|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Нуклеиновые кислоты как генетический материал
В предыдущем разделе была показана роль хромосом в передаче наследственности. Вопрос о том, из каких компонентов состоят хромосомы, встал после того, как выяснилось, что они являются материальной основой наследственности. Установлено, что хромосомы состоят в основном из нуклеиновых кислот и гистоновых белков. С помощью методов окрашивания доказано, что во время деления клетки большая часть белков отделяется от хромосом, тогда как нуклеиновые кислоты сохраняются. К настоящему времени наука располагает достаточным количеством убедительных данных, свидетельствующих о том, что генетическая информация хранится в молекулах нуклеиновых кислот и передается ими по наследству. У всех клеточных организмов и у ДНК-содержащих вирусов это осуществляется молекулами ДНК (дезоксирибонуклеиновой кислоты), а у РНК-содержащих вирусов – молекулами РНК (рибонуклеиновой кислоты). Молекулы ДНК и РНК имеют и общие, и отличительные признаки в структуре. В состав ДНК входит сахар дезоксирибоза и четыре типа азотистых оснований: пурины – аденин и гуанин, пиримидины – цитозин и тимин. Химическим остовом (скелетом) ДНК служат остатки сахара, чередующиеся с остатками фосфорной кислоты. Каждый остаток фосфорной кислоты связан с 5I углеродом одного остатка сахара и с 3I углеродом другого остатка сахара (рис. 32). К первому углероду каждого остатка сахара сбоку присоединено какое-нибудь из азотистых оснований.
Ф – остаток фосфата, Д – дезоксирибоза, П – пуриновое или пиримидиновое основание
Два сахарофосфатных остова удерживаются водородными связями соединенных попарно оснований.
В расшифровке структуры молекулы ДНК ключевую роль сыграли исследования Чаргаффа, который изучал соотношение различных нуклеотидов в ДНК. Нуклеотид состоит из сахара, фосфорного остатка и азотистого основания (рис. 33). Чаргафф обнаружил, что молярное содержание аденина примерно равно содержанию тимина, а гуанина – цитозину. Из этого следует, что на каждый остаток тимина в ДНК приходится один остаток аденина, на каждый остаток цитозина – один остаток гуанина. Рис. 33. Строение отдельного нуклеотида ДНК
Из этого правила и физических параметров молекулы вытекает, что каждая пара оснований должна состоять из одного пурина и одного пиримидина. Учитывая молекулярную совместимость (способность образовывать нужные водородные связи), можно сказать, что такими парами должны быть аденин – тимин и гуанин – цитозин. Структуры этих пар оснований приведены на рис. 34, а схема отрезка двухнитчатой молекулы Рис. 34. Возможные пары оснований в ДНК
Рис. 35. Двойная спираль ДНК Таким образом, каждая нить ДНК является полинуклеотидом, так как состоит из многих нуклеотидов. Такая структура не предусматривает никаких ограничений в отношении последовательности оснований вдоль каждой цепи. Оно может быть любым. С теоретической точки зрения существует неограниченное число возможных последовательностей нуклеотидов. Специфичность разных ДНК обусловлена последовательностью пар оснований в молекуле. Однако в силу специфичности спаривания оснований наличие известной последовательности одной цепи предопределяет последовательность оснований другой цепи. Эта особенность структуры ДНК получила название комплементарности. Благодаря такому комплементарному строению молекула ДНК способна точно воспроизводить себя. Каждая из двух нитей может служить матрицей для образования другой комплементарной нити. Для этого молекулы должны раскрутиться, легкие водородные связи при этом рвутся. Каждое пуриновое и пиримидиновое основание притягивает к себе комплементарный свободный нуклеотид и удерживает его на месте с помощью специфических водородных связей (рис. 36). Как только свободные нуклеотиды закрепляются на родительской матричной цепи, они сшиваются вместе в результате образования фосфодиэфирных связей между соседними остатками дезоксирибозы, образуя новую полинуклеотидную молекулу с предопределенной последовательностью оснований. В результате образуются две молекулы ДНК, имеющие идентичные последовательности четырех оснований и, следовательно, то же информационное содержание, что и родительская двухспиральная ДНК.
