|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
АВС – 71, АВс – 3, АbС – 14, Аbс – 17,I II Р ♀ ZW х ♂ ZZ Р ♀ ZO х ♂ ZZ Гаметалары ♀ Z; W ♂ Z ♀ Z; O ♂ Z F1 ♀ 1ZW: ♂ 1ZZ F1 ♀ 1ZO: ♂ 1ZZ Жынысты анықтаудың - гаплодиплоидея деп аталатын тағы бiр типi белгiлi. Мұны жынысты анықтаудың төртiншi типiне жатқызуға болады. Ол араларда және құмырсқаларда кеңiнен тараған. Бұл организмдерде жыныс хромосомалары болмайды. Мысалы, бал аралардың аналықтары диплоидты, ал аталықтары (трутендер) гаплоидты болады. Ұрықтанған жұмыртқалардан аналық дарабастар, ал ұрықтанбаған жұмыртқалардан партеногенез жолымен трутендер өсiп дамиды. Сперматогенез процесi кезiнде трутендердiң хромосомаларының саны редукцияға ұшырамайды. Хромосомалардың гаплоидты саны трутендердiң соматикалық клеткаларында сақталып, олар жыныс клеткаларын бередi. Ұрпақ беруге қабiлетсiз клеткалар өздерiнiң жеке дамуы кезiнде хромосомалардың диплоидты санын қайтадан қалпына келтiредi. 39. Жыныс хромосомалары ажырамаған кезіндегі белгілердің тұқым қуалауын түсіндіріңіз. Адамдардағы жыныс хромосомасының ажырамауы және хромосомалардың дұрыс ажырамауынан туындайтын ауруларды атаңыз және себебін түсіндіріңіз. Будандастырылуларда белгiлердiң берiлу заңдылықтарының мейоз бен ұрықтану кезiндегi хромосомалардың сипатына сәйкес келуi - гендер хромосомаларда орналасқан деген қорытындыға әкелдi. Жыныспен тiркескен тұқым қуалауды да осы негiзде жақсы түсiнуге болатындығы, бұл қорытындынығң нанымдылығын арттыра түседі.Жоғарыда айтылғандай, дрозофиланың ақ көздi аналықтарын қызыл көздi аталығымен шағылыстырса, F1 -дегi аналық ұрпақтардың барлығы қызыл көздi, ал анасынан алған жалғыз Х-хромосомасы бар аталықтардың барлығы ақ көздi болады. Бiрақ осындай шағылысулардан алынған екi-үш мың ұрпақтар iшiнде бiрен-саран «ерекше» дарабастар ақ көздi аналық немесе қызыл көздi аталық кездеседi. Бұл «ерекше» дарабастардың пайда болуын былайша түсiндiруге болады. Мейоз кезiнде «ерекше» дарабастардың анасының екi Х-хромосомасы әр полюске ажыраудың орнына, кездейсоқ не бiр жұмыртқаға түседi немесе екеуi де полярлы денешiкке кетедi, яғни Х-хромосомалардың гаметаларға ажырамау процесi өтедi. Нәтижесiнде ақ көздi аналықта әдеттегiден өзгеше жұмыртқаның екi түрi: ХХ-хромосомасы бар және ешбiр Х-хромосомасы жоқ жұмыртқалар пайда болады. Осындай жұмыртқалардың әрқайсысы не Х-хромосомалы спермиймен немесе У-хромосомалы спермиймен ұрықтана алады, бұдан зиготалардың төрт типi пайда болуы мүмкiн: 1) ХХХ-хромосомалы; 2) ХХУ-хромосомалы; 3) Х-хромосомалы; 4) жалғыз У-хромосомалы. «Ерекше» дарабастарды цитологиялық әдiстермен зерттегенде, F1 -дiң «ерекше» ақ көздi аналықтарында екi Х-хромосомаларына қосымша У-хромосомасы бар болып шықты, ал «ерекше» қызыл көздi аталықтарында бiр Х-хромосома бар, бiрақ аталықтарына тән У-хромосома жоқ болып шықты. Жыныс хромосомаларының бiрiншi реттiк ажырамауы нәтижесiнде пайда болған «ерекше» аталықтардың барлық сыртқы белгiлерi аталыққа тән, бiрақ олар ұрпақ бермейтiн бедеулер болады. Ал бұл дрозофиланың У-хромосомасында жынысты анықтайтын гендердiң жоқ екендiгiн, тек қана аталықтардың өсiмталдығын қамтамасыз ететiн гендердiң болатындығын көрсетедi. «Ерекше» аналықтар ХХY, керiсiнше, өсiмтал, бiрақ жыныс хромосомаларының мейоздық ажырауы кәдiмгi аналықтардан ХХ өзгеше етедi. Мұндай «ерекше» аналықтардың Х-хромосомалары бiр-бiрiмен жиiрек коньюгацияланады және әр полюстарға кетедi. Ал У-хромосома коньюгацияға қатыспайды және жас ядролар арасында кездейсоқ таралады, нәтижесiнде жұмыртқалардың бiрдей мөлшерде екi сорты пайда болады: жартысының Х-хромосомасы, ал жартысының Х-хромосомасы және У-хромосомасы болады. Мұнымен қоса шамамен 8 жағдайда Х-хромосомалардың бiреуiмен У-хромосома арасында коньюгация жүредi және онда Х-хромосомалар екiншi рет кездейсоқ таралады, нәтижесiнде жұмыртқалардың тағы да екi сорты пайда болады: ХХ-хромосомасы бар жұмыртқалар және Х-хромосомасы жоқ, бiрақ У-хромосомасы бар жұмыртқалар.Бұл құбылыс жыныс хромосомаларының екiншi реттiк ажырамауы деп аталады. Бiрсыпыра организмдерде, оның iшiнде адамда да жыныс хромосомаларының ажырамау фактiлерi белгiлi. Мысалы, Х-хромосомалары ажырамаған әйелдерде гаметалардың екi сорты (ХХ және О) түзiледi және олар аталықтардың екi сортты гаметаларымен (Х және У) ұрықтанғанда ұрпақтардың төрт типi пайда болады. ХХХ - мұндай зиготадан аналық дамиды, Х-хромосомасы бойынша олар трисомиктер, бiрақ, әдеттегiден өзгеше, олардың арасында ақыл кемiстiгi, ұрпақсыздық жиi кездеседi. ХО - мұндай зиготалар көпшiлiк жағдайда эмбрион кезiнде өледi, қалған зиготалардан аналық дамиды, бiрақ оларға Шерешевский-Тернер синдромына ұқсас белгiлер тән: аласа бойлы, жыныс органдары нашар жетiлген, ұрпақ беруге қабiлетсiз және басқа да инфантильдiк белгiлерi болады. ХХУ - мұндай зиготадан аталық дамиды, онда Клайнфельтер синдромына тән белгiлерi болады: ақылы кемдеу, әлсiз, аталық бездерi нашар дамыған және ұрпақ беруге қабiлетсiз (бедеу). Жыныс хромосомаларының ажырамауынан пайда болған бұл жағдай, дрозофиладан өзгеше, У-хромосомада аталық жынысты анықтайтын гендер бар екендiгiн дәлелдейдi, өйткенi адамда, екi ХХ және бiр У хромосома болуына қарамастан ХХУ зиготадан Клайнфельтер синдромына тән белгiлер болғанмен, аталық жыныс дамиды. У-хромосома жоқ болса аналық жыныс пайда болады. УО - мұндай зигота эмбрион сатысында өлiмге ұшырайды. Жыныс хромосомаларының ажырамауы дрозофилаға қарағанда адамда жиiрек кездеседi. Орта есеппен әрбiр жаңа туған 600 ер балаға хромосома формуласы ХХУ бiр ер бала келедi. Х-хромосомалардың ажырамауы әйелдердiң жасына қарай өсе түседi. Сондықтан 35-37 жастан асқан аналардан туылған балалардың iшiнде Клайнфельтер синдромымен ауыратын ер балалар мен Шерешевский-Тернер синдромымен ауыратын қыз балалар саны, жас аналардан туылған балалармен салыстырғанда жиiрек кездеседi. 