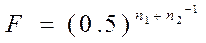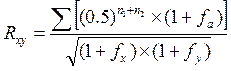|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Медико-генетическое консультирование. Для подавляющего большинства наследственных болезней эффективных способов лечения не существует
Для подавляющего большинства наследственных болезней эффективных способов лечения не существует. Из этого следует, что в борьбе с наследственной патологией основная роль отводится профилактике рождения аномального потомства. Общий профилактический характер носят мероприятия, направленные на оздоровление окружающей среды, способствующие снижению ее мутагенного воздействия на наследственный материал человеческого организма. В последние десятилетия распространенным и эффективным способом профилактики наследственных болезней является медико-генетическое консультирование. Медико-генетическое консультирование — это один из видов специализированной помощи населению, направленной в первую очередь на предупреждение появления в семье детей с наследственной патологией. С этой целью составляют прогноз рождения в данной семье ребенка с наследственной болезнью, родителям объясняют вероятность этого события и оказывают помощь в принятии решения. В случае большой вероятности рождения больного ребенка родителям рекомендуют либо воздержаться от деторождения, либо провести пренатальную диагностику, если она возможна при данном виде патологии. Консультирование семей, обращающихся к врачу-генетику, включает три основных этапа. Как правило, за консультацией обращаются семьи, где уже имеется ребенок с наследственной патологией, или семьи, в которых имеются больные родственники. На первом этапе консультирования производится уточнение диагноза, что является необходимой предпосылкой любого консультирования. Уточнение диагноза в медико-генетической консультации проводят с помощью генетического анализа. Для этой цели используют генеалогический, цитогенетический, биохимический и другие требуемые методы исследований, которым подвергаются пробанд и его родственники. Точный клинический и генетический диагноз заболевания позволяет установить степень генетического риска и выбор эффективных методов пренатальной диагностики и профилактического лечения. На втором этапе консультирования делают прогноз потомства. Генетический риск может быть определен либо путем теоретических расчетов, основанных на генетических закономерностях, либо с помощью эмпирических данных. Сущность генетического прогноза заключается в определении вероятности появления наследственной патологии в семье. Наиболее эффективным является проспективное консультирование, когда риск рождения больного ребенка определяют до наступления беременности или в ранние ее сроки. Такие консультации чаще проводят в случае кровного родства супругов, при отягощенной наследственности по линии мужа или жены, при воздействии вредных средовых факторов на супругов незадолго до наступления беременности. Ретроспективное консультирование проводят после рождения больного ребенка относительно здоровья будущих детей. Определение прогноза потомства при разных формах наследственной патологии различно. При моногенных, менделирующих болезнях прогноз основывается на расчете вероятности появления потомства в соответствии с генетическими закономерностями. При этом, если известен тип наследования данного заболевания и по родословной удается установить генотип родителей, оценка риска сводится к анализу мевделевского расщепления. Если у пробанда установлена вновь возникшая мутация, то повторный риск рождения ребенка с такой же патологией незначителен. Расчет риска при моногенном заболевании может осложниться при пониженной экспрессивности или неполной пенетрантности гена, позднем проявлении генетической аномалии, генетической гетерогенности заболевания и вообще в случае неточного диагноза. При хромосомных болезнях определение риска повторного рождения потомства с хромосомными аномалиями зависит от того, нормальны ли кариотипы родителей, не обнаружено ли у них мозаицизма, не наблюдается ли семейной формы структурных аномалий хромосом. В случае отсутствия нарушений в кариотипе родителей вероятность повторного рождения второго ребенка с хромосомной аномалией оценивается по эмпирическим данным для каждого вида аномалии с учетом возраста родителей. При мультифакториальных заболеваниях, т.е. заболеваниях с наследственным предрасположением, основой оценки риска являются эмпирические данные о популяционной и семейной частоте каждого из них. Специфический генетический риск до 5% принято считать низким, до 10% —повышенным в легкой степени, до 20% —средним, выше 20% — высоким. Генетический риск средней степени расценивают как противопоказание к зачатию или показание к прерыванию уже имеющейся беременности. Возможность проведения пренатальной диагностики является определяющей для принятия положительного решения в отношении завершения беременности. На третьем этапе консультирования врач-генетик в доступной форме объясняет семье степень генетического риска рождения наследственно аномального потомства, сущность пренатальной диагностики и помогает принять правильное решение в отношении деторождения. Однако окончательное решение этого вопроса остается за родителями. 30. Инбридинг (случайный, неслучайный, тотальный), его роль как фактор изменения генофонда популяции.
