецепторным комплексом вызывает дегрануляцию базофила и тучной клетки — залповый выброс биологически активных соединений (гистамин, гепарин и др.), содержащихся в гранулах, в межклеточное пространство. В результате развиваются бронхоспазм, вазодилатация, отек и прочие симптомы, характерные для анафилаксии. Вырабатываемые цитокины стимулируют клеточное звено иммунитета: образование Т2-хелпера и эозинофилогенез. Цитотоксические антитела (IgG, IgM), направленные против поверхностных структур (антигенов) соматических клеток макроорганизма, связываются с клеточными мембранами клеток-мишеней и запускают различные механизмы антителозависимой цитотоксичности (аллергическая реакция II типа). Массивный цитолиз сопровождается соответствующими клиническими проявлениями. Классическим примером является гемолитическая болезнь в результате резус-конфликта или переливания иногруппной крови. Цитотоксическим действием обладают также комплексы атиген—антитело, образующиеся в организме пациента в большом количестве после введения массивной дозы антигена (аллергическая реакция III типа). В связи с кумулятивным эффектом клиническая симптоматика аллергической реакции III типа имеет отсроченную манифестацию, иногда на срок более 7 суток. Тем не менее этот тип реакции относят к ГНТ. Реакция может проявляться как одно из осложнений от применения иммунных гетерологичных сывороток с лечебно-профилактической целью («сывороточная болезнь»), а также при вдыхании белковой пыли («легкое фермера»). Лабораторная диагностика аллергии при аллергических реакциях I типа основана на выявлении суммарных и специфических реагинов (IgE, IgG4) в сыворотке крови пациента. При аллергических реакциях II типа в сыворотке крови определяют цитотоксические антитела (антиэритроцитарные, антилейкоцитарные, антитромбоцитарные и др.). При аллергических реакциях III типа в сыворотке крови выявляют иммунные комплексы. Для обнаружения аллергических реакций IV типа применяют кожно-аллергические пробы, которые широко используют в диагностике некоторых инфекционных и паразитарных заболеваний и микозов (туберкулез, лепра, бруцеллез, туляремия и др.).
№ 65 Механизмы гиперчувствительности замедленного типа. Клинико-диагностическое значение.
К аллергическим реакциям относят два типа реагирования на чужеродное вещество: гиперчувствительность немедленного типа (ГНТ) и гиперчувствительность замедленного типа (ГЗТ). К ГНТ относятся аллергические реакции, проявляющиеся уже через 20—30 мин после повторной встречи с антигеном, а к ГЗТ — реакции, возникающие не ранее чем через 24—48 ч. Механизм и клинические проявления ГНТ и ГЗТ различны. ГНТ связана с выработкой антител, а ГЗТ — с клеточными реакциями. ГЗТ впервые описана Р. Кохом (1890). Эта форма проявления не связана с антителами, опосредована клеточными механизмами с участием Т-лимфоцитов. К ГЗТ относятся следующие формы проявления: туберкулиновая реакция, замедленная аллергия к белкам, контактная аллергия. В отличие от реакций I, II и III типов реакции IV типа не связаны с антителами, а обусловлены клеточными реакциями, прежде всего Т-лимфоцитами. Реакции замедленного типа могут возникать при сенсибилизации организма: 1. Микроорганизмами и микро Дата добавления: 2015-02-06 | Просмотры: 1192 | Нарушение авторских прав |