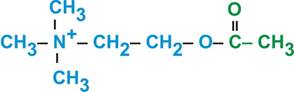|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Физиология синапсов: строение, классификация и механизмы деятельности. Медиаторы, нейрохимические основы поведения
В патогенезе большую роль играют: Дискинезия желчных путей Паразитарная инвазия Другая патология органов пищеварения Очаги хронической инфекции Обменные нарушения Частые вирусные инфекции и т.д.
Критерии диагностики хронического холецистита
Анамнестические 1. Постепенное начало заболевания 2. Хроническое или волнообразное течение 3. Наследственная предрасположенность 4. Связь заболевания с аномалией развития ЖП, инфекцией, гельминтозом (описторхоз), вегетативной дисфункцией
Клинические 1. Боль в правом подреберье (тупая, сильная, приступообразная) 2. Связь боли с погрешностью в диете, физической нагрузкой, интеркуррентными заболеваниями 3. Иррадиация боли в правое плечо, лопатку 4.Симптомы интоксикации 5. Лихорадка при обострении заболевания 6. Мышечное напряжение в правом подреберье 7. Положительные пузырные симптомы (Кера, Ортнера, мерфи и др.) 8. Возможно увеличение печени
Лабораторные 1 Признаки воспаления в периферической крови 2. При биохимическом исследовании крови: диспротеинемия, увеличение щелочной фосфатазы, фибриногена, липопротеидов, билирубина, аминотрансфераз 3. При исследовании желчи: слизь, лейкоциты, кристаллы холестерина, микробы; уменьшение билирубина, желчных кислот, щелочной фосфатазы
Инструментальные 1. Эхография (уплотнение, утолщение стенок ЖП, повышение эхогенности желчи, наличие хлопье слизи) 2. Холецистография (деформация ЖП, ослаб-ление концентрационной способности 2-4 степени) 3. Панкреатохолангиография (рубцовые стрик-туры и недостаточность большого дуоде-нального соска
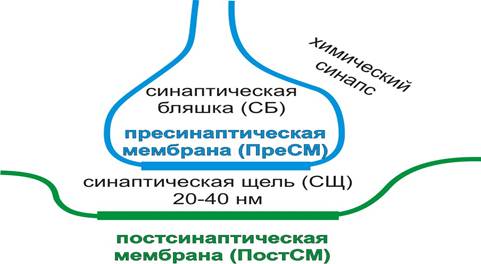
Физиология синапсов: строение, классификация и механизмы деятельности. Медиаторы, нейрохимические основы поведения. Слайд 1 В конце XIX века параллельно существовали две теории организации нервной системы (НС). Ретикулярная теория считала, что НС представляет собой функциональный синцитий: нейроны соединены с помощью отростков подобно капиллярам кровеносной системы. Согласно клеточной теории Вальдейера (1981 г.) НС состоит из отдельных, разделенных мембранами, нейронов. Чтобы решить вопрос о взаимодействии отдельных нейронов, Шеррингтон в 1987 г. предположил наличие особого мембранного образования – синапса. С помощью электронного микроскопа наличие синапсов было безоговорочно подтверждено. Однако, клеточная теория строения НС стала общепризнанной, по иронии науки в 1959 г. Фершпан и Поттер открыли в НС ракообразных синапс с щелевыми контактами (электрический синапс). Синапс – это мембранное образование двух (или более) клеток, в котором происходит передача возбуждения (информации) от одной клетки к другой. Существует следующая классификация синапсов: 1) по механизму передачи возбуждения (и по строению): - химические; - электрические (эфапсы); - смешанные. 2) по выделяемому нейромедиатору: - адренергические – нейромедиатор норадреналин; - холинергические – нейромедиатор ацетилхолин; - дофаминергические – нейромедиатор дофамин; - серотонинергические – нейромедиатор серотонин; - ГАМК-ергические – нейромедиатор гамма-аминомасляная кислота (ГАМК) 3) по влиянию: - возбуждающие; - тормозные. 4) по местоположению: - нервно-мышечные; - нейро-нейрональные: а) аксо-соматические; б) аксо-аксональные; в) аксо-дендрические; г) дендросоматические. Рассмотрим три типа синапсов: химический, электрический и смешанный (совмещающий свойства химического и электрического синапсов). Независимо от типа, синапсы имеют общее черты строения: нервный отросток на конце образует расширение (синаптическую бляшку, СБ); конечная мембрана СБ отлична от других участков мембраны нейрона и носит название пресинаптической мембраны (ПреСМ); специализированная мембрана второй клетки обозначается постсинаптической мембраной (ПостСМ); между мембранами синапса находится синаптическая щель (СЩ, рис. 1, 2).