Спаривание родственных между собой самцов и самок называется инбридингом. Потомство, полученное в результате родственного подбора, называют инбредным. Спаривание неродственных животных называют аутбридингом. Родство между животными означает, что они имеют одного или нескольких общих предков. В результате этого инбредные животные имеют между собой и с родителями определенное генетическое сходство, в частности по составу аллелей в их генотипе. Поэтому в результате спаривания родственных, а следовательно, и сходных между собой животных у их потомства происходит накопление аллелей и генотипов того предка, который является общим для спариваемых родственных особей. Инбридинг приводит к повышению частоты гомозиготных генотипов у потомков и снижению частоты гетерозиготных генотипов при сохранении частот аллелей. Родственное спаривание сопровождается снижением генетической изменчивости. Наиболее близкородственной формой размножения в животном мире является самооплодотворение, встречающееся редко у низших филогенетических форм. У растений такой процесс прослеживается при самоопылении (например, у гороха, ячменя и др.). В практике животноводства применяют разные типы инбридинга, когда спаривают близких между собой родственников (отца с дочерью, мать с сыном, брата с сестрой); это называется кровосмешением. Другая группа подбора при инбридинге — это разведение в близком родстве, когда спариваются менее близкие родственники: полубрат — полусестра, бабушка — внук, внучка — дед, и еще более отдаленные родственники, то есть разведение в умеренном и отдаленном родстве. В практике животноводства инбридинг применяют как при чистопородном разведении, так и при межпородном скрещивании. При этом степень родства между спариваемыми животными разнообразна. В современной селекции приняты следующие схемы и терминология инбридинга: · родственное разведение - инбридинг; · использование родственного разведения в нескольких поколениях - ин-энд-инбридинг; · кровосмешение (близкородственное разведение) — клозебридинг; · спаривание животных из разных инбредных линий одной породы — инбредлайнкроссинг; · спаривание животных из близкородственных линий — стра-инкроссинг; · спаривание инбредных самцов с неинбредными самками — топкроссинг, · скрещивание инбредных самцов с неинбредными самками другой породы — топкроссбридинг; · скрещивание инбредных самцов одной породы с инбредными самками другой породы — инкроссбридинг; · спаривание инбредных маток с аутбредными самцами — боттомкроссинг. Наиболее распространен в практике учет инбридинга, предложенный Шапоружем (1909). По этому методу учитывается число рядов поколений"; отделяющих потомка от предка, на которого осуществлен инбридинг. Начиная от ряда родительского поколения, обозначаемого римской цифрой I, далее каждый последующий ряд записывается как II, III, IV и т. д. Повторяющийся предок в родословной потомка со стороны матери и отца записывается с указанием ряда поколений, в которых он присутствует по материнской и отцовской стороне родословной. Более точный метод определения степени инбридинга был разработан С. Райтом (1921) на основании принципа путевого анализа. Для этого была предложена формула коэффициента инбридинга. Позднее профессором Д. А. Кисловским в формулу Райта было внесено уточнение, что делает вычисление более удобным. Формула Райта — Кисловского представляет коэффициент инбридинга F в виде дроби в границах от 0 до 1 (или в %).