40. Жынысты анықтаудың баланстық теориясының механизмін түсіндіріңіз. Жыныс хроматині. Гинандроморфизмнің пайда болу себебін көрсетіңіз. 1919 жылы Бриджес дрозофиланың триплоидты өсiмтал бiрнеше аналықтарын тапты, оларды диплоидты аталықтарымен шағылыстырды. Алынған ұрпақтарға морфологиялық, цитологиялық және генетикалық талдаулар жасау дрозофилада жыныстың анықталу себептерiн түсiнуге мүмкiндiк бердi. Осындай шағылысулардан Х-хромосомалар мен аутосомалар арасындағы арақатынасы түрлiше болып келген дарабастар алынды. Алынған осындай мәлiметтерге сүйене отырып Бриджес өзiнiң ген тепе-теңдiгi туралы теориясын тұжырымдады. Ол теория бойынша организм бипотенциалды, яғни онда аналық жыныстың да, аталық жыныстың да бастамалары болады. Жыныс дарабастың дамуы кезiнде жыныс детерминаторларының - аналық және аталық гендердiң тепе-теңдiгi негiзiнде анықталады. Жыныстық хромосома деп атау дәл емес, өйткенi Х-хромосома да, аутосомалар да бiрдей дәрежеде жыныс хромосомалары болып табылады. ХХ және ХY жүйесiнiң болуының арқасында Х-хромосома жыныстың Мендельдiк анықталуына негiз болып есептеледi. Дрозофиланың Х-хромосомасында аналыққа сәйкес гендер жиынтығы болады, ал аутосомаларда аталық гендер жинақталған. Дами бастаған зиготаның жынысының қандай болуы жыныстық Х-хромосомалар санымен аутосомалар саны арасындағы тепе-теңдiкпен анықталады (7.1-кесте). Егер бұл тепе-теңдiк Х: А = 1 болса қалыпты аналық дарабастар, ал тепе-теңдiк 3Х: 2А = 1,5 болса жұмыртқа безi және басқа белгiлерi нашар жетiлген асқын аналық дарабастар дамиды. Егер тепе-теңдiк Х: 2А = 0,5 болса қалыпты аталық дарабастар, ал тепе-теңдiк Х: 3А = 0,33 болса аталық белгiлерi жетiлмеген ұрықсыз аталық дарабастар - асқын аталық дамиды. Тепе-теңдiк 2Х: 3А = 0,67 болғанда жынысы аралық сипатта дамитын интерсексуальды дарабастар (интерсекстер) дамиды. Дрозофилада У-хромосома жынысты анықтамайды. Осы мәселелердi зерттей отырып Бриджес, триплоидты аналықтардан алынатын интерсекстер жынысының ығысуына сыртқы орта жағдайларының әсер ететiндiгiн тапты. Мысалы, жоғары температурада интерсектердiң жынысы аналық жаққа, ал төмендегi температурада - аталық жаққа ығысатыны байқалды. Осыған байланысты сұрыптау нәтижесiнде аталық немесе аналық дарабастары көп интерсекстi линиялар шығаруға болады. 41. Жынысты анықтауда сыртқы ортаның рөлін көрсетіңіз. Жыныспен тіркесіп тұқым қуалайтын ауруларды атап көрсетіңіз және себебін түсіндіріңіз. Адамның тұқым қуалайтын ауруларының саны 2000-нан асады.Олардың көпшілігі психикалық ауытқулармен сипатталады.Бұл аурулар адам генетикасындағы хромосомалардың санының, құрамының һзгеруіне немесе гендердің мутациясына байланысты. Мутагендік факторлердің әсерінен жыныс клеткаларының пісіп жетілуі кезінде хромосомдар дұрыс ажырамайды, немесе құрамында өзгеріс пайда болады.Олай болса, мұндай хромосомды жұмыртқа клеткасы қрықтанғаннан соң қалпында дамымайды.А.А.Прокофьева-Бельговскаяның деректері бойынша барлық түсініктердің 5% -тінің себебі жұмыртқа клеткасындағы хромосомдық өзгерістер.Хромосомалық өзгерістердңғ (аномалиялардың) нәтижесінде туған ауруларды хромосомдық аурулар деп атайды.Олардың қатарына Клейнфельтер, Тернер Шерешевский, даун синдромдары Трисомия – X синдромдары және т.б. жатады. Аутосомдық хромосомдардың саны өзгерціне байланысты тағы басқа синдромдар кездеседі.Эдварс синдромы – 18 жұп хромосом бойынша трисомия (үш хромосом), Патау синдромы – 15 жұп хромосомдар бойынша трисомия.Ұрпақтарында бұл синдромдардың болуы да ата-аналарында гаметалар (жыныс клеткалары) түзілген кезде аутосомдық (21,18,15 жұп) хромосомдардың дұрыс ажырамауының салдары болып табылады. Трисомия Х – синдромы әйелдерде екздеседі.Оларда жыныс бездері жетік дамымайды, дене және ақыл ой дамуында кемістіктер байқалмайды. Даун синдромы — өте жиі таралған аутосомалық ауру, Ол жаңа туылған 700 баланың бірінде кездеседі. Бұл аурау үшін ақыл – ой мешеулігі, алақан дерматоглификациясының (қатпарлығының) өзгеруі және моңғолдық көз қиығы тән, осыған байланысты аурулар өзара ұқсас болады. Даун синдромына душар болғандардың өмірі қысқа болады. Олардың кейбіреулері жыныстық жетілу жасына дейін өмір сүруі мүмкін, бірақ олардың ұрпақ қалдыруы тым сирек болады. Аурулардың жасушаларында 47 хромосома орналасады. (21 жұптың бір хромосомасы артық болады). Кейде аурудың хромосома жиынтығы 46 хромосомаға тең болады, бірақ мұнда 21 жұп хромосомасы артық жәнеде оның біреуі қысқа иықтарымен қосылған D тобы хромосомасының бірінің қысқа иықтарымен қосылған (D/G гетеро зиготалы Робертсон транслокациясы). 21 хромосома бойынша трисомик (ұрпақтардың 4/5 бөлігі) спонтанды түсіктерде өледі.Туылғандардың 31,1%-ы бір жасқа жетпей, 46,6%-ы үшінші жылдың аяғында қайтыс болады.