 Рис. 36. Репликация ДНК по механизму Дж. Уотсона и Ф. Крика Рис. 36. Репликация ДНК по механизму Дж. Уотсона и Ф. Крика
пределения вещества наследственности никто не мог представить себе до открытия комплементарной двойной спирали. Уотсон и Крик оказались правыми в решении вопроса о структуре ДНК, но это не значит, что они правы и в вопросе о способе репликации. Они предполагали, что новые одноцепочные спирали синтезируются одновременно, как показано на рисун-ке 36. Однако все известные ферменты, которые катализируют синтез ДНК in vitro, работают только в направлении 5I 3I. Таким образом, одна цепь должна синтезироваться в направлении репликационной вилки, а другая – от нее, как показано на рис. 37.
Рис. 37. Прерывистый синтез ДНК
Эта модель прерывистого синтеза ДНК была предложена в 1968 г. на основании экспериментов японского ученого Р. Оказаки, проведенных на Е.coli. В настоящее время имеются убедительные данные о том, что и у прокариот, и у эукариот синтез в направлении от вилки идет путем образования фрагментов, которые соединяются под действием ДНК-лигазы. Синтез ДНК в направлении репликационной вилки идет, по-видимому, непрерывно. В репликации ДНК участвуют различные ферменты (белки). Расплетающий фермент разделяет цепи ДНК в области репликационной вилки, а белок, дестабилизирующий двойную спираль, удерживает их в расплетенном состоянии, ДНК-полимераза отвечает за синтез ДНК при репликации. После расшифровки структуры и механизма репликации ДНК стало возможным объяснить, каким образом происходят ошибки копирования. Напомним, что согласно схеме репликации, две комплементарные полинуклеотидные цепи родительской молекулы ДНК расходятся, и каждая из них служит матрицей для синтеза новой комплементарной цепи. Этот синтез протекает путем образования специфических водородных связей между пуриновыми и пиримидиновыми основаниями родительской полинуклеотидной цепи, с одной стороны, и мононуклеотидными «строительными блоками», присутствующими во внутриклеточном метаболическом фонде – с другой. Для образования специфических водородных связей между аденином и тимином и между гуанином и цитозином необходимо, чтобы эти основания находились в особой таутомерной форме (рис. 34). В действительности в любой водной среде каждое из этих оснований существует в нескольких различных таутомерных формах. Формы, приведенные на рис. 34, наиболее вероятны. В некоторых других формах, менее вероятных, атом водорода, участвующий в образовании водородной связи, занимает иное положение. Вследствие этого специфическая связь образуется не с тем основанием, с которым данное основание спаривается, а уже с другим. Например, аденин в редкой иминной форме может спариваться с цитозином вместо тимина, а тимин в редкой енольной форме спаривается с гуанином вместо аденина (рис. 38). Такое спаривание оснований ведет к включению «неправильного» нуклеотида в синтезируемую нить ДНК. Результатом этого оказывается устойчивое изменение последовательности нуклеотидов, или мутация. Этот молекулярный механизм мутаций предложен также Дж. Уотсоном и Ф. Криком. Он положен в основу современных представлений о процессе мутирования.
Рис. 38. «Незаконное» спаривание редкой иминоформы аденина с цитозином и редкой енольной формы тимина с гуанином Другая нуклеиновая кислота (РНК) представляет собой полинуклеотидную цепь, напоминающую ДНК. Основное химическое различие между ДНК и РНК состоит в том, что в РНК углеродный остов нуклеотидной цепи построен из остатков рибозы, а не дезоксирибозы, как в ДНК и, кроме того, в РНК одним из четырех азотистых оснований служит не тимин, а другой пиримидин – урацил. Оказалось, что эти два довольно незначительных химических различия (дополнительная гидроксильная группа в молекуле сахара РНК и дополнительная метильная группа в одном из пиримидинов ДНК) имеют чрезвычайно большое значение для биологической роли, выполняемой этими двумя полинуклеотидами. Молекулы РНК, как правило, однонитчатые. По величине они разные, но в общем они меньше молекул ДНК. Только в РНК-содержащих вирусах молекулы эти двухнитчатые, построенные как ДНК. Такие РНК служат хранителями наследственной информации и передают ее следующему поколению. Все остальные РНК разделены на несколько типов в зависимости от их функций. Все типы РНК синтезируются на ДНК-матрицах. Матричный синтез РНК катализируется РНК-полимеразами. Итак, генетический материал у всех живых организмов представлен нуклеиновыми кислотами, и в основном ДНК. Содержащаяся в ДНК генетическая информация записана в виде определенной последовательности нуклеотидов, стереотипно воспроизводящихся при клеточных делениях. Синтез молекулы ДНК осуществляется с участием многих ферментов, но отличительной чертой этого синтеза является использование в качестве матрицы предшествующей последовательности нуклеотидов, материально передаваемой от клетки к клетке. В том случае, когда генетический материал представлен РНК, состоящей из одинарной спирали нуклеотидов, ее репродукция осуществляется по тому же принципу: матрицей служит двухцепочечная репликативная форма РНК, образующаяся в клетках после инфицирования их вирусами. Знания эти способствуют конкретизации представления о гене. В классической генетике словом «ген» обозначали единицу генетического материала, которая передается от родителей к потомству, и может быть обнаружена в эксперименте по ее способности мутировать в различные состояния, рекомбинировать с другими такими же единицами и функционировать, наделяя организм каким-либо конкретным фенотипом. Эти три проявления – мутация, рекомбинация и функция – относили к одной и той же наследственной единице – гену. Сейчас установлено, что единицей мутации и рекомбинации является отдельная пара нуклеотидов в ДНК, единицей функции служит последовательность сотен или тысяч нуклеотидов, которые детерминируют последовательность аминокислот в белке. Дата добавления: 2015-01-18 | Просмотры: 1376 | Нарушение авторских прав |
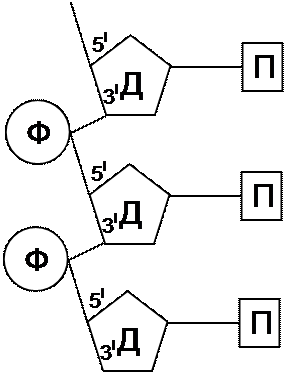
 Структуру ДНК расшифровали Дж. Уотсон и Б. Крик в 1953 году. Любая модель ДНК должна учитывать биологические свойства вещества наследственности: точное удвоение, способность хранить генетическую информацию и потенциальную способность к мутациям. Оригинальное сочетание химических методов и методов рентгеноструктурного анализа позволило установить, что молекула ДНК представляет собой двойную спираль, состоящую из антипараллельных полуспиралей: в одной из них остатки сахара соединяются фосфатными группами в направлении 3I 5I, в другой в направлении 5I 3I.
Структуру ДНК расшифровали Дж. Уотсон и Б. Крик в 1953 году. Любая модель ДНК должна учитывать биологические свойства вещества наследственности: точное удвоение, способность хранить генетическую информацию и потенциальную способность к мутациям. Оригинальное сочетание химических методов и методов рентгеноструктурного анализа позволило установить, что молекула ДНК представляет собой двойную спираль, состоящую из антипараллельных полуспиралей: в одной из них остатки сахара соединяются фосфатными группами в направлении 3I 5I, в другой в направлении 5I 3I.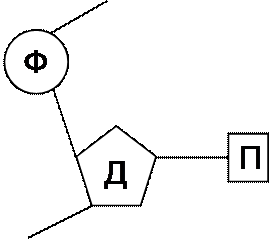


 Такой механизм репликации помогает понять, почему количество ДНК (единственного химического компонента клетки) строго постоянно во всех клетках организма, а хромосомы, на долю которых приходится большая часть содержащейся в клетке ДНК, представляют собой единственные органоиды, репродукция которых отличается большой точностью, как в качественном, так и количественном отношении. Такой способ рас-
Такой механизм репликации помогает понять, почему количество ДНК (единственного химического компонента клетки) строго постоянно во всех клетках организма, а хромосомы, на долю которых приходится большая часть содержащейся в клетке ДНК, представляют собой единственные органоиды, репродукция которых отличается большой точностью, как в качественном, так и количественном отношении. Такой способ рас-


