|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Понятие об адаптации. Стресс. Учение об общем адаптационном синдроме. Роль систем гипоталамус -гипофиз - надпочечники в адаптации организма к нагрузкам.Раздражимость и возбудимость клетки. Биоэлектрические явления в состоянии покоя и деятельности клетки. Значение биоэлектрических явлений в процессах передачи информации в организме. Раздражимостью называется способность живых клеток, тканей или целого организма отвечать на внешние воздействия изменением своей структуры, а также возникновением, усилением или ослаблением своей деятельности. Эти внешние воздействия называют раздражителями, ответные реакции на них клеток, тканей и всего организма - биологическими реакциями. Сам процесс воздействия раздражителя называется раздражением. Но своей природе раздражители могут быть химическими, электрическими, механическими, температурными, радиационными, световыми, биологическими и др. По своему биологическому значению для каждой клетки все раздражители делятся на адекватные и неадекватные. Адекватными называются те раздражители, которые при минимальной силе раздражения вызывают возбуждение в данном виде клеток, выработавших в процессе эволюции специальную способность реагировать на эти раздражители. Чувствительность клетки к адекватным раздражителям очень велика. Все остальные раздражители называют неадекватными. В той или иной степени способны отвечать на раздражение все живые клетки и ткани. Однако нервная, мышечная и железистая ткань в отличие от других способны осуществлять быстрые реакции на раздражения. Эти ткани получили название возбудимых тканей. К возбудимым клеткам относят и специализированные рецепторные клетки, например, палочки и колбочки сетчатки глаза. Способность нервных, мышечных и железистых клеток и тканей, а также рецепторных клеток быстро отвечать на раздражение изменениями своих физиологических свойств и возникновением возбуждения называется возбудимостью. Возбуждение - это волнообразный процесс, который проявляется в специфической ответной реакции ткани (мышечная - сокращается, железистая - выделяет секрет, нервная - генерирует электрический импульс) и неспецифической (изменение t, обмена веществ и др.)- Обязательным признаком возбуждения является изменение электрического заряда наружной оболочки или мембраны клетки. Минимальная сила раздражителя, необходимая для возникновения минимальной ответной реакции клетки и ткани, называется порогом раздражения. Он измеряется в различных физических величинах, которыми характеризуется величина раздражителя (в градусах, килограммах, децибеллах и т.д.). Минимальная сила раздражения, необходимая для возникновения возбуждения клетки и генерации потенциала действия, называется порогом возбуждения. Порог возбуждения измеряется в миллиВольтах. Любая живая клетка покрыта полупроницаемой мембраной, через которую осуществляется пассивный и активный избирательный перенос положительно и отрицательно заряженных ионов. Благодаря этому переносу между наружной и внутренней поверхностью мембраны клетки существует электрическая разность потенциалов - мембранный потенциал. Существует три отличающихся друг от друга проявления мембранного потенциала - мембранный потенциал покоя, местный потенциал и потенциал действия. Если на клетку не действуют внешние раздражители, то мембранный потенциал долго сохраняется постоянным. Мембранный потенциал такой покоящейся клетки называется мембранным потенциалом покоя. Для внутренней среды клетки потенциал покоя всегда отрицателен и равен для нервной и поперечно-полосатой мышечной ткани от -50 до -100 мВ, для эпителиальной и гладкомышечной ткани от -20 до -30 мВ. Причиной возникновения потенциала покоя является разная концентрация катионов и анионов снаружи и внутри клетки и избирательная проницаемость для них клеточной мембраны. Цитоплазма покоящейся нервной и мышечной клетки содержит примерно в 20-100 раз больше катионов калия, в 5-15 раз меньше катионов натрия и в 20-100 раз меньше анионов хлора, чем внеклеточная жидкость. В мембране клетки имеются специфические натриевые, калиевые, хлорные и кальциевые каналы, которые избирательно пропускают, соответственно, только Na+, K+, Cl- и Са2+. Эти каналы обладают воротным механизмом и могут быть открытыми или закрытыми. В состоянии покоя практически все натриевые каналы мембраны клетки закрыты, а большинство калиевых - открыто. Всякий раз, когда ионы калия наталкиваются на открытый канал, они диффундируют через мембрану. Поскольку внутри клетки ионов К+ гораздо больше, то их выходит из клетки гораздо больше, что увеличивает положительный заряд наружной поверхности мембраны. Этот выходящий поток К+ должен был бы вскоре выровнять осмотическое давление (или концентрацию) этого иона, но этому препятствует электрическая сила отталкивания положительных ионов К+ от положительно заряженной наружной поверхности мембраны. Ионы К+ будут выходить из клетки до тех пор, пока сила электрического отталкивания не станет равной силе осмотического давления K+. При таком уровне потенциала мембраны уравновесится выход и вход ионов К+ через мембрану клетки. Поскольку в состоянии покоя почти все натриевые каналы мембран закрыты, то ионы Na+ поступают в клетку в незначительном количестве и поэтому не могут возместить потерю положительного заряда внутренней среды клетки, вызванную выходом ионов К+. Избыток ионов Na+ на наружной поверхности мембраны совместно с выходящими из клетки ионами К+ создают положительный потенциал снаружи мембраны покоящейся клетки. В состоянии покоя мембрана нервных клеток проницаема несколько хуже, а мышечных клеток проницаемость несколько лучше для анионов Сl-, чем для катионов К+ Анионы Сl-, которых больше вне клетки, диффундируют внутрь клетки и несут с собой отрицательный заряд. Уравниванию концентраций ионов Cl- препятствует сила электрического отталкивания одноименных зарядов. Мембрана клетки практически непроницаема для крупных органических анионов, в частности молекул белков, анионов органических кислот. Поэтому они остаются внутри клетки и совместно с поступающими внутрь клетки ионами Cl- обеспечивают отрицательный потенциал на внутренней поверхности мембраны покоящейся клетки. При действии на клетку различных раздражителей по силе примерно в 1,5-2 раза меньше порога раздражения мембранный потенциал покоя начинает уменьшаться, т.е. происходит деполяризация мембраны клетки. С увеличением силы раздражения деполяризация мембраны нарастает. Однако, если сила раздражения не достигла порога, то прекращение раздражения приводит к быстрому восстановлению потенциала покоя. В мышечной и нервной тканях при пороговом раздражении уменьшение потенциала мембраны ограничено небольшим участком в месте нанесения раздражения и получило название местного потенциала или локального ответа. При достижении раздражения пороговой силы возникает быстрое кратковременное изменение величины и полярности заряда мембраны клетки, которое получило название потенциала действия (применяются также тещины "волна возбуждения", для нервных клеток -"нервный импульс"). Потенциалы действия всегда возникают при деполяризации мембраны нервной и поперечно-полосатой мышечной клетки примерно до -50 мВ. Причиной возникновения местного потенциала, а затем и потенциала действия является раскрытие натриевых каналов и поступление ионов Na+ внутрь клетки. При нарастании силы раздражения до пороговой этот процесс идет медленно и возникает местный потенциал. При достижении критического уровня деполяризации мембраны (примерно -50 мВ) проницаемость натриевых каналов мембраны лавинообразно возрастает. Ионы Na+ поступают внутрь клетки, что приводит не только к быстрой нейтрализации отрицательного заряда у внутренней поверхности мембраны, но и к возникновению положительного заряда (инверсия потенциала). Как только количество ионов Na+ снаружи и внутри клетки сравняется, направленный ток в клетку /^прекращается и инверсия заканчивается при величине примерно 430-40 мВ (рис. I).
