|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Нервные сплетенияСуществуют шейное, плечевое, поясничное и крестцовое сплетения, образованные ветвями спинномозговых нервов. Шейное сплетение образовано передними ветвями 4 верхних шейных нервов, лежит на глубоких мышцах шеи, ветви делятся на двигательные, смешанные и чувствительные. Двигательные ветви иннервируют глубокие мышцы шеи, мышцы шеи, расположенные ниже подъязычной кости, трапецевидные и грудино-ключично-сосцевидные мышцы. Смешанной ветвью является диафрагмальный нерв. Двигательные волокна его иннервируют диафрагму, чувствительные – плевру и перикард. Чувствительные ветви иннервируют кожу затылка, уха, шеи, кожу под ключицей и над дельтовидной мышцей. Плечевое сплетение образовано передними ветвями 4 нижних шейных нервов и передней ветвью первого грудного нерва. Иннервирует мышцы груди, плечевого пояса и спины. Подключичный отдел плечевого сплетения образует 3 пучка – медиальный, латеральный и задний. Нервы, выходящие из этих пучков, иннервируют мышцы и кожу верхней конечности. Передние ветви грудных нервов (1-11) сплетений не образуют, идут как межреберные нервы. Чувствительные волокна иннервируют кожу груди и живота, двигательные – межреберные мышцы, некоторые мышцы груди и живота. Поясничное сплетение образовано передними ветвями 12 грудного, 1-4 ветвями поясничных нервов. Ветви поясничного сплетения иннервируют мышцы живота, поясницы, мышцы передней поверхности бедра, мышцы медиальной группы бедра. Чувствительные волокна иннервируют кожу ниже паховой связки, промежности, кожу бедра. Крестцовое сплетение образовано ветвями 4 и 5 поясничных нервов. Двигательные ветви иннервируют мышцы промежности, ягодицы, промежности; чувствительные – кожу промежности и наружных половых органов. Длинные ветви крестцового сплетения образуют седалищный нерв – самый крупный нерв тела, иннервирующий мышцы нижней конечности.
3. Классификация нервных волокон. По функциональным свойствам (строению, диаметру волокна, электровозбудимости, скорости развития потенциала действия, длительности различных фаз потенциала действия, по скорости проведения возбуждения) Эрлангер и Гассер разделили нервные волокна на волокна групп А, В и С. Группа А неоднородна, волокна типа А в свою очередь делятся на подтипы: А-альфа, А-бета, А-гамма, А-дельта. Волокна типа А покрыты миелиновой оболочкой. Наиболее толстые из них А-альфа имеют диаметр 12-22мкм и высокую скорость проведения возбуждения — 70-120 м/с. Эти волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам (двигательные волокна) и от проприорецепторов мышц к соответствующим нервным центрам. Три другие группы волокон типа А (бета, гамма, дельта) имеют меньший диаметр от 8 до 1 мкм и меньшую скорость проведения возбуждения от 5 до 70 м/с. Волокна этих групп относятся преимущественно к чувствительным, проводящим возбуждение от различных рецепторов (тактильных, температурных, некоторых болевых рецепторов внутренних органов) в ЦНС. Исключение составляют лишь гамма-волокна, значительная часть которых проводит возбуждение от клеток спинного мозга к интрафузальным мышечным волокнам. К волокнам типа В относятся миелинизированные преганглионарные волокна вегетативной нервной системы. Их диаметр — 1- мкм, а скорость проведения возбуждения — 3-18 м/с. К волокнам типа С относятся безмиелиновые нервные волокна малого диаметра — 0,5-2,0 мкм. Скорость проведения возбуждения в этих волокнах не более 3 м/с (0,5-3,0 м/с). Большинство волокон типа С — это постганглионарные волокна симпатического отдела вегетативной нервной системы, а также нервные волокна, которые проводят возбуждение от болевых рецепторов, некоторых терморецепторов и рецепторов давления.