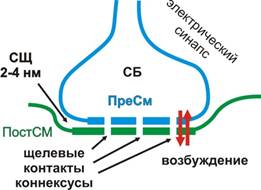
Рис. 1. Схема строения химического синапса Электрические синапсы (эфапсы, ЭС) сегодня обнаружены в НС не только ракообразных, но и моллюсков, членистоногих, млекопитающих. ЭС обладают рядом уникальных свойств. Они имеют узкую синаптическую щель (около 2-4 нм), благодаря чему возбуждение может передаваться электрохимическим способом (как по нервному волокну за счет ЭДС) с высокой скоростью и в обоих направлениях: как от ПреСМ мембраны к ПостСМ, так и от ПостСМ к ПреСМ. Между клетками имеются щелевые контакты (коннексусы или коннексоны), образованные двумя белками коннексинами. Шесть субъединиц каждого коннексина формируют каналы ПреСМ и ПостСМ, через которые клетки могут обмениваться низкомолекулярными веществами молекулярной массой 1000-2000 Дальтон. Работа коннексонов может регулироваться ионами Са2+(рис. 2).
Рис. 2. Схема электрического синапса
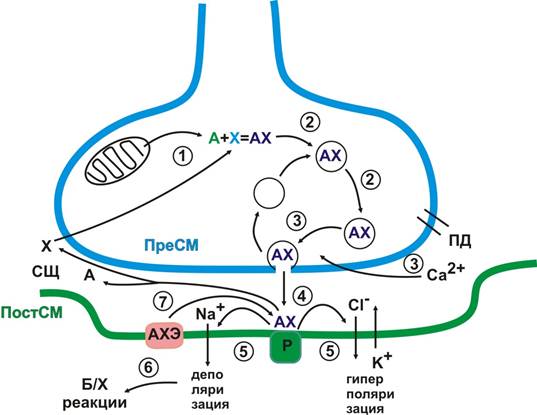
ЭС обладают большей специализацией по сравнению с химическими синапсами и обеспечивают высокую скорость передачи возбуждения. Однако он, по-видимому, лишен возможности более тонкого анализа (регуляции) передаваемой информации. Химические синапсы доминируют в НС. История их изучения начинается с работ Клода Бернара, который в 1850 г. опубликовал статью «Исследование о кураре». Вот что он писал: «Кураре – сильный яд, приготовляемый некоторыми народностями (большей частью людоедами), обитающими в лесах... Амазонки». И далее, «Кураре сходен с ядом змеи в том отношении, что его можно безнаказанно ввести в пищеварительный тракт человека или животных, в то время как впрыскивание его под кожу или в какую-либо часть тела быстро приводит к смерти. …через несколько мгновений животных ложатся, как будто они устали. Затем дыхание останавливается и их чувствительность и жизнь исчезают, причем животные не издают крика и не проявляют никаких признаков боли». Хотя К.Бернар не пришел к мысли о химической передачи нервного импульса, его классические опыты с кураре позволили этой мысли зародиться. Прошло более полувека, когда Дж. Ленгли установил (1906 г.), что парализующее действие кураре связано с особой частью мышцы, которую он назвал рецептивной субстанцией. Впервые предположение о передаче возбуждения с нерва на эффекторный орган с помощью химического вещества было высказано Т. Элиотом (1904). Однако окончательно утвердили гипотезу химического синапса только работы Г. Дейла и О. Лёви. Дейл в 1914 г. установил, что раздражение парасимпатического нерва имитируется ацетилхолином. Лёви в 1921 г. доказал, что ацетилхолин выделяется из нервного окончания блуждающего нерва, а в 1926 г. открыл ацетилхолинэстеразу – фермент, разрушающий ацетилхолин. Возбуждение в химическом синапсе передается с помощью медиатора. Этот процесс включает в себя несколько стадий. Рассмотрим эти особенности на примере ацетилхолинового синапса, который широко распространении в ЦНС, вегетативной и периферической нервной системе (рис. 3).