УШ - уровень потенциала покоя; МП - местный потенциал; КУД -критический уровень деполяризации мембраны; ПД - потенциал действия; КУД И - величина инверсии (овершут) СДП - следовой деполяризационный потенциал; СГП-следовой гиперполяризационный потенциал. СГП К этому моменту резко увеличивается проницаемость мембраны для ионов К+ которые в большом количестве выходят из клетки. В результате у внутренней поверхности мембраны вновь создается отрицательный заряд, а на наружной поверхности - положительный, т.е. происходит реполяризация мембраны. Быстрые изменения величины и полярности заряда мембраны получили название пика потенциала действия. Вслед за пиком потенциала действия наблюдаются деполяризационный и гиперполяризационный следовые потенциалы, обусловленные инерционностью процессов движения ионов Na+ и К+ через клеточную мембрану. Длительность потенциала действия составляет около I мс в нервах, 10 мс в скелетной мышце и более 200 мс в миокарде сердца. Поддержание разности концентраций ионов Na+ и К+ между цитоплазмой клетки и внеклеточной жидкости в состоянии покоя и восстановление этой разницы после раздражения клетки обеспечивается работой так называемого натрий-калиевого насоса мембраны. Натрий-калиевый насос осуществляет активный перенос ионов против градиентов их концентраций, непрерывно откачивая Na+ из клетки в обмен на К+. Насос работает за счет энергии АТФ, в расщеплении которой участвует фермент - мембранная Na, К АТФаза. Для работы насоса необходимо наличие в клетке ионов У Na+, а во внеклеточной жидкости - ионов К+. Распространение потенциала действия по ткани, в особенности нервного импульса по нервам, является самым быстрым и точно адресованным способом передачи информации в организме. Скорость передачи нервного импульса в быстропроводящих волокнах двигательных нервов (тип Аα) достигает 120 м/с. Другие способы передачи информации гораздо медлительнее: гуморальный не превышает 0,5 м/с (скорость тока крови в аорте), аксонный транспорт веществ от тела нейрона к окончаниям аксона не превышает 40 см в сутки.
Передача информации в организме путем проведения потенциалов действия осуществляется по мембране нервного волокна. При нанесении раздражения достаточной силы на нервное волокно в точке раздражения возникает зона возбуждения (рис. 2). Эта зона имеет на внутренней поверхности мембраны положительный заряд, а на наружной -отрицательный. Соседние невозбужденные участки мембраны нервного волокна имеют обратное соотношение полярности зарядов. Между возбужденным и невозбужденными участками мембраны возникают электрические токи. Они получили название местных токов. Эти токи раздражают соседние невозбужденные участки мембраны. В результате в них изменяется проницаемость ионных каналов, развивается деполяризация и возникает потенциал действия. Эти участки становятся возбужденными. Процесс повторяется и таким образом происходит распространение нервного импульса по нерву в обе стороны от первоначального места нанесения раздражения. Таков механизм проведения возбуждения по безмякотному нервному волокну, в котором оно проводится с небольшой скоростью, постепенно ослабевая. В мякотных нервных волокнах потенциалы действия возникают только в перехватах Ранвье, где нет миелиновой оболочки, являющейся электрическим изолятором. В результате возбуждение в мякотном нервном волокне передается скачками, от одного перехвата Ранвье к другому. Скорость передачи возбуждения в нем выше, чем в безмякотном волокне, и передается оно практически без ослабления.