Олар респираторлық инфекцияларға, жүрек және рак ауруларына тез шалдығады. Рессесивті гендер анықтайтын ауруларға альбинизм, гемофилия, фенилкетонурия, алкапонурия, т.б. жатады. Африка мен Азияның кейбір аудандарында кездесетін қан ауруы анықталды.Бұл ауруда эритроциттердің (қанның қызыл түйіршіктерінің) пішіні орақ тәрізді болады.Себебі, құрамындағы гемоглобин атты белок электронеатралды болады да эритроцидтер бір бірімен жабысып, оттегін тасымалдау қасиетін жоғалады.Ауру балалар ата енесі сау семьяларда тууы мүмкін.Егер әкесі де, шешесі де ауру тудвратвн ген бойынша гетерозиготалы болатын болса, олардың гаметаларының тең жартысы дұрыс хромосомды болса, қалған жартысында мутанттық ген болады.Демек, ¼ жағдайда 25% өзгерген гендері бар гаметалар ұрықтануы мүмкін.Бұл жағдайда ауру бала туады. Кейбір белгілер жыныстық хромосомдармен тіркесіп тұқым қуалайды.Мысалы,құлақ қалқаншасының жүнді болуы (гипертрихоз) әкесінің ұлына 100% беріледі.Гипертрихоздың гені У – хромосоммен тіркесіп тұқым қуалайды.Ер бала У – хромосомды тек әкесінен алады, сондықтан ол ген балаға толық беріледі. Х – хромосоммен тіркесіп тұқым қуалайтын белгілердің ұрпақтан ұрпаққа берілуінде геннің доминантты немесе рецессивті болуына байланысты ерекшкліктері болады. Мысалы, гемофелия (қанның ұйымауы) ауру шешесінен ұлына беріледі. Шешесі ауырмайды тек ұлдарына тасымалдап береді.Гемофилия гені – рецессивті, жыныстық Х – хромосоммен тіркесіп тұқым қуалайды. Егер еркектің (ХⁿУ) Х – хромосомында гемофилия гені (Һ) болса, онда ол ауруға шалдығады, себебі ноың бір ғана Х – хромосомы бар, ал У – хромосомда қанның ұюына әсер ететін ешбір ген болмайды.Сондықтан Х – хромосомның құрамындағы рецессивті ген (Һ) өзін көрсетеді.ХⁿХⁿ кароитині бар әйел гемофилиямен ауырмайды, себебі оның бір Х – хромосомында гемофилия гені (Һ) болса, екінші Х – хромосомда доминанттық, қанның қалыпты ұюын реттейтін ген (Н) бар.Сондықтан, гемофилия генінің әсерін доминанттылық ген басып, байқалтпайды.Гемофилия тәрізді дальтонизм де (қызыл және көк түсті анықтай алмау) Х – хромосоммен рнцессивтік типте тіркесіп тұқым қуалайды. 42. Жұмсақ бидайдың масағы түкті, қылтанақсыз түрімен масағы түксіз, қылтанақты өсімдігімен будандастырғанда, барлық өсімдіктердің масағы түкті және қылтанақсыз болған. Ал, екінші ұрпақта: 90 өсімдік масағы түкті және қылтанақсыз; 24 түксіз, қылтанақсыз; 33 түкті және қылтанақсыз; 11 түксіз, қылтанақсыз өсімдіктерге ажыраған. Белгілер қалай тұқым қуалайды және бастапқы өсімдіктер мен F1 өсімдіктерінің генотиптері қандай? F2 өсімдіктерінің қанша бөлігі гомозиготалы масағы түксіз және қылтанақсыз болады? Жауабы: ААвв х ааВВ Ғ1 де АаВв түкті, қылтанақты Ғ2-де 12:4 43. Кроссинговер. Хромосомалардың айқасуының генетикалық дәлелдемесін нақты мысалдармен келтіре отырып, дәлелдеңіз және себебін түсіндіріңіз. Гендер тiркесу құбылысы ашылғаннан кейiн Т.Морган мен оның қызметтестерi бiр жұп гомологты хромосомада орналасқан гендердiң тұтасымен тұқымқуалап тiркесу топтарын құрайтындығын анықтады. Белгiлi бiр топқа жататын гендердi будандастыру кезiнде бiр-бiрiмен тiркесетiндiгi байқалды, ал әртүрлi топтарға жататын гендер бiр-бiрiне тәуелсiз тұқым қуалайды. Жүргiзiлген зерттеулердiң нәтижесi гендердiң тiркесу топтарының саны хромосомалар жұптарының санына сәйкес келетiндiгiн көрсеттi. Мысалы, дрозофила шыбынында 4 жұп хромосома және 4 тiркесу тобы, арпа мен бұршақта 7 жұп хромосома және 7 жұп тiркесу тобы, жүгерiде 10 жұп хромосома және 10 жұп тiркесу тобы, қызанақта (томат) 12 жұп хромосома және 12 жұп тiркесу тобы бар. Кейiннен бұлардан басқа да көптеген өсiмдiктер мен жануарларда тiркесу топтары зерттелдi. Адамның 23 жұп хромосомаларына сәйкес келетiн 23 тiркесу топтарының болатындығы анықталды. Осы күнге дейiн тiркесу топтарының саны хромосома жұптарының санынан артық болып келетiн бiрде-бiр организм табылмаған. Әрбiр жеке организм әртүрлi гендердiң көптеген аллельдерi бойынша бiр-бiрiнен айырмашылықтары бар аталық және аналық гаметалардың қосылуы нәтижесiнде пайда болады. Жоғарыда аталып көрсетiлгендей екi тiркескен гендер бойынша айырмашылықтары бар организмдердi будандастырғанда: Дегенмен толық тiркесу құбылысымен қатар заңды түрде толымсыз тiркесу құбылысы да болып отырады. Толымсыз тiркесу құбылысы жағдайында генотипi Кроссинговер тек гендер гетерозиготалы ( Хромосомалардың айқасуы туралы, пайда болған рекомбинанттардың жиiлiгiне генетикалық талдау жасау негiзiнде ғана, яғни мейоздағы хромосомаларға цитологиялық зерттеулер және гендердiң жаңа үйлесiмi бар зиготаларға талдаулар жүргiзу арқылы ғана пiкiр айтуға болады. Тiркесу және айқасуға мысал ретiнде К.