4. Законы проведения возбуждения по нервам. Нервное волокно обладает следующими физиологическими свойствами: возбудимостью, проводимостью, лабильностью. Проведение возбуждения по нервным волокнам осуществляется по определенным законам. Закон двустороннего проведения возбуждения по нервному волокну. Нервы обладают двусторонней проводимостью, т.е. возбуждение может распространяться в любом направлении от возбужденного участка (места его возникновения), т. е., центростремительно и центробежно. Это можно доказать, если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют электроды по обе стороны от места раздражения. Естественным направлением распространения возбуждения является: в афферентных проводниках - от рецептора к клетке, в эфферентных - от клетки к рабочему органу. Закон анатомической и физиологической целостности нервного волокна. Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность, т.е. передача возбуждения возможна только по структурно и функционально не измененному, неповрежденному нерву (законы анатомической и физиологической целостности). Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т. д.) приводят к нарушению физиологической целостности, т. е., к нарушению механизмов передачи возбуждения. Несмотря на сохранение его анатомической целостности проведение возбуждения в таких условиях нарушается. Закон изолированного проведения возбуждения по нервному волокну. В составе нерва возбуждение по нервному волокну распространяется изолированно, без перехода на другие волокна, имеющиеся в составе нерва. Изолированное проведение возбуждения обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон. Поэтому основная часть тока, возникающего между возбужденным и невозбужденным участками нервного волокна, проходит по межклеточным щелям, не действуя на рядом расположенные нервные волокна. Изолированное проведение возбуждения имеет важное значение. Нерв содержит большое количество нервных волокон (чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки; ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервноговолокна на другое, то нормальное функционирование органов было бы невозможно. Возбуждение (потенциал действия) распространяется по нервному волокну без затухания. Периферический нерв практически неутомляем. Механизм проведения возбуждения по нерву. Возбуждение (потенциал действия - ПД) распространяется в аксонах, телах нервных клеток, а также иногда в дендритах без снижения амплитуды и без снижения скорости (бездекрементно). Механизм распространения возбуждения у различных нервных волокон неодинаков. При распространении возбуждения по безмиелиновому нервному волокну механизм проведения включает два компонента: раздражающее действие катэлектротона, порождаемое локальным ПД, на соседний участок электровозбудимой мембраны и возникновение ПД в этом участке мембраны. Локальная деполяризация мембраны нарушает электрическую стабильность мембраны, различная величина поляризации мембраны в смежных ее участках порождает электродвижущую силу и местный электрический ток, силовые линии которого замыкаются через ионные каналы. Активация ионного канала повышает натриевую проводимость, после электротонического достижения критического уровня деполяризации (КУД) в новом участке мембраны генерируется ПД. В свою очередь этот потенциал действия вызывает местные токи, а они в новом участке мембраны генерируют потенциал действия. На всем протяжении нервного волокна происходит процесс новой генерации потенциала действия мембраны волокна. Данный тип передачи возбуждения называется непрерывным. Скорость распространения возбуждения пропорциональна толщине волокна и обратно пропорциональна сопротивлению среды. Проведение возбуждения зависит от соотношения амплитуды ПД и величины порогового потенциала. Этот показатель называется гарантийный фактор (ГФ) и равен 5 - 7, т.е. ПД должен быть выше порогового потенциала в 5- 7 раз. Если ГФ = 1 проведение ненадёжно, если ГФ < 1 проведения нет. Протяженность возбуждённого участка нерва L является произведение времени (длительности) ПД и скорости распространения ПД. Например, в гигантском аксоне кальмара L= 1 мс ´ 25 мм/мс = 25 мм. Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки - перехватов Ранвье создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. В миелинизированном волокне токи проводятся только в зонах, не покрытых миелином, - перехватах Ранвье, в этих участках и генерируется очередной ПД. Перехваты длиной 1 мкм расположены через 1000 - 2000 мкм, характеризуются высокой плотностью ионных каналов, высокой электропроводностью и низким сопротивлением. Распространение ПД в миэлинизированных нервных волокнах осуществляется сальтаторно - скачкообразно от перехвата к перехвату, т.е. возбуждение (ПД) как бы «перепрыгивает» через участки нервного волокна, покрытые миелином, от одного перехвата к другому. Скорость такого способа проведения возбуждения значительно выше, и он более экономичен по сравнению с непрерывным проведением возбуждения, поскольку в состояние активности вовлекается не вся мембрана, а только ее небольшие участки в области перехватов, благодаря чему уменьшается нагрузка на ионный насос.
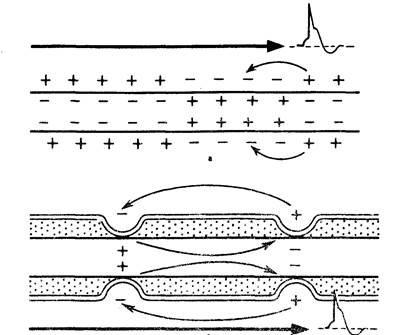
Схема распространения возбуждения в безмиелиновых и миелиновых нервных волокнах. 5. Парабиоз. Нервные волокна обладают лабильностью — способностью воспроизводить определенное количество циклов возбуждения в единицу времени в соответствии с ритмом действующих раздражителей. Мерой лабильности является максимальное количество циклов возбуждения, которое способно воспроизвести нервное волокно в единицу времени без трансформации ритма раздражения. Лабильность определяется длительностью пика потенциала действия, т. е. фазой абсолютной рефрактерности. Так как длительность абсолютной рефрактерности у спайкового потенциала нервного волокна самая короткая, то лабильность его самая высокая. Нервное волокно способно воспроизвести до 1000 импульсов в секунду. Явление парабиоза открыто русским физиологом Н.Е.Введенским в 1901 г. при изучении возбудимости нервно-мышечного препарата. Состояние парабиоза могут вызвать различные воздействия – сверхчастые, сверхсильные стимулы, яды, лекарства и другие воздействия как в норме, так и при патологии. Н. Е. Введенский обнаружил, что если участок нерва подвергнуть альтерации (т. е. воздействию повреждающего агента), то лабильность такого участка резко снижается. Восстановление исходного состояния нервного волокна после каждого потенциала действия в поврежденном участке происходит медленно. При действии на этот участок частых раздражителей он не в состоянии воспроизвести заданный ритм раздражения, и поэтому проведение импульсов блокируется. Такое состояние пониженной лабильности и было названо Н. Е. Введенским парабиозом.Состояние парабиоза возбудимой ткани возникает под влиянием сильных раздражителей и характеризуется фазными нарушениями проводимости и возбудимости. Выделяют 3 фазы: первичную, фазу наибольшей активности (оптимум) и фазу сниженной активности (пессимум). Третья фаза объединяет 3 последовательно сменяющие друг друга стадии: уравнительную (провизорная, трансформирующая – по Н.Е.Введенскому), парадоксальную и тормозную. Первая фаза (примум) характеризуется снижением возбудимости и повышением лабильности. Во вторую фазу (оптимум) возбудимость достигает максимума, лабильность начинает снижаться. В третью фазу (пессимум) возбудимость и лабильность снижаются параллельно и развивается 3 стадии парабиоза. Первая стадия - уравнительная по И.П.Павлову - характеризуется выравниванием ответов на сильные, частые и умеренные раздражения. В уравнительную фазу происходит уравнивание величины ответной реакции на частые и редкие раздражители. В нормальных условиях функционирования нервного волокна величина ответной реакции иннервируемых им мышечных волокон подчиняется закону силы: на редкие раздражители ответная реакция меньше, а на частые раздражители—больше. При действии парабиотического агента и при редком ритме раздражении (например, 25 Гц) все импульсы возбуждения проводятся через парабиотический участок, так как возбудимость после предыдущего импульса успевает восстановиться. При высоком ритме раздражении (100 Гц) последующие импульсы могут поступать в тот момент, когда нервное волокно еще находится в состоянии относительной рефрактерности, вызванной предыдущим потенциалом действия. Поэтому часть импульсов не проводится. Если проводится только каждое четвертое возбуждение (т.е. 25 импульсов из 100), то амплитуда ответной реакции становится такой же, как на редкие раздражители (25 Гц)—происходит уравнивание ответной реакции. Вторая стадия характеризуется извращенным реагированием – сильные раздражения вызывают меньший ответ, чем умеренные. В эту - парадоксальную фазу происходит дальнейшее снижение лабильности. При этом на редкие и частые раздражители ответная реакция возникает, но на частые раздражители она значительно меньше, т. к. частые раздражители еще больше снижают лабильность, удлиняя фазу абсолютной рефрактерности. Следовательно, наблюдается парадокс — на редкие раздражители ответная реакция больше, чем на частые. В тормозную фазу лабильность снижается до такой степени, что и редкие, и частые раздражители не вызывают ответной реакции. При этом мембрана нервного волокна деполяризована и не переходит в стадию реполяризации, т. е. не восстанавливается ее исходное состояние. Ни сильные, ни умеренные раздражения не вызывают видимой реакции, в ткани развивается торможение. Парабиоз — явление обратимое. Если парабиотическое вещество действует недолго, то после прекращения его действия нерв выходит из состояния парабиоза через те же фазы, но в обратной последовательности. Однако, при действии сильных раздражителей за тормозной стадией может наступить полная потеря возбудимости и проводимости, а в дальнейшем – гибель ткани. Работы Н.Е.Введенского по парабиозу сыграли важную роль в развитии нейрофизиологии и клинической медицины, показав единство процессов возбуждения, торможения и покоя, изменили господствовавший в физиологии закон силовых отношений, согласно которому реакция тем больше, чем сильнее действующий раздражитель. Явление парабиоза лежит в основе медикаментозного локального обезболивания. Влияние анестезирующих веществ вязано с понижением лабильности и нарушением механизма проведения возбуждения по нервным волокнам. Дата добавления: 2015-12-15 | Просмотры: 1634 | Нарушение авторских прав |