Понятие о рецепторах, органах чувств, анализаторах и сенсорных системах. Отделы анализаторов. Общие свойства анализаторов. Значение анализаторов для восприятия явлений внешней и внутренней среды. Организм человека и животных может нормально функционировать только при постоянном получении информации о состоянии и изменениях внешней среды, в которой он находится, а также о состоянии внутренней среды, всех частей тела. Без информации, поступающей в мозг, не могут осуществляться простые и сложные рефлексы вплоть до психической деятельности человека. Сложные акты поведения человека во внешней среде требуют постоянного анализа внешней ситуации, а также осведомленности нервных центров о состоянии внутренних органов. Специальные структуры нервной системы, обеспечивающие вход информации в мозг и анализ этой информации, И.П. Павлов назвал анализаторами. С помощью анализаторов осуществляется познание окружающего мира. При раздражении рецепторов в коре больших полушарий возникают ощущения, которые отражают отдельные свойства предметов и явлений. На основе ощущений формируются понятия и представления, отражающие взаимосвязи и зависимости между этими предметами и явлениями, делаются выводы и заключения, осуществляются адекватное поведение во внешней среде и практическая деятельность человека. Анализаторы при нормальном функционировании в пределах чувствительности своих рецепторов дают верное представление об окружающей действительности, что подтверждается практикой. Это дает возможность человеку познавать окружающий мир, достигать прогресса в областях знания, науки и техники. Информация, поступающая от различных рецепторов в центральную нервную систему, необходима для поддержания деятельного состояния ЦНС и всего организма в целом. Искусственное выключение большинства органов чувств в специальных экспериментах на животных приводило к резкому снижению тонуса коры и сонному состоянию животного. Разбудить его можно было только путем воздействия на невыключенные органы чувств. Специальные эксперименты на людях, помещенных в камеры, исключающие поступление зрительных, слуховых и других раздражений, показали, что резкое снижение поступления сенсорной информации отрицательно сказывается на способности концентрировать внимание, логически мыслить, выполнять умственные задачи. В ряде случаев появлялись зрительные и слуховые галлюцинации. Информация, передаваемая в ЦНС от рецепторов интероцептивного анализатора, расположенных во внутренних органах, служит основой процессов саморегуляции. Так, например, если изменяется давление крови, то в барорецепторах стенок сосудов возникает возбуждение. Оно передается в сосудодвигательный центр продолговатого мозга, импульсы от которого вызывают расширение сосудов и восстановление кровяного давления до нормальной величины. Помимо первичного сбора информации об окружающей среде и внутреннем состоянии организма важной функцией анализаторов является информирование нервных центров о результатах рефлекторной деятельности, т.е. осуществление обратных связей. Например, для точного выполнения ответной двигательной реакции на какое-либо раздражение ЦНС должна получать информацию от двигательного и вестибулярного анализаторов о силе и длительности выполняемых сокращений мышц, о скорости и точности перемещения тела, положении тела в пространстве, об изменениях темпа движений и т.д. Без этой информации невозможно формирование и совершенствование двигательных навыков, в том числе трудовых и спортивных. Восприятие любой информации о внешней и внутренней среде начинается с раздражения рецепторов. Рецептор - это нервное окончание или специализированная клетка, которая способна воспринимать раздражение и преобразовывать энергию раздражения в нервный импульс. Рецепторы подразделяют на экстерорецепторы, воспринимающие раздражения из внешней среды, и интерорецепторы, сигнализирующие о состоянии внутренних органов. Разновидностью интерорецепторов являются проприорецепторы, информирующие о состоянии и деятельности опорно-двигательного аппарата. В зависимости от характера раздражителей, к которым рецептор обладает избирательной чувствительностью, рецепторы подразделяют на несколько групп: механорецепторы. терморецепторы, фоторецепторы, хеморецепторы, электрорецепторы, болевые рецепторы. Трансформирование энергии раздражителя в процесс возбуждения, или нервный импульс, происходит за счет обмена веществ самих рецепторов. Раздражитель, действуя на рецептор, вызывает деполяризацию его мембраны и возникновение рецепторного, или генераторного потенциала, который сходен по своим свойствам с местным потенциалом. Когда рецепторный потенциал достигает величины критического потенциала, он вызывает возникновение афферентного импульса в нервном волокне, идущем от рецептора. Более широким понятием, чем рецептор, является понятие орган чувств, под которым понимают образование, включающее рецепторы, а также другие клетки и ткани, способствующие лучшему восприятию рецепторами какого-то определенного раздражения. Например, рецепторы зрения (фоторецепторы) - это палочки и колбочки сетчатки глаза. Вместе с преломляющей системой, оболочками, мышцами, кровеносными сосудами глазного яблока фоторецепторы составляют орган чувств - глаз. Однако для возникновения ощущения одного органа чувств недостаточно. Необходимо, чтобы возбуждение от органа чувств было передано по афферентным путям в ЦНС в соответствующие проекционные зоны в коре больших полушарий. Это было установлено русским ученым И.П. Павловым, который ввел в физиологию понятие анализатор, объединяющее все анатомические образования, в результате деятельности которых возникает ощущение. Анализатор состоит из периферического отдела (соответствующего органа чувств), проводникового отдела (афферентных проводящих путей) и коркового, или центрального, отдела (определенного участка в коре больших полушарий). Например, периферический отдел зрительного анализатора представлен глазом, проводниковый отдел - это зрительный нерв, корковый отдел - зрительная зона коры больших полушарий. Следует отметить, что в настоящее время в термин орган чувств часто вкладывают то же понятие, что и в анализатор. Дальнейшее изучение механизмов восприятия и анализа информации, а также реакции на нее организма привело к появлению более общего, чем анализатор, понятия сенсорные системы. Сенсорная система включает в себя не только сложную многоуровневую систему передачи информации от рецепторов к коре больших полушарий и анализа ее, что И.П. Павлов назвал анализатором, но и включает процессы синтеза различной информации в коре и регулирующие влияния коры к нижележащим нервным центрам и рецепторам. Сенсорные системы имеют сложную структуру. Возбуждение от рецепторов проводится в кору больших полушарий по так называемым специфическому и неспецифическому путям. Специфический путь включает в себя: I)рецептор; 2)первый чувствительный нейрон, расположенный всегда вне ЦНС в спинномозговых ганглиях или в ганглиях черепномозговых нервов; 3)второй нейрон, расположенный в спинном или продолговатом, или среднем мозге; 4)третий нейрон, находящийся в зрительных буграх промежуточного мозга; 5)четвертый нейрон, расположенный в проекционной зоне данного анализатора в коре больших полушарий. Со вторых нейронов специфического пути, т.е. в спинном, продолговатом и среднем мозге происходит также передача сенсорного возбуждения на пути в другие отделы головного мозга, в том числе в ретикулярную формацию. Из ретикулярной формации возбуждение может направляться по так называемым неспецифическим путям во все отделы коры больших полушарий. Анализаторам характерны следующие общие свойства. 1) Высокая чувствительность к адекватным раздражителям. Например, в ясную темную ночь человеческий глаз может различить свет свечи на расстоянии до 20 км. 2) Адаптация анализаторов, т.