Гетчинсонның жүгерiмен жүргiзген тәжiрибесiн қарастырайық. Тұқымы тегiс, боялған өсiмдiкпен (CCSS) тұқымы бұдырлы боялмаған өсiмдiк (ccss) будандастырылған. Тұқымының тегiстiгi (С) мен бояуы (S) доминантты белгiлер, сондықтан бiрiншi ұрпақта барлығы да боялған тегiс тұқымды (CcSs) болады. Осындай тұқымдардан өсiп шыққан өсiмдiктер рецессивтi (ақ түстi бұдырлы) дарабастармен будандастырылған: Боялған тегiс ( Ақ түстi бұдырлы ( Боялған бұдырлы ( Ақ түстi тегiс ( Басқаша айтқанда, бастапқы екi түрдiң тұқымдары олардың барлық сандық мөлшерiнiң 96,4%-iн, ал тұқымдарының қалған екi тобы (кроссинговердiң нәтижесiнде пайда болған) 3,6%-iн құрайды. Бұл тәжiрибенiң нәтижесi, шындығында гендер тiркесуiнiң бар екендiгiн, бiрақ кейде кроссинговердiң салдарынан оның аздаған мөлшерде өзгеретiндiгiн, яғни мейоз кезiнде ұқсас гомологты хромосомалардың арасындағы бiркелкi бөлiмдерiнiң алмасатындығын көрсеттi. Ұқсас гомологты хромосомалардың бiрдей бөлiмдерiнде орналасатын гендер олардың бiреуiнен екiншiсiне ауысып отырады. Гомологты хромосомалардағы гендер хромосомалардың айқасуына байланысты орын алмастырады. Кроссинговерге ұшыраған хромосомалары бар гаметалар кроссоверлi, ал кроссинговерге ұшырамағандар кроссоверлi емес деп аталады. Ажырауға талдау жасау кезiнде, екiншi будандастыруда кроссинговер жағдайында әртүрлi класты дарабастардың белгiлi бiр сандық ара қатынаста болуы өзiне көңiл аудартады. Кроссоверлi емес гаметалардан пайда болған бастапқы ата-аналық белгiлердiң екi комбинациясы талдаушы будандастырудың нәтижесiнде алынған ұрпақта сан жағынан алғанда тең (41,5%) болып шығады. Сонда ата-анаға сай келетiн кроссоверлi емес гендердiң b vg және b+ vg+ жиынтығы 83% құрайды. Ал жаңа комбинациялар, яғни b vg+ және b+ vg рекомбинациялары 17% құрайды. Басқа тәжiрибеде ата-аналар ретiнде дәл сондай белгiлерi бар, бiрақ басқа үйлесiмдiлiктегi шыбындар алынған: қара денелi қалыпты қанатты ( 44. Кроссинговерге әсер етуші факторларды атап көрсетіңіз. Хром/ң айқасуының өзі күрделі биохим, физиол/қ процес болғ/н сыртқы ортаның, жалпы орг мен жеке клетканың функционалдық жағд ж/е өзінің ішкі құрылымына, яғни генотипіне тығыз байлн. Көптеген жану/р мен өсімд кроссинговер аталық ж/е аналық жын. клетка/ң мейоздық бөлінуі кезінде п.б. Ал жәндіктерде (жібек құрты) хром айқасуы тек гоморфты жыныста ж.а. Крос. мөлшері орг/ң жасына да байлн. Жас орг ол жоғары, ұлғайған сайын мөлшері кемиді. Кей орг генотипінде мыс, қарабидайда хиазманың п.б. бақылап отыратын ген бары анықт. Сонымен қатар крос/і орг әр түрлі факторлармен әсер ету арқ қолдан жасауға да болады, оны индукциялық кроссинговер д.а. Мыс, радиоактивті сәулелермен, хим агенттермен н/е жоғары или төменгі темп әсер ету арқылы. ДНҚ-ң құрылымы бұзылады. Соған байлн кроссинговер мөлшері өзгереді. Сөйтіп кроссинговер нәт гендер рекомбинацияланады. 45. Көп аллельдiлiк және қан тобын анықтау генетикасын түсіндіріңіз. Көп аллельділікті мысалдар келтіру арқылы дәлелдеңіз. Бұрынғы тарауларда қарастырылған фенотип бойынша ажыраулардың нәтижелерi геннiң екi аллельдiк жағдайда мысалы, бұршақта ген А-а және В-в болатындығы ескерiле отырып түсiндiрiлдi. Бiрақ, бiраз гендердiң бiрнеше немесе одан да көп аллельдерi болады. Мұны геннiң көп аллельдiлiгi деп айтады. Көп аллельдiлiк жағдайында гаметада немесе спорада белгiлi бiр геннiң тек жалғыз ғана аллелi, ал жоғары сатыдағы өсiмдiктердiң және жануарлар мен адамның дене клеткаларында ондай геннiң екi аллелi болатындығын атап өткен жөн. Сондықтан көп аллельдердiң ажырауы әрқашанда моногибридтi болып қалады. Ендi көп аллельдiлiкке бiраз мысалдар келтiрейiк. Үй қояндарында жүнi мен көзайнасы қабықшасының түстерiне жауапты С генiнiң көп аллельдерi болатындығы белгiлi. Бұл жағдайда әрбiр атап көрсетiлген аллельдер өзiнен кейiнгiлерге басымдылық көрсетiп отырады, оны былайша жазуға болады С>cch>ch>c. Жабайы типке тән С аллелi қалған үшеуiне қарағанда доминантты болып келедi. С генi бойынша гомозиготалы немесе жоғарыда көрсетiлген кез келген аллельдермен С генi арқылы гетерозиготалы үй қояндарының жүнi жабайы қояндардiкi сияқты сұр (агути) түстi болып келедi. cch аллелi бойынша гомозиготалы дарабастардың түсi шиншилланың түсiне келедi және жабайы түрiне қарағанда бiршама ашықтау болады. Ал cch және ch немесе с аллельдерiне гетерозиготалыларда түсi ашық сұр (шиншилла мен ақтың аралығында) болып келедi. Олай болса cch аллелi ch және с аллелдерiне толымсыз доминанттылық көрсетедi. Гомозиготалы chch пен гетерозиготалы cchс болып келетiн қояндардың фенотиптерi гималай тәрiздi, яғни башпайлары мен құйрығының ұшы, құлақтары және тұмсығы қара да, денесiнiң қалған жерлерi ақ болады. Ал С аллелi бойынша гомозиготалыларда пигмент мүлдем болмайды, олар таза ақ түстi келедi. Адамда болатын көпаллельдiлiктiң мысалы ретiнде қан топтарын анықтайтын АВО аллельдерiн алуға болады. Адамда қанның төрт тобы болады: О,А,В және АВ. Олар бiр геннiң үш түрлi аллельдерi арқылы анықталады: IA, IB және i. IA және IB аллельдерi i аллелiне қарағанда доминантты, бiрақ олар бiр-бiрiне кодоминантты болып келедi, яғни IA және IB аллельдерiнiң екеуi де гетерозиготаларда бiрдей мөлшерде байқалады. Адамда түрлi қан топтарының болатындығы анықталғанға дейiн қан құю кезiнде ауыр тiптi кейде қазаға ұшыраған жағдайлар да болып тұрды. Оның себебi тобы сәйкес келмейтiн қанды құйған кезде құйылған қанның эритроциттерi (қызыл түйiршiктерi) аглютинацияланып (желiмделiп) қалады, соның салдарынан қан құйылған адамның қан тамырлары бiтелiп қан жүрмейдi. Мұның мәнi мынада: АВ тобына жататын эритроциттерде екi антиген болады, яғни А тобында - А антигенi, В тобында - В антигенi, ал О тобында А және В антигендерi болмайды. Осы төрт түрлi топтардың қан сарысуларында мынадай айырмашылықтар болады: О тобында жәнедеп белгiленетiн екi антидене, ал А тобында антиденесi, В тобында антиденесi болса, АВ тобының сарысуында мен антиденелерi болмайды. Адамдағы қан топтарының тұқымқуалау заңдылығының тек қан құюда ғана емес, кейбiр даулы мәселелердi мысалы, баланың әкесi кiм екендiгiн анықтауда да маңызы бар. 46. Көзі қалыпты көретін әйелдің әкесі дальтоник болған. Екі бауыры және шешесі жағынан ағасы гемофилия ауруымен ауырады, ал қызы дальтоник. Отбасы мүшелерінің генотиптерін анықтаңыздар. Қызының гемофилия ауруын тасымалдаушы екендігін дәлелдеңіз? Жауабы: D//d x d/еркек, қыздар тасымалдаушы болады, яғни D//d жыныспен тіркесіп т.