Чем больше величина F приближается к единице (или 100%), тем сильнее инбридирован потомок на предка, тем больше у потомков можно ожидать проявления инбредной депрессии и тем более вероятность повышения гомозиготности потомка по генам предка. Коэффициент инбридинга не указывает в абсолютных цифрах или в процентах, насколько гомозиготен потомок; он только свидетельствует о вероятности того, насколько примененный родственный подбор увеличит гомози-готность потомка по сравнению с исходным состоянием генотипа. У животных с высокой степенью гетерозиготности инбридинг усиливает гомозиготность значительно быстрее, чем у Животных, уже имевших значительную гомозиготность до инбридинга. Уровень гомозиготности потомка зависит от того, были ли его родители гетерозиготны или гомозиготны. Использование гетерозиготных производителей — наиболее перспективный вариант инбридинга при проведении племенной работы в стаде, так как он не сопровождается депрессией у потомка, но обеспечивает более стабильное сходство с ценным предком. Формула, предложенная Райтом для вычисления степени возрастания генетического сходства между двумя сравниваемыми животными, такова:
где: Rxy – коэффициент генетического сходства между животными x и y; n1 и n2 число поколений от данных животных до общего предка по женской и мужской стороне родословной; fx и fyкоэффициент возрастания гомозиготности для животных x и y; fa – тот же коэффициент для общего инбридированного предка. Так, в XVIII в. в Англии братья Коллинги в результате селекции получили быка Комета шортгорнской породы, который был инбридирован на четырех выдающихся предков: быков Фаворита и Фольджамба и коров Феникс и Леди Майнард, при этом корова Леди Майнард и бык Фольджамб были инбредными животными. Коэффициент инбридинга был достаточно высоким (F=46,87%), но, несмотря на это, Комет сыграл важную роль в совершенствовании породы. Депрессия может наблюдаться и при так называемом ложном инбридинге, который является следствием содержания животных в ряде поколений в одинаковых, часто изнеживающих условиях. В целях устранения депрессии при истинном и ложном инбридинге целесообразно периодически содержать животных, намеченных к спариванию, в различных условиях, что способствует формированию некоторого биологического несходства гамет самцов и самок. Этот прием называют интербридингом. Использование инбридинга в селекции растений. Раздельнополость и приспособленность к перекрестному опылению, столь распространенные в растительном мире, и существование у растений многочисленных препятствий к самоопылению выработаны в процессе эволюции. Они не могли бы так широко распространиться, если перекрестное опыление не имело бы в борьбе за жизнь каких-то вполне определенных преимуществ перед самоопылением и не было бы, поэтому подхвачено естественным отбором. Естественным отбором создана и приспособленность растений определенных видов к соответствующим насекомым — их опылителям и, наоборот, насекомых определенных видов к соответствующим растениям. У ветроопыляемых же растений эволюционно развилась необходимость вырабатывать огромный избыток пыльцы; только тогда при случайных порывах ветра на рыльца пестиков попадет какая-то ее часть. К такому выводу Ч. Дарвин пришел на основании изучения природы. Для подтверждения его Дарвин поставил специальные опыты, продолжавшиеся более 10 лет на растениях 57 различных видов. Растения каждого вида он разделил на две разные части и представителей одной из них размножал в ряде последующих поколений путем самоопыления, а другой — путем перекрестного опыления. Дочерние растения выращивали в возможно более сходных условиях — в одних горшках или на одних грядках. У полученных от перекрестного опыления растений показатели высоты, веса и плодовитости оказались в среднем значительно больше. Так, высота растений, происшедших от самоопыления, по сравнению с полученными от перекрестного опыления составляла в среднем только 87%, а вес — лишь 67%.
31. Естественный отбор, определение его величины в человеческих популяция.