е. свойство приспосабливаться к постоянной интенсивности длительно действующего раздражителя. При действии сильного раздражителя возбудимость анализатора уменьшается и пороги раздражения возрастают, при действии слабого раздражителя возбудимость анализатора увеличивается и пороги раздражения снижаются. Не все анализаторы обладают одинаковой способностью к адаптации. Хорошо адаптируются обонятельный, температурный, тактильный анализаторы, очень мало адаптируются вестибулярный, двигательный и болевой анализаторы. Скорость и степень адаптации у разных анализаторов к разным раздражителям тоже различна. Например, темновая адаптация при переходе от яркого света к темноте развивается в течение часа, а световая адаптация при переходе от темноты к свету наступает в течение минуты. Физиологическое значение адаптации заключается в установлении оптимального количества сигналов, поступающих в ЦНС, и ограничении поступления импульсов, не несущих новую информацию. 3) Иррадиация и индукция в нейронах анализатора. Иррадиация - это распространение возбуждения на другие нейроны в корковом отделе того же анализатора. Ее можно наблюдать при рассматривании квадратов одинакового размера на разном фоне. Так, белый квадрат на черном фоне кажется больше, чем таких же размеров черный квадрат на белом фоне. Индукция бывает одновременная и последовательная. Одновременная индукция является процессом, противоположным иррадиации. Суть ее в том, что одновременно с развитием возбуждения в одних нейронах анализатора в соседних вызывается торможение. Последовательная индукция состоит в том, что после прекращения возбуждения в нервных центрах анализатора развивается процесс торможения, а после прекращения торможения - процесс возбуждения. Процессы одновременной и последовательной индукции лежат в основе явлений контраста. Например, кислое после сладкого кажется еще более кислым; теплая вода после холодной кажется горячей и т.д. 4) Следовые процессы в анализаторах. После прекращения раздражения рецепторов физиологические процессы в анализаторе еще длятся некоторое время в виде положительных и отрицательных следовых явлений. Положительные следовые процессы являются как бы кратковременным продолжением процессов, происходивших в анализаторах при действии раздражителя. Т.е. ощущение (зрительное, слуховое, вкусовое и т.д.) длится еще некоторое время после прекращения действия раздражителя на рецепторы. Благодаря положительным следовым явлениям возможно слитное восприятие раздельных кадров в кинофильме. 5) Взаимодействие анализаторов. Все анализаторы функционируют не изолированно, а во взаимодействии друг с другом. Их взаимодействие может усиливать или наоборот ослаблять ощущения. Па-пример, звуковые раздражители воспринимаются легче при сочетании их со световыми, на чем основана светомузыка..
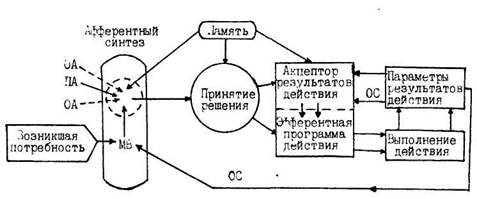
Системный принцип управления физиологическими функциями как основа сложного поведения. Понятие о функциональной системе поведенческого акта (П.К.Анохин). Составные элемент функциональной системы. Организм - это самостоятельно существующая единица органического мира. Он представляет собой саморегулирующуюся систему, реагирующую как единое целое на различные изменения внешней среды. В организме частные физиологические процессы подчинены закономерностям работы сложной целостной системы. Например, изменение обмена веществ и функций любой клетки, ткани, органа и системы органов вызывает изменения обмена веществ других клеток, тканей, органов и систем органов. Поэтому обмен веществ и функции клеток, тканей и органов, изолированных из организма, отличаются от происходящих внутри его. Целостному организму свойственны функции, такие как размножение, поведение, мышление, отсутствующие в изолированных его частях. Характерной для всякого организма является определенная организация его структур. Различают молекулярный уровень организации (вирусы), клеточный (простейшие), тканевой, органный и системный. Каждый орган или система органов выполняет специфическую функцию. Однако самостоятельность системы или органа является относительной. Живой организм представляет собой систему систем, которая в процессе взаимодействия с внешней средой обеспечивает получение полезного приспособительного результата. Так, при реализации поведенческой реакции, связанной с удовлетворением потребности животного в пище, различные физиологические реакции подчиняются решению главной задачи - получению пищи. Потребности живого организма могут быть удовлетворены только в результате активного взаимодействия его с внешней средой. Управление процессами жизнедеятельности в организме строится по принципу системной иерархичности, т.е. элементарные процессы подчинены более сложным. Ведущее значение в физиологических механизмах сложных поведенческих актов принадлежит нервной системе. ЦНС регулирует и координирует физиологические функции, определяя их ритм и общую направленность. В свою очередь, частные формы физиологических функций благодаря обратным связям оказывают влияние на высший управляющий аппарат. Такая форма контроля и взаимного влияния физиологических функций лежит в основе системного управления в целостном организме. П.К. Анохин первый обратил внимание на то, что системы в живом организме не просто анатомически соединяют входящие в них отдельные элементы, но и объединяют их для осуществления отдельных жизненно важных функций организма. Осуществление любого психического или физиологического процесса связано с образованием в организме функциональных систем, обеспечивающих достижение нужных результатов и обусловливающих целенаправленное поведение. Анохин понимал временное саморегулирующееся объединение рецепторов, различных мозговых структур и исполнительных органов, взаимодействующих совместно с целью достижения приспособительных полезных для организма результатов. В отличие от традиционных анатомо-физиологических систем, которые состоят из определенного постоянного набора органов, функциональные системы производят избирательное объединение различных органов в разных комбинациях из разных анатомических систем для достижения полезных для организма приспособительных результатов. Один и тот же орган, включенный в различные функциональные системы, может выполнять различные функции. Функциональная система целостного поведенческого акта (рис.1) включает в себя следующие механизмы: I) афферентный синтез; 2) принятие решения; 3) акцептор результатов действия; 4) выполнение действия; 5) получение результатов действия и сравнение их на основе обратной афферентации с программой действия. Стадия афферентного синтеза складывается из мотивационного возбуждения, обстановочной афферентации, использования аппарата памяти, пусковой афферентации. Работа функциональной системы направлена на получение полезного приспособительного результата для удовлетворения возникшей биологической или социальной потребности. Вызвав активность в определенных структурах мозга, потребность приводит к возникновению мотивации. В организм постоянно поступает много разнообразной информации и одновременно может существовать несколько мотиваций. В каждый момент времени мотивация, в основе которой лежит наиболее важная потребность, становится доминирующей. Доминирующее мотивационное возбуждение определяет все последующие этапы мозговой деятельности по формированию поведенческих программ. Для правильного программирования дальнейшего поведения организму необходимо оценить окружающую обстановку и свое положение в ней. Это достигается благодаря обстановочной афферентации, т.е. поступлению от рецепторов потока импульсов, несущих информацию об условиях, в которых предполагается осуществить поведенческий акт, направленный на удовлетворение возникшей потребности. Обязательным компонентом, который неоднократно используется в функциональной системе, является нейрофизиологический аппарат памяти. Благодаря памяти, обстановочная афферентация сравнивается с теми условиями в прошлом, при которых была успешной деятельность, которую организму предстоит совершить.