қ. 47. Күздік жұмсақ бидай дакылының гүл қабығының түкті және түксіз түрлерін өзара шағылыстырғанда, F1 ұрпақтарының гүл қабықтары түкті, ал F2-де алынған өсімдіктердің 160-ы түкті және 45-і түксіз болған. Бірінші ұрпақта алынған дақылдардың қабыршақтары түксіз түрлерімен шағылыстырғанда, 110 түкті және 98 түксіз өсімдіктер алынған. Белгі қалай тұқым қуалайды? Бастапқы және F1 өсімдіктерінің генотиптерін анықтаңыздар. F1 алынған дақылдарды бастапқы аталық-аналық қабыршақтары түкті түрімен шағылыстырса қандай нәтиже алынар еді? Жауабы: АА х аа; Аа х Аа Ф 3:1; Аа х аа Ф1:1; Аа х АА Ф 1:1; 48. Қалыпты көретін отбасында, дальтонизммен ауыратын бір ұл және қалыпты көретін екі қыз болған. Ұлының әйелі мен қыздарының жұбайларының көру қабілеті қалыпты. Ұлы жағынан бір немересі қалыпты, ал, бірінші қызының ұлы дальтонизммен ауырған, ал қызы сау болған. Ал, екінші қызының қалыпты көретін бес қызы болған. Осы мәліметтерді пайдалана отырып, шежіре құрып, генетикалық талдау жасаңыздар. Жауабы: белгі аутосомды рецессивті тұқым қуалайды. Егер ата- аналарының біреуі осы белгіні тасымалдаса, белгі балаларвнда көрінбеуі мүмкін н/е ұрпақтарында байқалуы мүмкін. Пробанд белгісі аа. 49. Қандарының ұю қабілеті қалыпты (сау), А қан тобына жататын әйел В қан тобына жататын сау кісіге тұрмысқа шығады. Бұл отбасында үш бала туылған: Айман – сау, қан тобы А; Ерлан – сау, қан тобы О; Шыңғыс – гемофилик, қан тобы А. Әйелдің әке-шешесі сау болғаны белгілі, шешесінің қан тобы О, әкесінің қан тобы АВ болған. Ал күйеуінің әке–шешесі сау, қан топтары А және В болған. Шыңғысқа гемофилия ауруы кім арқылы берілгенін және барлық отбасы мүшелерінің генотипін анықтаңыздар. Жауабы: P ♀ IAIOXH Xh x ♂ IAIOXH Xh Айман- IAIOXH Xh, Ерлан- IOIOXH Y , Шыңғыс- IAIO XhY 50. Қоян құлақтарының ұзындығы 28 см, ал, басқа тұқымды қоян құлағының ұзындығы – 12 см. Қоян құлақтарының ұзындығы бойынша айырмашылық екі жұп кумулятивті гендердің әсеріне тәуелді деп есептеледі. Құлағының ұзындығы 28 см болып келетін қояндардың генотипі D1D1D2D2, ал, 12 см қояндардың генотипі – d1d1d2d2 болып келеді. Демек, әрбір доминантты ген құлақ ұзындығының 4 см-ге ұзаруына әсер етеді. Құлақ ұзындықтары әртүрлі қояндарды өзара шағылыстырудан, F1-де 14, ал, F2 – 32 көжектер туылған. F1 және F2-де алынған көжектердің генотиптерін анықтау керек? Жауабы: P ♀ D1 D1 D2 D2 x ♂ d1 d1d2d2 Г: D1 D2 x d1 d2 F1 D1 d1 D2 d2 Р (F1) ♀ D1 d1 D2 d2 x ♂ D1 d1 D2 d2 Г: D1 D2, D1 d2 , d1 D2 , d1 d2 x D1 D2, D1 d2 , d1 D2 , d1 d2 F2 1 D1 D1 D2 D2 – 28см
4 D1 D1 D2 d2 – 24 см D1 d1 D2 D2
D1 d1 D2 d2 6 D1 D1 d2 d2 -20 см d1 d1 D2 D2
4 D1 d1 d2 d2 -16 см d1 d1 D2 d2 1 d1 d1 d2 d2 -12 см 51. Моно- және дигибридті будандастыру дегеніміз не? Моно- және дигибридті будандастыруға талдаушы будандастыру арқылы мысал келтіріп, салыстырыңыз. Қарама-қарсы бiр жұп белгiлерi бойынша ажыратуға болатын ата-аналық формалар будандастырылса, оны моногибридтi деп айтады. Мысалы Мендель тәжірибелерi үшiн альтернативтi жетi белгiлерi бойынша ажыратылатын бұршақтың әртүрлi сорттарын тандап алды: тұқымы сары немесе жасыл, тұқымы тегiс немесе бұдыр, тұқым қабығы сұр немесе ақ, бойы биiк немесе аласа т.б. Дигибридтiк будандастыруларды бiр-бiрiне тәуелсiз екi моногибридтi будандастыру деп қарауға да болады. Әрбiр моногибридтiк будандастыруда доминантты белгiлерi бар дарабастардың пайда болу мүмкiндiгi 3/4 бөлiгiне, рецессивтi белгiлерi бар дарабастардың пайда болу мүмкiндiгi 1/4 бөлiгiне тең болса, онда екi белгiнiң бiр мезгiлде пайда болу ықтималдылығы төмендегi көбейтiндiлерге тең болады: сырты тегiс сары дән 3/4 х 3/4 = 9/16, сырты тегiс жасыл дән 3/4 х 1/4 = 3/16, сырты бұдыр сары дән 3/4 х 1/4 = 3/16, сырты бұдыр жасыл дән 1/4 х 1/4 = 1/16. Бұл алынған сандар Пеннет торындағы 9: 3: 3: 1 қатынасына сәйкес келедi және белгiлердiң фенотип бойынша ажырауын көрсетедi. Мендельдiң тәжiрибесiнде F2 -дегi 556 тұқым фенотип бойынша осылай ажырады. Сонымен гибридтердiң екiншi ұрпағы F2 фенотиптерi бойынша9: 3: 3: 1 қатынасында, ал генотиптерi бойынша 1:2:1:2:4:2:1:2:1 қатынасында ажырайды. Бұл ажырауды Пеннет торынан көруге де болады. Генотиптiк ажырауды математикалық әдiспен де есептеп шығаруға болады. Ол үшiн дигибридтi ажырауды бiр-бiрiне тәуелсiз екi моногибридтi ажыраулар көбейтiндiсi ретiнде қарастыру керек: (1АА: 2Аа:1аа) х (1ВВ: 2Вв: 1вв). Сонда генотиптерi бойынша ажырау 1ААВВ: 2ААВв: 1ААвв:2АаВВ: 4АаВв: 2Аавв:1ааВВ: 2ааВв:1аавв болып шығады. Дигибридты будандастырулардың нәтижесiнен жалпы мынадай қорытынды шығады: бiр-бiрiне тәуелсiз ажырау гендердiң екi жұбында да жүредi, осыған байланысты екi жұп гендерi бойынша гетерозиготалы организм бiрдей сандық қатынаста гаметалардың төрт сортын түзедi, оның екеуiндегi гендер үйлесiмi бастапқы ата-аналарындағы сияқты болады да, ал қалған екеуiндегi гендер керi үйлеседi (рекомбинациялық) болады. Мендель ашқан, кейiннен сан алуан организмдермен жүргiзiлген тәжiрибелерде дәлелденген осы заңдылық тәуелсiз тұқым қуалау немесе тәуелсiз комбинациялану заңы деп аталады. 