В природных популяциях организмов, размножающихся половым способом, существует большое разнообразие генотипов и, следовательно, фенотипов. Благодаря индивидуальной изменчивости в условиях конкретной среды обитания приспособленность разных генотипов (фенотипов) различна. В эволюционном контексте приспособленность определяют как произведение жизнеспособности в данной среде, обусловливающей большую или меньшую вероятность достижения репродуктивного возраста, на репродуктивную способность особи. Различия между организмами по приспособленности, оцениваемой передачей аллелей следующему поколению, выявляются в природе с помощью естественного отбора. Главный результат отбора заключается не просто в выживании более жизнеспособных, а в относительном вкладе таких особей в генофонд дочерней популяции. Необходимой предпосылкой отбора является борьба за существование — конкуренция за пищу, жизненное пространство, партнера для спаривания. Естественный отбор происходит на всех стадиях онтогенеза организмов. На дорепродуктивных стадиях индивидуального развития, например в эмбриогенезе, преобладающим механизмом отбора служит дифференциальная (избирательная) смертность. В конечном итоге отбор обеспечивает дифференциальное (избирательное) воспроизведение (размножение) генотипов. Благодаря естественному отбору аллели (признаки), повышающие выживаемость и репродуктивную способность, накапливаются в ряду поколений, изменяя генетический состав популяций в биологически целесообразном направлении. В природных условиях естественный отбор осуществляется исключительно по фенотипу. Отбор генотипов происходит вторично через отбор фенотипов, которые отражают генетическую конституцию организмов. Как элементарный эволюционный фактор естественный отбор действует в популяциях. Популяция является полем действия, отдельные особи — объектами действия, а конкретные признаки — точками приложения отбора. Эффективность отбора по качественному и количественному изменению генофонда популяции зависит от величины давления и направления его действия. Величину давления отбора выражают коэффициентом отбора S, который характеризует интенсивность устранения из репродуктивного процесса или сохранения в нем соответственно менее или более приспособленных форм по сравнению с формой, принятой за стандарт приспособленности. Так, если некий локус представлен аллелями A1 и А2, то популяция по генотипам делится на три группы: A1A1; A1A2; A2A2. Обозначим приспособленность этих генотипов W0, W1, W2. Выберем в качестве стандарта первый генотип, относительная приспособленность которого максимальна и равна 1. Тогда приспособленности других генотипов составят доли этого стандарта:
AlA1 A1A2 A2A2 W0 W1 W2 или W0/W0 = 1, W1/W0=l—S1, W2/W0=l—S2.
Величины S1 и S2 означают пропорциональное снижение в очередном поколении воспроизводства генотипов A1A2 и А2А2 сравнительно с генотипом AlA1. Отбор особенно эффективен в отношении доминантных аллелей при условии их полного фенотипического проявления и менее эффективен в отношении рецессивных аллелей, а также в условиях неполной пенетрантности. На результат отбора влияет исходная концентрация аллеля в генофонде. При низких и высоких концентрациях отбор происходит медленно. Изменение доли доминантного аллеля в сравнении с рецессивным при коэффициенте отбора 0,01 приведено ниже.
В теории, упрощая ситуацию, допускают, что отбор через фенотипы действует на генотипы благодаря различиям в приспособительной ценности отдельных аллелей. В реальной жизни приспособительная ценность генотипов зависит от влияния на фенотип и взаимодействия всей совокупности генов. Оценка величины давления отбора по изменению концентрации отдельных аллелей технически часто невыполнима. Поэтому расчет проводят по изменению концентрации организмов определенного фенотипа. Пусть в популяции присутствуют организмы двух фенотипических классов А и В в отношении СA/СB =U1. Вследствие различий по приспособленности происходит естественный отбор (селекция), который изменяет соотношение особей с фенотипами А и В. В следующем поколении оно станет СA/СB =U2 =U1 (1 +S), где S — коэффициент отбора. Отсюда S = U2/U1 — 1. При селективном преимуществе фенотипа А U2>U1, aS> 0. При селективном преимуществе фенотипа В U2< U1 и S < 0. Если приспособленность фенотипов А и В сопоставима и U2 =U1, aS=0. В рассмотренном примере при S > 0 отбор сохраняет в популяции в ряду поколений фенотипы А и устраняет фенотипы В, при S < 0 имеет место обратная тенденция. Отбор, сохраняющий определенные фенотипы, по своему направлению является положительным, тогда как отбор, устраняющий фенотипы из популяции,— отрицательным. В зависимости от результата различают стабилизирующую, движущую и дизруптивную формы естественного отбора (рис. 11.4). Стабилизирующий отбор сохраняет в популяции средний вариант фенотипа или признака. Он устраняет из репродуктивного процесса фенотипы, уклоняющиеся от сложившейся адаптивной «нормы», приводит к преимущественному размножению типичных организмов. Так, сотрудник одного из университетов США подобрал после снегопада и сильного ветра 136 оглушенных воробьев Passerdomesticus. Из них 72 выживших воробья имели крылья средней длины, тогда как 64 погибшие птицы были либо длиннокрылыми, либо короткокрылыми. Стабилизирующая форма соответствует консервативной роли естественного отбора. При относительном постоянстве условий среды благодаря этой форме сохраняются результаты предшествующих этапов эволюции. Движущий (направленный) отбор обусловливает последовательное изменение фенотипа в определенном направлении, что проявляется в сдвиге средних значений отбираемых признаков в сторону их усиления или ослабления. При смене условий обитания благодаря этой форме отбора в популяции закрепляется фенотип, более соответствующий среде. После того как новое значение признака придет в оптимальное соответствие условиям среды, движущая форма отбора сменяется стабилизирующей. Примером такого отбора является замещение в популяции гавани Плимут (Англия) крабов Carcinusmaenas с широким головогрудным щитком животными с узким щитком в связи с увеличением количества ила.