Рис. I. Упрощенная схема поведенческого акта с основными механизмами функциональной системы: ОА - обстановочная афферентация; ПA - пусковая афферентация; MB - мотивационное возбуждение; ОС - обратные связи. Если окружающая обстановка и состояние организма благоприятствуют предполагаемому поведенческому акту, то информация, поступающая от рецепторов, становится пусковой (пусковая афферентация) для принятия решения о выполнении действий для удовлетворения потребности. На основе афферентного синтеза осуществляется принятие решения. Извлекая из памяти информацию о собственном или чужом опыте по удовлетворению потребности в аналогичной обстановке, мозг выбирает один из многих способов для достижения поставленной цели. При этом избирательно возбуждаются нервные центры, которые обеспечивают осуществление выбранной поведенческой реакции. Деятельность нервных структур, мешающих выполнению данной реакции, тормозится. Вслед за принятием решения вырабатывается эфферентная программа действия и одновременно формируется специальный аппарат прогнозирования будущих результатов - акцептор результатов действий. Эфферентная программа действия представляет собой определенную последовательность набора нервных команд, поступающих на исполнительные органы - эффекторы. В каждом конкретном случае это могут быть различные комбинации органов из разных анатомических систем организма. Но они объединяются нервными и эндокринными влияниями и некоторое время функционируют взаимозависимо и совместно для достижения полезного приспособительного результата. Нередко различные функциональные системы для достижения различных приспособительных результатов могут использовать одни и те же органы. Например, сердце является необходимым компонентом и в функциональной системе для поддержания постоянного уровня кровяного давления, и в функциональных системах по обеспечению газообмена, терморегуляции и др. Акцептор результатов действия представляет собой нейронную модель предполагаемого результата, к которому должно привести данное действие. Предвидение будущих результатов происходит благодаря последовательному возбуждению корково-подкорковых структур мозга, которое опережает реальные события и возникает еще до поступления афферентных сигналов от рабочего органа (обратных связей) о выполнении действия. Информация о последовательности возбуждения нервных центров хранится, вероятно, в долговременной памяти. Благодаря акцептору результатов действия осуществляется быстрое включение согласно с программой исполнительных органов функциональной системы и происходит выполнение действия. Осуществление действия приводит к реальному результату, информация о котором с помощью обратной афферентации (обратных связей) поступает в акцептор действия, где сравнивается с запрограммированным результатом. Если полученный эффект соответствует запрограммированному, то человек испытывает положительные эмоции. Программа, приводящая к успешному выполнению поведенческого акта и полезному приспособительному результату, закрепляется в долгосрочной памяти, а сформировавшаяся функциональная система перестает существовать, т.к. произошло удовлетворение потребности и соответствующая мотивация перестает быть доминирующей. При отсутствии ожидаемого результата возникают отрицательные эмоции и может произойти один из вариантов: I) повторная попытка выполнения тех же рефлекторных реакций по той же программе;- 2) при стойкой мотивации происходит перестройка программы действия, вносятся поправки в его выполнение; 3) при нестойкой мотивации отсутствие ожидаемого результата может привести к изменению самой мотивации или к ее исчезновению. Таким образок, сложные поведенческие акты организма строятся не по типу раздражение рецептора - ответная реакция эффектора, а по принципу рефлекторных кольцевых взаимодействий, которые является одним из основных механизмов деятельности функциональных систем. Можно привести следующий пример образования и деятельности функциональной системы в организации поведения в повседневной жизни. Приближение праздника 8-е Марта вызывает социальную потребность у подростка поздравить свою мать, в результате чего возникает доминирующее мотивационное возбуждение. Сын задумывается на тем, какой подарок сделать маме и вспоминает, что ей нравятся цветы гладиолусы, роман М. Митчел "Унесенные ветром", повести В.Быкова и французские духи. Обстановочная афферентация показывает, что в начале марта цветущих гладиолусов не найти, а духи дорогие и на них у подростка не хватает денег. Доступность книг по цене делает эту афферентную информацию пусковой. Принимается решение - купить какую-нибудь из нравящихся маме книг, желательно роман "Унесенные ветром" т.к. она давно уже хотела его иметь. Школьник вспоминает, что нужную книгу он недавно видел в двух магазинах. Составляется программа выполнения - посмотреть и купить роман в ближайшем книжном магазине. Однако в магазинах подросток узнает, что необходимый роман уже раскупили. Эта информация является отрицательной обратной связью. Она поступает в акцептор результатов действия. Поскольку.полученный результат (роман не куплен) не совпадает с запрограммированным, акцептор результатов действия вносит поправку в программу действий: съездить еще на книжный рынок и если не будет романа "Унесенные ветром", то купить книгу повестей В.Быкова. На книжном рынке подросток находит повести В.Быкова и покупает их. Полезный результат достигнут. Потребность школьника удовлетворяется, мотивация угасает и данная функциональная система перестает существовать.