52. Моногибридті шағылыстыру. Мендельдің моногибридті шағылыстыруды талдау барысында белгілердің негізгі тұқымқуалау заңдылықтарына нақты мысалдар арқылы Мендель заңдылықтарын дәлелдеңіз. Альтернативтi немесе қарама-қарсы бiр жұп белгiлерi бойынша ажыратуға болатын ата-аналық формалар будандастырылса, оны моногибридтi деп айтады. Мысалы Мендель тәжірибелерi үшiн альтернативтi жетi белгiлерi бойынша ажыратылатын бұршақтың әртүрлi сорттарын тандап алды: тұқымы сары немесе жасыл. будандасудан алынған F1 гибридтерiнде альтернативтi жұп белгiнiң тек бiреуi ғана көрiнiс бередi. Мұндай белгiнi Мендель доминантты деп атады. Ол жұп белгiлердiң iшiнен: тұқымның сары түсi жасыл түске қарағанда доминанттылық көрсететiнiн байқады. Бiрiншi гибрид ұрпақта көрiнбеген белгiлердi Мендель рецессивтi белгiлер деп атады. Тұқым қуалауға талдау жасағанда бiрiншi гибрид ұрпақ бiрдей пiшiндi, бiр типтес болғандықтан бұл құбылыс Мендельдiң бiрiншi заңы немесе бiрiншi ұрпақ гибридтерiнiң бiркелкiлiк заңы деп аталады. Бұл заңды доминанттылық ережесi деп те атайды. Бұл барлық өсiмдiктерге, жануарларға, адамға да тән жалпы кұбылыс. бiрiншi гибрид ұрпағы F1 өздiгiнен тозаңданса, онда олардың екiншi F2 ұрпағында атасының да, анасының да белгiлерi көрiнетiн болады. Мысалы, бұршақтың сары(А) және жасыл(а) тұқым жарнақтары бар түрлерiн будандастырудан алынған бiрiншi ұрпақтың түсi сары(Аа) болады. Ал осы F1 гибридтерiн өздiгiнен тозаңданудан алынған F2 ұрпағында сары және жасыл тұқымдары бар өсiмдiктер пайда болады. Яғни бiрiншi будан ұрпақта көрiнбеген белгiлер (жасыл түс) екiншi ұрпақта көрiнедi. Доминантты және рецессивтi белгiлер F2 ұрпағында белгiлi бiр сандық ара қатынаста болады. Мендельдiң тәжiрибесiнде F1 -де алынған Аа-258 өсiмдiктiң өздiгiнен тозандануынан F2 -де 8023 тұқым алынды. Осы тұқымдардың ¾(1АА, 2Аа) бөлiгi (яғни 6022) сары ал, (аа)1/4 бөлiгi (яғни 2001) жасыл түстi болып шыққан. Сонымен F2 -де алынған доминантты және рецессивтi белгiлерi бар тұқымдардың сандық арақатынасы 3:1 болды. Бiр жұп белгiлердiң осындай арақатынаста ажырауы Мендельдiң екiншi заңы немесе ажырау заңы деп аталады. Жүргiзген зерттеулерiне талдау жасаудың негiзiнде Мендель қорытынды жасады: рецессивтi бастамалар (задатки) гетерозиготалы организмде жоғалмайды, өзгерiске ұшырамайды және көбею кезiнде дәл сондай рецессивтi бастамалармен кездескен кезде, яғни келесi ұрпақтарда қайтадан көрiнедi. Мендельдiң бұршақпен жүргiзген тәжiрибелерiнен байқалған доминанттылық құбылысын толық доминанттылық. Егер F1 -дегi гибридтердiң белгiлерi ата және аналық формалардың ешқайсысына толық ұқсамай аралық сипатта болса оны толымсыз доминанттылық деп атайды. Қызыл гүлдi өсiмдiктердi (АА) ақ гүлдi өсiмдiктермен (аа) будандастырғанда F1 -де генотиптерi гетерозиготалы (Аа) гүлдерi қызғылт түстi ұрпақ алынған. Реципрокты будандастыру деп доминантты және рецессивтi белгiсi бар организмдерi аналық ретiнде де, аталық ретiнде де пайдалануды айтады.Мыс: АА х аа - тура, ал аа х АА - керi будандастыру. қайыра бұдандастыру немесе беккросс деп – (Аа хАА; Аа х аа). Талдаушы будандастыру деп - (Аа х аа) 53. Митоз, мейоз кезеңдері. Мейоздың биологиялық маңызын сызба түрінде көрсетіңіз. Митоз – дене клеткасының бөлінуі. Бөлiнуге кiрiскен кез келген клетка бiрқатар өзгерiстерге ұшырайды, сол өзгерiстерден клеткалық циклдi құрайды. Клеткалық цикл төрт кезеңнен тұрады: синтезге дейiнгi (G1), ДНҚ синтезi (S), синтезден кейiнгi (G2) және митоз (М).ядроның өзiнiң бөлiнуi - кариокинезден және цитоплазманың бөлiнуi – цитокинез. Интерфазаның G1 - кезеңiнде клетка тiршiлiгi үшiн аса қажеттi заттар - нуклеотидтер, аминқышқылдары, ферменттер т.б. синтезделедi. S - Клеткадағы ДНҚ мөлшерi екi есе артады. Синтезден кейiнгi G2 - кезеңiнде РНҚ синтезделеді. профазада хромосомалар ширатылады, қысқарады және жуандайды. Профазаның соңына қарай ядрошықтар жойылады, ядролық мембрана еридi, сөйтiп хромосомалар цитоплазмада жатқандай болып шығады. Метафазада хромосомалар экватор бағытына қарай жылжиды. Хромосомалардың жылжуы центромера учаскелерiне бекiген ахроматин жiпшелерi (ұршық тәрiздi) арқылы жүзеге асады. Хромосоманың бір жазықтықта орналасуы метафазалық пластинка д.аталады. анафазада хромосомалар қарама-қарсы полюстерге жылжиды. Телофазада хромосомалар полюстерге толық ажырайды. ядролық мембрана түзiледi, ядрошықтар пайда болады, кариокинез, цитокинез жүреді. 2 клетка пайда болады. 46-46дан. Мейоз – жыныс клеткасының бөлінуі. Мейоз нәтижесiнде гаметалар - жұмыртқа клеткалары мен сперматазоидтар яғни жыныс клеткалары пайда болады. Ол 2 бөлінуден тұрады: редукциялық(мейоз1) және эквациялық(мейоз2). Редукциялық бөлiну I-шi профазадан басталады. Ол 5 кезеңге бөлiнедi: Лептотена Хромосомалар әлсiз, өте нәзiк әрi ұзын жiпшелер түрiнде көрiнедi. Зиготена -көлемi бiрдей хромосомалар коньюгацияланып қосылады. Коньюгацияланушы гомо-логты хромосомаладың әр жұбы бивалент түзедi. гомологты хромосомалардың қосы-луын синапсис деп атайды. Пахитена –хромосомалар толық ширатылып, жуандап, қысқарады.Биваленттердiң саны хромосомалардың гаплоидты санына (n) тең болады. Диплотенада гомологті хромосомалар бір-бірін тебеді, бөлім алмастыру-конъюгация жүреді. Хроматидтер арасында хиазма д.