Направленный отбор составляет основу искусственного отбора. Так, в одном эксперименте на протяжении ряда поколений из популяции шестинедельных мышей отбирали для скрещивания наиболее тяжелых и наиболее легких животных. Избирательное воспроизведение по признаку массы тела привело к образованию двух самостоятельных популяций, соответственно с возрастающей и убывающей массой тела (рис. 11.5). По окончании опыта, занявшего 11 поколений, ни одна из этих популяций не вернулась к первоначальной массе. Дизруптивный (разрывающий) отбор сохраняет несколько разных фенотипов с равной приспособленностью. Он действует против особей со средним или промежуточным значением признаков. Так, в зависимости от преобладающего цвета почвы улитки Cepaeanemoralis имеют раковины коричневой, желтой, розовой окраски. Дизруптивная форма отбора «разрывает» популяцию по определенному признаку на несколько групп. Она поддерживает в популяции состояние генетического полиморфизма. В зависимости от формы отбор сокращает масштабы изменчивости, создает новую или сохраняет прежнюю картину разнообразия. Как и другие элементарные эволюционные факторы, естественный отбор вызывает изменения соотношений аллелей в генофондах популяций. Особенность его действия состоит в том, что эти изменения направленны. Отбор приводит генофонды в соответствие с критерием приспособленности. Он осуществляет обратную связь между изменениями генофонда и условиями обитания, накладывает на эти изменения печать биологической целесообразности (полезности). Естественный отбор действует совместно с другими эволюционными факторами. Поддерживая генотипическое разнообразие особей в ряду поколений, мутационный процесс, а также популяционные волны, комбинативная изменчивость создают для него необходимый материал. Естественный отбор нельзя рассматривать как «сито», сортирующее генотипы по приспособленности. В эволюции ему принадлежит творческая роль. Исключая из размножения генотипы с малой приспособительной ценностью, сохраняя благоприятные генные комбинации разного масштаба, он преобразует картину генотипической изменчивости, складывающуюся первоначально под действием случайных факторов, в биологически целесообразном направлении. Результатом творческой роли отбора является процесс органической эволюции, идущей в целом по линии прогрессивного усложнения морфофизиологической организации (арогенез), а в отдельных ветвях — пути специализации (аллогенез). 32. Хромосомный мозаицизм, его формирование, фенотипическое проявление у человека. Фенокопии, их сущность. Мозаицизм (генетический мозаицизм, хромосомный мозаицизм — mosaicism; мозаичность; могут употребляться синонимы «мозаичная форма», «мозаичный кариотип») — от фр. mosaique «мозаика» — наличие в тканях (растения, животного, человека) генетически различающихся клеток. Следует отличать мозаицизм от химеризма, при котором два (или более) генотипа происходят более чем от одной зиготы. Понятие мозаицизма связано с понятиями трисомии и анеуплоидии. Дата добавления: 2015-12-16 | Просмотры: 747 | Нарушение авторских прав |