Понятие об адаптации. Стресс. Учение об общем адаптационном синдроме. Роль систем гипоталамус -гипофиз - надпочечники в адаптации организма к нагрузкам. В общебиологическом смысле адаптация - это совокупность врожденных и приобретенных анатомо-морфологических, физиологических, поведенческих и других особенностей организма, обеспечивающая его приспособление к условиям внешней среды и создающая возможность специфического образа жизнедеятельности. Адаптация поддерживает гомеостаз и возникает в результате процессов, протекающих на молекулярном, клеточном, органном, системном и организменном уровнях. Ч. Дарвин показал, что адаптационные приспособления закрепляются в результате действия естественного отбора. В результате длительной эволюции и онтогенеза организмы адаптированы к своим адекватным условиям обитания. Например, рыбы приспособлены к жизни в воде, птицы - к полету и т.д. Приспособление к периодическому колебанию таких адекватных условий происходит в основном с помощью готовых специфических адаптивных механизмов. Различают общие адаптации и частные адаптации (специализации). К одним факторам среды организмы могут достигать полной, к другим - только частичной адаптации. На первом этапе адаптации к колебаниям адекватных условий среды активируется условнорефлекторная деятельность организма. В дальнейшем, несмотря на повторные воздействия раздражителей, в процессе адаптации происходит угасание ориентировочной реакции и "привыкание" к действию раздражителя, В этом случае термин "адаптация" применяется в более узком смысле и обозначает снижение чувствительности рецепторов, а также приспособление центрального отдела соответствующего анализатора к постоянно действующему адекватному раздражителю. Адаптация рецепторов отличается от их утомления тем, что она быстро возникает после начала раздражения. Когда же действие раздражителя прекращается, то адаптация довольно быстро исчезает и чувствительность рецепторов повышается. При выраженных изменениях окружающей среды возникают неадекватные условия для жизнедеятельности организма. Это включает в действие неспецифические адаптивные механизмы. В 1936 году канадский ученый Г. Селье в опытах на животных установил, что при действии на организм сильных и продолжительных раздражителей возникает комплекс неспецифических защитных реакций. Г. Селье назвал этот комплекс общим адаптационным синдромом. Состояние организма в период воздействия вредных факторов он назвал стрессом (от англ. стресс - напряжение), а факторы, вызывающие состояние стресса - стрессорами. Каждый стрессор вызывает в организме характерные изменения. Так, например, вирус гриппа приводит к специфическому заболеванию - гриппу, но наряду со специфическими изменениями в организме каждый стрессор вызывает ряд неспецифических, присущих всем видам стресса, стереотипных ответных реакций. Этот комплекс реакций, направленных на мобилизацию защитных сил организма, на сохранение его жизни, представляет собой общий адаптационный синдром. Он является механизмом общей адаптации организма. В результате общего адаптационного синдрома обеспечивается: I) мобилизация энергетических ресурсов организма и энергетическое обеспечение функций; 2) мобилизация пластического резерва организма и синтез ферментов и белков, необходимых для защиты организма от стрессора; 3) мобилизация защитных способностей организма. Важной стороной механизма общей адаптации является то, что в результате приспособительного синтеза белков, происходит переход в долговременную адаптацию, в основе которой лежит изменение и совершенствование клеточных структур. Примером перехода краткосрочных адаптационных реакций в долговременную адаптацию является физическая тренировка, которая сопровождается повышением функциональных возможностей организма. Развитие общего адаптационного синдрома невозможно без участия гипофиза и коры надпочечников. При их удалении у животных этот синдром не развивается, и они быстро гибнут под воздействием стрессора. Г. Селье выделил три стадии в развитии общего адаптационного синдрома: стадию тревоги, стадию резистентности (устойчивости), стадию истощения. Стадия тревоги начинается с момента начала действия на организм сильного раздражителя - стрессора. Стрессор вызывает повышение функциональной активности гипоталамуса, которое может осуществляться разными способами. Во-первых, рефлекторным путем. т.к. многие стрессовые раздражители, воздействуя на экстерорецепторы и интерорецепторы, вызывают поток импульсов от них к гипоталамусу. Во-вторых, большинство стрессоров вызывают возбуждение симпатического отдела нервной системы и усиление секреции адреналина мозговым веществом надпочечников. Адреналин, поступая с кровью в гипоталамус, значительно усиливает его активность. В-третьих, активирование гипоталамуса может быть вызвано также гуморальным путем в результате непосредственного воздействия продуктов обмена веществ и распада тканей, которые могут появиться в циркулирующей крови под действием сильного стрессора. В-четвертых, повышение функции гипоталамуса может возникнуть в результате воздействия импульсов, поступающих из коры больших полушарий при психическом стрессе. Повышение функциональной активности гипоталамуса приводит к увеличению выработки в нем кортиколиберина, который поступает в переднюю долю гипофиза и там способствует повышению образования адренокортикотропного гормона (АКТГ). АКТГ с током крови поступает в кору надпочечников и вызывает усиление секреции глюкокортикоидов. Глюкокортикоиды обладают противовоспалительным и противоаллергическим действием, активируют синтез многих ферментов, повышают проницаемость клеточных мембран для воды и ионов, повышают возбудимость ЦНС. Глюкокортикоиды оказывают сильное воздействие на обмен белков, жиров и углеводов. Они способствуют распаду белков до аминокислот, что увеличивает количество исходного "строительного" материала для синтеза других белков и ферментов, необходимых в условиях стресса. Кроме того, под действием глюкокортикоидов в печени происходит образование углеводов из остатков аминокислот. Глюкокортикоиды усиливают мобилизацию жира из жировых депо и использование его в процессах энергетического обмена. Под влиянием глюкокортикоидов увеличивается запасы гликогена в печени и концентрация глюкозы в крови. В результате такого многостороннего влияния глюкокортикоидов на обмен веществ улучшается энергетическое обеспечение физиологических функций и повышается устойчивость организма к стрессовым факторам. Вторая стадия - стадия резистентности (устойчивости) характеризуется увеличением массы передней доли гипофиза и надпочечников, повышенной секрецией АКТГ и глюкокортикоидов. Увеличенное содержание глюкокортикоидов в крови повышает устойчивость организма к действию стрессора и общее состояние организма нормализуется, т.