аталатын Х әрпіне ұқсас фигуралар п.б. Диакинезде ширатылу күшейе түседi де, хиазмалардың саны азайып, биваленттер ядроның шет жағына орналасады. I-профазада Ядроның қабығы мен ядрошықтар жойылады. I-шi метафазада биваленттер экватор жазықтығына орналасады. I-шi анафазада хромосомалар қарама-қарсы полюстерге таралады. олар бiр-бiрiне тәуелсiз қозғалады. I-шi телофазада гаплойдты ядроның клеткада түзілуі(2 ядро). Эквациялық бөлiну басталады.II-шi профазада хромосомалар ширатылып,тығыздалады. II-шi метафазада хромосомалардың центромералары экватор жазықтығына орналасады. II-шi анафазада екi еселенген центромералар бiр-бiрiнен ажырайды нәтижесiнде жаңа түзiлген хроматидтер әр полюстерге тарайды. II-шi телофазада гаплоидты төрт ядролар түзiледi. Содан соң цитокинез жүредi де, нәтижесiнде төрт клетка пайда болады. Мейозда кезiнде хромосомалардың әр жүбының тәуелсiз таралуы, бөлiмдер алмасуы және сол хромосомалардың айқасуы нәтижесiнде түзiлетiн клеткалардың хромосомалар жиынтығы ұқсас болмайды. Мейоз комбинативтiк өзгергiштiктi камтамасыз етедi. онымен, жануарлар мен өсiмдiктердiң жынысты көбеюiнде ұрпақтар арасындағы сабақтастылық тек жыныс клеткалары арқылы ғана қамтамасыз етiледi. 54. Митохондриялы тұқым қуалауға нақты мысалдар келтіре отырып, дәлелдеңіз. Митохондриялар пластидтер сияқты жартылай автономды органеллалар болып табылады және олар бактериялар мен көкжасыл балдырлардан басқа барлық клеткаларда кездеседі. Митохондрия - клетканың тыныс алуы кезінде энергия бөліп шығаратын органоид. Митохондриялық геномдардың көпшілігі тұйық сақиналы, жоғары спиральды дуплексті ДНҚ түрінде болады. Олар көбінесе ұқсас функцияларды кодтайды. Бірқатар организмдердің митохондрияларында, соның ішінде кейбір саңырауқұлақтар мен қарапайымдылардың геномы сызықша түрде болады. Митохондриялық геном Мендельдік (ядролық) геномдармен салыстырғандағы басты айырмашылығы: олар бөлінбейді және қайтадан бірікпейді. Оның орнына ол анадан қызына көшіріледі немесе клондалады. Мутацияларды санамағанда, қазіргі аналық ұрпақтардағы генетикалық ауруы немесе кездейсоқ SNP (бір нуклеотидтің полиморфизмі) осыдан 300-400 жыл бұрынғылармен бірдей. Митохондриялардағы ядродан тыс тұқым қуалау Митохондриялық геномда 37 ген бар (13-і белоктар үшін (көппептиды), 22-сі РНҚ тасымалы үшін (tRNA) және біреуі үлкен, біреуі кішкентай рибосомалық РНҚ бірліктері үшін). Олардың біраз бөлігі АТФ-тың түзілуіне және реттелуіне қатысады. 55. Мутациялық өзгергіштік. Мутация типтерін нақты мысалдар арқылы сипаттаңыз. Мутация деп хромосомалардың немесе гендердің кенеттен өзгеруін айтады. Мутацияның пайда болуы жөне қалыптасуы мутагенез деп аталады. Ал тұқым қуалайтын өзгерістерді тудыратын факторларды мутаген деп атайды. Мутациялық процестің нәтижесінде жаңа қасиеттерге ие болған дараларды мутанттар дейді. Мутациялық өзгергіштік тұқым қуалайды. Олардың пайда болуы бірнеше кезеңдерді қамтитын күрделі процесс. "Мутация" деген терминді 1901 жылы Голландия генетигі Гуго де Фриз "Мутациялық теория" деген еңбегінде енгізді. Жалпы мутацияны эволюциялық процестің алғашқы материалдық негізі деп есептеген жөн. Мутациялық өзгергіштіктің барлық түрлері тірі организмдерге тән. Организмдердің ерекшеліктеріне: фенотипінің және генотипінің өзгеруіне қарай мутациялар: морфологиялық, физиологиялық, гендік, хромосомалық жөне геномдық болып бөлінеді. Морфологиялық мутацияларға өсімдіктер мен жануарлар мүшелерінің тұқым қуалайтын белгілерінің өзгеруі жатады. Мысалы, өсімдіктердің бойының биіктігі, гүлдер мен гүл шоғырлары түсінің, жапырак тақтасы мен жемістерінің, сондай-ақ тұқым түрлерінің өзгеруін айтады. Жануарларда: терісінің түсі, аяғының қысқалығы, кауырсынның болмауы. Жәндіктерде: денесінің, көзінің түсінің өзгеруі, қанатының мөлшері мен пішінінің езгеруі. Физиологиялық мутацияларға есімдіктер мен жануарлардың өнімділігінің төмендеуі немесе артуы, сонымен қатар олардың сыртқы ортаның қолайсыз жағдайларына, (суык, ыстыққа) ауруға төзімсіздігі мен тезімділігін көрсететін қасиеттері жатады. Физиологиялық мутацияларға летальді және сублетальді мутациялар кіреді. Мутациялық өзгергіштік 2-ге б/і. Секірмелі (спонт) -сыртқы ортаның н/е қалыпты физиоло, биохим процес/ң өзгеруі нәт. өздігінен п.б. мут. Индкуциялық (жасанды) -радиоактивті сәулелер, хим. заттар н/е темп әсер ету нәт. п.б. мут. 56. Негізгі объект - дрозофиланың биологиясы, морфологиясы және көбеюіне анықтама беріңіз, сызба түрінде тәжірибеде қолданылатын мутантты линияларды сипаттаңыз. Дата добавления: 2015-12-16 | Просмотры: 3441 | Нарушение авторских прав |
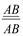 х
х  , дигетерозиготалы форма
, дигетерозиготалы форма 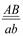 пайда болады. Толық тiркесу жағдайында дигетерозигота тек екi түрлi ғана гаметалар түзедi: АВ және ав. Талдаушы будандастыру кезiнде зиготалардың екi класы пайда болады:
пайда болады. Толық тiркесу жағдайында дигетерозигота тек екi түрлi ғана гаметалар түзедi: АВ және ав. Талдаушы будандастыру кезiнде зиготалардың екi класы пайда болады: 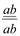 , олардың әрқайсысы 1: 1 тең болады. Екi кластың да дарабастары өздерiнiң ата-аналарының белгiлерiн қайталайды. Мұндай көрiнiс талдаушы будандастыру кезiнде дигибридтi ажырау емес моногибридтi ажырауды еске түсiредi.