Рис. 3. Схема функционирования химического синапса 1. Медиатор ацетилхолин (АХ) синтезируется в синаптической бляшке из ацетил-СоА (ацетил-кофермент А образуется в митохондриях) и холина (синтезируется печенью) с помощью ацетилхолинтрансферазы (рис. 3, 1). 2. Медиатор упакован в синаптические везикулы ( Кастильо, Катц; 1955 г.). Количество медиатора в одной везикуле составляет несколько тысяч молекул (квант медиатора). Часть везикул расположена на ПреСМ и готова к высвобождению медиатора (рис. 3, 2). 3. Высвобождается медиатор путем экзоцитоза при возбуждении ПреСМ. Важную роль в разрыве мембран и квантовом высвобождении медиатора играет входящий ток Са2+ (рис. 3, 3). 4. Высвободившийся медиатор связывается со специфическим белком-рецептором ПостСМ (рис. 3, 4). 5. В результате взаимодействия медиатора и рецептора изменяется ионная проводимость ПостСМ: при открытии Na+ каналов происходит деполяризации; открытие K+ или Cl- каналов приводит к гиперполяризации (рис. 3, 5). 6. Вслед за деполяризацией запускаются биохимические процессы в постсинаптической цитоплазме (рис. 3, 6). 7. Рецептор освобождается от медиатора: АХ разрушается ацетилхолинэстеразой (АХЭ, рис. 3. 7). Начало формы
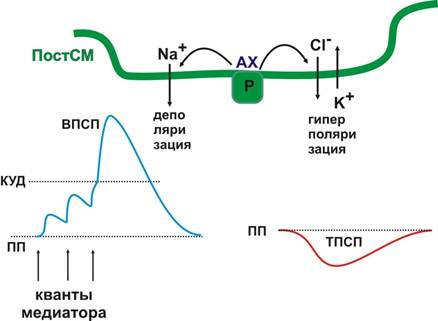
братите внимание, что медиатор в норме взаимодействует со специфическим рецептором с определенной силой и длительностью. Почему кураре - яд? Местом действия кураре как раз является АХ синапс. Кураре более прочно связывается с ацетилхолиновым рецептором и лишает его взаимодействия с медиатором (АХ). Возбуждение с соматических нервов на скелетные мышцы, в том числе с диафрагмального нерва на основную дыхательную мышцу (диафрагму) передается с помощью АХ, поэтому кураре вызывает релаксацию (расслабление) мышц и остановку дыхания (из-за чего, собственно, и наступает смерть). Отметим основные особенности передачи возбуждения в химическом синапсе. 1. Возбуждение передается с помощью химического посредника – медиатора. 2. Возбуждение передается в одном направлении: от ПреСм к ПостСМ. 3. В химическом синапсе происходит временная задержка в проведении возбуждения, поэтому синапс обладает низкой лабильностью. 4. Химический синапс обладает высокой чувствительностью к действию не только медиаторов, но и других биологически активных веществ, лекарств и ядов. 5. В химическом синапсе происходит трансформация возбуждений: электрохимическая природа возбуждения на ПреСМ продолжается в биохимический процесс экзоцитоза синаптических везикул и связывания медиатора со специфическим рецептором. За этим следует изменение ионной проводимости ПостСМ (тоже электрохимический процесс), который продолжается биохимическими реакциями в постсинаптической цитоплазме. В принципе, такая многостадийность передачи возбуждения должна иметь весомое биологическое значения. Обратите внимание, что на каждом из этапов возможна регуляция процесса передачи возбуждения. Несмотря на ограниченное количество медиаторов (чуть больше десятка), в химическом синапсе имеются условия для широкого разнообразия в решении судьбы приходящего в синапс нервного возбуждения. Совокупность особенностей химических синапсов объясняет индивидуальное биохимическое разнообразие нервных и психических процессов. Теперь остановимся на двух важных процессах, протекающих в постсинаптическом пространстве. Мы отметили, что в результате взаимодействия АХ с рецептором на ПостСМ могут развиваться как деполяризация, так и гиперполяризация. От чего же зависит, будет ли медиатор возбуждающим или тормозным? Результат взаимодействия медиатора и рецептора определяется свойствами рецепторного белка (еще одно важное свойство химического синапса – ПостСМ активна по отношению к приходящему к ней возбуждению). В принципе химический синапс – динамическое образование, изменяя рецептор, клетка, принимающая возбуждение, может влиять на его дальнейшую судьбу. Если свойства рецептора таковы, что его взаимодействие с медиатором открывает Na+каналы, то при выделении одного кванта медиатора на ПостСМ развивается локальный потенциал (для нервно-мышечного синапса он носит название миниатюрного потенциала концевой пластинки – МПКП). Когда же возникает ПД? Возбуждение ПостСМ (возбуждающий постсинаптический потенциал – ВПСП) возникает как результат суммации локальных потенциалов. Можно выделить два типа суммационных процессов. При последовательном выделении нескольких квантов медиатора в одном и том же синапсе (вода и камень точит) возникает временн а я суммация. Если кванты медиаторы выделяются одновременно в разных синапсах (на мембране нейрона их может быть несколько тысяч) возникает пространственная суммация. Реполяризация ПостСМ мембраны происходит медленно и после выделения отдельных квантов медиатора ПостСМ некоторое время находится в состоянии экзальтации (так называемая синаптическая потенциация, рис. 4). Возможно, таким образом, происходит обучение синапса (выделение квантов медиатора в определенных синапсах могут «подготовить» мембрану к решающему взаимодействию с медиатором). При открытии K+ или Cl- каналов на ПостСМ возникает тормозный постсинаптический потенциал (ТПСП, рис. 4).
Рис. 4. Потенциалы постсинаптической мембраны Естественно, что в случае развития ТПСП дальнейшее распространение возбуждения может быть остановлено. Другой вариант прекращения процесса возбуждения – пресинаптическое торможение. Если на мембране синаптической бляшки образуется тормозный синапс, то в результате гиперполяризации ПреСМ экзоцитоз синаптических визикул может быть заблокирован. Второй важный процесс – развитие биохимических реакций в постсинаптической цитоплазме. Изменение ионной проводимости ПостСМ активирует так называемые вторичные мессенджеры (посредники): цАМФ, цГМФ, Са2+-зависимую протеинкиназу, которые, в свою очередь активируют различные протеинкиназы путем их фосфорилирования. Эти биохимические реакции могут «спускаться» вглубь цитоплазмы вплоть до ядра нейрона, регулируя процессы белкового синтеза. Таким образом, нервная клетка может ответить на пришедшее возбуждение не только решением его дальнейшим судьбы (ответить ВПСП или ТПСП, т.е. провести или не провести далее), а изменить количество рецепторов, или синтезировать белок-рецептор с новыми свойствами по отношению к определенному медиатору. Следовательно, еще одно важное свойство химического синапса: благодаря биохимическим процессам постсинаптической цитоплазмы клетка готовится (обучается) к будущим взаимодействиям. В нервной системе функционируют разнообразные синапсы, которые отличаются медиаторами и рецепторами. Название синапсов определяется медиатором, точнее названием рецептора к конкретному медиатору. Поэтому, рассмотрим классификацию основных медиаторов и рецепторов нервной системы (смотрите так же материал, розданный на лекции!!). Мы уже отмечали, что эффект взаимодействия медиатора и рецептора определяется свойствами рецептора. Поэтому известные медиаторы, за исключением g-аминомасляной кислоты, могут выполнять функции как возбуждающих, так и тормозных медиаторов.По химической структуре выделяют следующие группы медиаторов. Ацетилхолин, широко распространен в ЦНС, является медиатором в холинергических синапсах вегетативной нервной системы, а также в соматических нервно-мышечных синапсах (рис. 5).