е. организм адаптируется к действию стрессора. Однако всякое приспособление имеет свои границы. При длительном или слишком частом повторении воздействия сильного стрессора или при одновременном действии на организм нескольких стрессоров стадия резистентности переходит в третью стадию - стадию истощения. В эту стадию кора надпочечников не в состоянии вырабатывать еще большее количество глюкокортикоидов, названных Г. Селье адаптивными гормонами. Поэтому защитные силы организма и его сопротивление уже не могут полностью противостоять действию стрессоров. Состояние организма ухудшается, может наступить его заболевание и смерть. Глюкокортикоиды играют также важную роль в адаптации организма к мышечным нагрузкам. При увеличении физической работы повышается активность коры надпочечников и содержание глюкокортикоидов в крови увеличивается. Это приводит к мобилизации энергетических ресурсов организма и он способен достаточно долго, без ущерба для себя выполнять данную физическую или психическую нагрузку. Однако при длительных утомительных нагрузках вслед за первоначальным усилением происходит уменьшение выработки глюкокортикоидов. Энергетическое обеспечение работы становится недостаточным и организм снижает ее интенсивность или совсем прекращает. В противном случае может наступить переутомление и истощение организма, что может стать причиной заболеваний.
Понятие о железах внутренней секреции. Гуморальная регуляция функций. Факторы гуморальной регуляции. Понятие о гормонах и их свойствах. Взаимосвязь неравной и гуморальной регуляции функций. Различают два основных механизма регуляции функций - нервный и гуморальный, которые взаимосвязаны и образуют единую нейрогуморальную регуляцию. Гуморальный (от латинского humor - жидкость), или химический механизм регуляции является филогенетически более древним. Он осуществляется за счет химических веществ, находящихся в циркулирующих в организме жидкостях, т.е. в крови, лимфе и тканевой жидкости. Факторами гуморальной регуляции функций могут быть: 1) физиологически активные вещества - гормоны, вырабатываемые эндокринными железами и некоторыми другими органами и клетками организма (например, гормон адреналин вырабатывается эндокринной железой - мозговым веществом надпочечников, а также хромафинными клетками, находящимися в нервных узлах, стенке кровеносных сосудов и др. органах); 2) некоторые специфические продукты обмена веществ клеток, в том числе и медиатора (ацетилхолин, норадреналин и др.); 3) некоторые неспецифические продукты обмена веществ клеток (например, СО2 оказывает возбуждающее действие на клетки дыхательного центра продолговатого мозга);4) некоторые вещества, поступающие вместе с продуктами питания, при дыхании, через кожу (например, никотин, вдыхаемый с табачным дымом, снижает возбудимость нервных клеток и оказывает отрицательное воздействие на деятельность многих клеток и тканей). Важнейшим видом гуморальной регуляции функций является гормональная регуляция, осуществляемая посредством гормонов, которые вырабатываются эндокринными железами. Кроме того, гормоноподобные вещества выделяются и некоторыми другими органами и клетками организма, выполняющими, помимо эндокринной, другую специализированную функцию (почки, плацента, клетки слизистой оболочки пищеварительного тракта и др.). Эти вещества получили название тканевых гормонов. Эндокринные железы (от греч. endon. - внутри, crino - выделяю) нt имеют выводных протоков и выделяют гормоны во внутреннюю среду организма, вследствие чего они получили второе название - железы внутренней секреции. К эндокринным железам человека и высших животных относятся: гипофиз (передняя, промежуточная и задняя доли), щитовидная железа, паращитовидные железы, надпочечник (мозговое и корковое вещество), поджелудочная железа, половые железы (яичники и семенники), эпифиз, вилочковая железа. Половые железы и поджелудочная железа осуществляют наряду с внутрисекреторной и внешне-секреторную функцию, т.е. являются железами смешанной секреции. Так, половые железы вырабатывают не только половые гормоны, но и половые клетки - яйцеклетки и сперматозоиды, а часть клеток поджелудочной железы вырабатывает поджелудочный сок, который выделяется по протоку в 12-перстную кишку, где участвует в пищеварении. Эндокринные железы осуществляют гуморальную регуляцию посредством вырабатываемых ими гормонов. Термин гормон (от греч. hormao) - привожу в движение, возбуждаю) был введен В. Бейлисом и E. Старлингом. По химическому строению гормоны высших животных и человека можно разделить на три основные группы: I) белки и пептиды; 2) производные аминокислот; 3) стероиды. Биосинтез гормонов запрограммирован в генетическом аппарате специализированных эндокринных клеток. По своему функциональному действию гормоны подразделяются на эффекторные, которые оказывают влияние непосредственно на орган-мишень, и тропные, основной функцией которых является регуляция синтеза и выделения эффекторных гормонов. Кроме того, нейронами гипоталамуса вырабатываются нейрогормоны, одни из которых - либерины стимулируют секрецию гормонов передней доли гипофиза, а другие тормозят этот процесс - статины. Гормоны оказывают большое регулирующее влияние на различные функции организма. Выделяют 3 основные функции гормонов: I) регуляция обмена веществ, в результате которой обеспечивается адаптация организма к условиям существования и поддерживается гомеостаз; 2) обеспечение развития организма, т.к. гормоны влияют на размножение организма, рост и дифференцировку клеток и тканей; 3) Kоррекция физиологических процессов в организме, т.