, олардың әрқайсысы 1: 1 тең болады. Екi кластың да дарабастары өздерiнiң ата-аналарының белгiлерiн қайталайды. Мұндай көрiнiс талдаушы будандастыру кезiнде дигибридтi ажырау емес моногибридтi ажырауды еске түсiредi. гетерозиготалы дарабастарды рецессивтi
гетерозиготалы дарабастарды рецессивтi 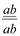 формалармен будандастырғанда олардың ұрпағында жаңа белгiлерi бар дарабастар пайда болады:
формалармен будандастырғанда олардың ұрпағында жаңа белгiлерi бар дарабастар пайда болады:  және
және  . Мұндай жаңа фенотиптер рекомбинанттар деп аталады. Ажырау барысында зиготалардың жаңа кластарының пайда болуы екi ген бойынша гетерозиготалы формалардың гаметогенезi процесiнде тек қана АВ және ав емес, сонымен қатар Ав және аВ гаметалары да түзiлетiндiгiн көрсетедi. Олай болса F1 буданына бiр ғана хромосомамен келген гендер, одан гамета түзiлу кезiнде қандай да бiр жолмен ажырап кетедi. Гендердiң жаңа үйлесiмi бар мұндай гаметалары қалай пайда болды? Демек, олар тек гомологты хромосомалардың арасында бiр-бiрiмен бөлiмдерiн алмастыру, яғни кроссинговер кезiнде пайда болуы мүмкiн. Сонымен кроссинговер ұқсас (гомологты) хромосомаларда болатын гендердiң жаңа үйлесiмiн қамтамасыз етедi. Кроссинговер құбылысы да тiркесу сияқты барлық жануарлар мен өсiмдiктерге және микроорганизмдерге тән құбылыс.
. Мұндай жаңа фенотиптер рекомбинанттар деп аталады. Ажырау барысында зиготалардың жаңа кластарының пайда болуы екi ген бойынша гетерозиготалы формалардың гаметогенезi процесiнде тек қана АВ және ав емес, сонымен қатар Ав және аВ гаметалары да түзiлетiндiгiн көрсетедi. Олай болса F1 буданына бiр ғана хромосомамен келген гендер, одан гамета түзiлу кезiнде қандай да бiр жолмен ажырап кетедi. Гендердiң жаңа үйлесiмi бар мұндай гаметалары қалай пайда болды? Демек, олар тек гомологты хромосомалардың арасында бiр-бiрiмен бөлiмдерiн алмастыру, яғни кроссинговер кезiнде пайда болуы мүмкiн. Сонымен кроссинговер ұқсас (гомологты) хромосомаларда болатын гендердiң жаңа үйлесiмiн қамтамасыз етедi. Кроссинговер құбылысы да тiркесу сияқты барлық жануарлар мен өсiмдiктерге және микроорганизмдерге тән құбылыс. және
және  х
х  . Сонда алынған ұрпақтың төмендегiдей сандық арақатынаста төрт түрлi тұқымдарының болатындығы байқалған:
. Сонда алынған ұрпақтың төмендегiдей сандық арақатынаста төрт түрлi тұқымдарының болатындығы байқалған: ) - 4032
) - 4032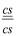 ) - 4035
) - 4035 ) - 152
) - 152 ) - 149
) - 149 ) шыбынды, сұр денелi қанаты жетiлмеген (
) шыбынды, сұр денелi қанаты жетiлмеген ( ) шыбынмен будандастырған. Содан кейiн қайтадан екi түрлi талдаушы будандастыру жүргiзiлген. Бiрiншi будандастыруда F1 -дiң аталықтары, екiншiсiнде F1 -дiң аналықтары пайдаланылды. Екеуiнде де олар қос гомозиготалы рецессивпен (
) шыбынмен будандастырған. Содан кейiн қайтадан екi түрлi талдаушы будандастыру жүргiзiлген. Бiрiншi будандастыруда F1 -дiң аталықтары, екiншiсiнде F1 -дiң аналықтары пайдаланылды. Екеуiнде де олар қос гомозиготалы рецессивпен ( ) будандастырылған. Сонда қайтадан, талдаушы будандастыру кезiнде ата-аналық және рекомбинанттық белгiлердiң арақатынасы бұрынғыдай болып шыққан. Егер F1 -дегi аталықтар алынса, онда белгiлердiң тек қана ата-аналық комбинациясы, ал егер F1 -дегi аналықтарды алса, онда 83% ата-аналық және 17% рекомбинациялық белгiлер байқалады, яғни белгiлердiң үйлесiмi алғашқы тәжiрибедегiдей ара қатынаста болады.
) будандастырылған. Сонда қайтадан, талдаушы будандастыру кезiнде ата-аналық және рекомбинанттық белгiлердiң арақатынасы бұрынғыдай болып шыққан. Егер F1 -дегi аталықтар алынса, онда белгiлердiң тек қана ата-аналық комбинациясы, ал егер F1 -дегi аналықтарды алса, онда 83% ата-аналық және 17% рекомбинациялық белгiлер байқалады, яғни белгiлердiң үйлесiмi алғашқы тәжiрибедегiдей ара қатынаста болады.