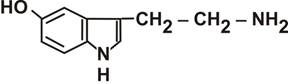
Рис. 5. Молекула ацетилхолина Известны два типа холинорецепторов: никотиновые (Н-холинорецепторы) и мускариновые (М-холинорецепторы). Название получили по веществам, вызывающим сходный с ацетилхолином эффект в этих синапсах: Н-холиномиметиком является никотин, а М-холиномиметиком - токсин мухомора Amanita muscaria (мускарин). Блокатором (холинолитиком) Н-холинорецептора является d-тубокурарин (основной компонент яда кураре), а М-холинолитиком является токсин красавки Atropa belladonna – атропин. Интересно, что свойства атропина давно известно и было время, когда женщины использовали атропин красавки, чтобы вызвать расширение зрительных зрачков (сделать глаза темными и «красивыми»). Четыре следующих основных медиаторов имеют сходство в химической структуре, поэтому их относят к группе моноаминов. Это серотонин или 5-гидрокситриптами (5-HT), играет важную роль в механизмах подкрепления (гормон радости). Синтезируется из незаменимой для человека аминокислоты – триптофана (рис. 6).
Рис. 6. Молекула серотонина (5-гидрокситриптамина) Три других медиатора синтезируются из незаменимой аминокислоты фенилаланина, поэтому объединены общим названием катехоламинов – это дофамин (допамин), норадреналин (норэпинефрин) и адреналин (эпинефрин, рис. 7).
Рис. 7. Катехоламины
Среди аминокислот к медиаторам относят гамма-аминомасляную кислоту (g-АМК или ГАМК – известна как только тормозный медиатор), глицин, глутаминовую кислоту, аспарагиновую кислоту. К медиаторам относят ряд пептидов. В 1931 г. Эйлером в экстрактах мозга и кишечника было обнаружено вещество, вызывающее сокращение гладких мышц кишечника, расширение кровеносных сосудов. Этот медиатор был в чистом виде выделен из гипоталамуса и получил название вещества Р (от англ. powder – порошок, состоит из 11 аминокислот). В дальнейшем установлено, что вещество Р играет важную роль в проведении болевых возбуждений (название не пришлось менять, т.к. боль по англ. - pain). Пептид дельта сна получил свое название за способность вызывать в электроэнцефалограмме медленные высокоамплитудные ритмы (дельта-ритмы). В мозге синтезируется целый ряд белковых медиаторов наркотической (опиатной) природы. Это пентапептиды Met-энкефалин и Leu-энкефалин, а также эндорфины. Это важнейшие блокаторы болевых возбуждений и медиаторы подкрепления (радости и удовольствия). Другими словами, наш мозг является отличной фабрикой эндогенных наркотиков. Главное, научить мозг их вырабатывать. «Как?» - спросите вы. Все просто – эндогенные опиаты вырабатываются, когда мы получаем удовольствие. Делайте все с удовольствием, заставляйте свою эндогенную фабрику синтезировать опиаты! Нам от природы с рождения дана эта возможность – подавляющее большинство нейронов реактивны на положительное подкрепление. Исследования последних десятилетий позволили открыть еще один очень интересный медиатор – оксид азота (NO). Оказалось, что NO не только играет важную роль в регуляции тонуса кровеносных сосудов (известный вам нитроглицерин является источником NO и расширяет коронарные сосуды), но и синтезируется в нейронах ЦНС. В принципе, история медиаторов еще не закончена, есть целый ряд веществ, которые участвуют в регуляции нервного возбуждения. Просто пока точно не установлен факт их синтеза в нейронах, они не обнаружены в синаптических везикулах, не найдены специфические к ним рецепторы. Конец формы
Дата добавления: 2014-09-03 | Просмотры: 4125 | Нарушение авторских прав |