е. гормоны могут вызвать, усилить или ослабить работу каких-то органов и осуществление физиологических реакций, что также обеспечивает адаптацию и гомеостаз организма. Действие гормонов на клетки-мишени осуществляется путем влияния на активность ферментов, на проницаемость клеточных мембран и на генетический аппарат клетки. Механизм действия стероидных гормонов отличается от механизма действия гормонов белково-пептидной и аминокислотной групп. Гормоны белково-пептидной и аминокислотной групп не проникают внутрь клетки, а присоединяются на ее поверхности к специфическим рецепторам клеточной мембраны. Рецептор связывает фермент аденилатциклазу и она находится в неактивной форме. Гормон, действуя на рецептор, активирует аденилатциклазу, которая расщепляет АТФ с образованием циклического аденозинмонофосфата (цАМФ). Включаясь в сложную цепь реакций, цАМФ вызывает активацию определенных ферментов, что и обуславливает конечный эффект действия гормона. Стероидные гормоны имеют относительно небольшие размеры молекул и могут проникать через клеточную мембрану. В цитоплазме гормон взаимодействует со специфическим веществом, являющимся для него рецептором. Гормон-рецепторный комплекс транспортируется в ядро клетки, где обратимо взаимодействует с ДНК. В результате этого взаимодействия активируются определённые гены, на которых образуется информационная РНК. Информационная РНК поступает в рибосому, где происходит синтез фермента. Образовавшийся фермент катализирует определённые биохимические реакции, что влияет на физиологические функции клеток, тканей и органов. В связи с тем что стероидные гормоны не активируют готовые ферменты, а вызывают синтез новых молекул, действие стероидных гормонов проявляется медленнее, но длится дольше, чем влияние белков белково-пептидной и аминокислотной групп. Гормоны обладают рядом характерных свойств: 1. Высокая биологическая активность. Это означает, что гормоны в очень малых концентрациях могут вызывать значительные изменения физиологических функций. Так, 1 г адреналина достаточно, чтобы усилить работу изолированных сердец 10 миллионов лягушек, г инсулина достаточно, чтобы понизить уровень сахара у 125000 кроликов. Гормоны транспортируются кровью не толи о в свободном, но и в связанном виде с белками плазмы крови или ее форменными элементами. Поэтому активность действия гормона в этом случае зависит не только от концентрации его в крови, но и от скорости отщепления его от транспортирующих белков и форменных элементов. 2. Специфичность действия. Каждый гормон имеет свою определённую химическую структуру. Поэтому в организме гормон хотя и достигает с током крови всех органов и тканей, но действует только на те клетки, ткани и органы, которые обладают специфическими рецепторами, способными взаимодействовать с гормоном. Такие клетки, ткани и органы получили название клеток-мишеней, тканей-мишеней и органов-мишеней. 3.Дистантность действия. Гормоны за исключением тканевых гормонов, переносятся кровью далеко от места их образования и оказывают действие на отдаленные органы и ткани. 4. Гормоны стероидной группы и в меньшей степени гормоны щитовидной железы сравнительно легко проникают через мембраны клеток. 5. Гормоны сравнительно быстро разрушаются в тканях и особенно в печени. 6. Гормоны стероидной и аминокислотной групп не имеют видовой специфичности и поэтому возможно применение для лечения человека гормональных препаратов, полученных от животных. Интенсивность синтеза и выделения гормона железой регулируется в соответствии с величиной потребности организма в данном гормоне. Как только изменения, вызываемые каким-либо гормоном, достигают оптимальной величины, образование и выделение этого гормона уменьшаются. Регуляция уровня выделения гормонов осуществляется несколькими способами: I) прямое влияние на клетки железы того вещества, уровень которого контролируется данным гормоном (например, при повышении концентрации глюкозы в крови, протекающей через поджелудочную железу, увеличивается секреция инсулина, снижающего уровень глюкозы); 2) гормоны, вырабатываемые одними железами, оказывают влияние на выделение гормонов другими железами (например, тиреотропный гормон гипофиза стимулирует секрецию гормонов щитовидной железой); 3) нервная регуляция образования гормонов осуществляется главным образом через гипоталамус путем изменения уровня секреции нейронами гипоталамуса либеринов и статинов, которые поступают в переднюю долю гипофиза и влияют там на выделение гормонов; 4) выработка гормонов клетками мозгового вещества надпочечников и эпифиза увеличивается при непосредственном поступлении к ним нервных импульсов. Нервные волокна, иннервирующие другие эндокринные железы регулируют в основном тонус кровеносных сосудов и кровоснабжение железы, тем самым влияя и на секрецию гормонов. Разные гормоны, вырабатываемые разными железами, могут взаимодействовать между собой. Это взаимодействие может выражаться в синергизме действия, антагонизме действия и в позволяющем действии гормонов. Примером синергического, или однонаправленного, влияния можно привести действие адреналина (гормон мозгового вещества надпочечников) и глюкагона (гормон поджелудочной железы), которые активируют распад гликогена печени до глюкозы и повышают уровень глюкозы в крови. Пример антагонизма действия гормонов: адреналин повышает уровень глюкозы в крови, а инсулин (гормон поджелудочной железы) снижает уровень глюкозы. Позволяющее действие гормонов выражается в том, что гормон, сам не влияющий на данный физиологический показатель, создает условие для лучшего действия какого-то другого гормона. Например, сами глюкокортикоиды (гормоны коры надпочечников) не влияют на тонус мускулатуры сосудов, но повышают их чувствительность к адреналину.
Временные связи и условные рефлексы. Значение условных рефлексов в приспособительной эволюции животного мира. Механизм образования условных рефлексов. Торможение условных рефлексов. И. П. Павлов показал, что все рефлекторные реакции можно разделить на две группы: безусловные и условные. Дата добавления: 2015-12-15 | Просмотры: 1492 | Нарушение авторских прав |
 Рис. I. Развитие потенциала действия в нерве:
Рис. I. Развитие потенциала действия в нерве: