|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
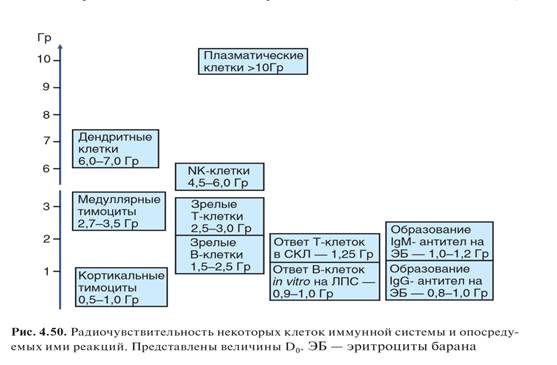
Иммунодефицитные состояния, обусловленные гибелью иммуноцитовКлассические примеры таких иммунодефицитов — нарушения иммунитета, вызванные действием ионизирующей радиации и цитотоксических лекарственных средств. Лимфоциты относят к немногочисленным клеткам, реагирующим на действие ряда факторов, в частности повреждающих ДНК, развитием апоптоза. Этот эффект проявляется при действии ионизирующей радиации и многих цитостатиков, используемых в лечении злокачественных опухолей (например, цисплатина, внедряющегося в двойную спираль ДНК). Причина развития апоптоза в этих случаях — накопление нерепарированных разрывов, регистрируемых клеткой с участием киназы АТМ, от которой сигнал поступает по нескольким направлениям, в том числе к белку р53. Этот белок отвечает за запуск апоптоза, биологический смысл которого состоит в защите многоклеточного организма ценой гибели единичных клеток, которые несут генетические нарушения, чреватые риском малигнизации клетки. В большинстве других клеток (как правило, покоящихся) срабатыванию этого механизма противодействует защита от апоптоза, обусловленная повышенной экспрессией белков Bcl-2 и Bcl-XL. Радиационные иммунодефициты Уже в первое десятилетие после открытия ионизирующих излучений была обнаружена их способность ослаблять резистентность к инфекционным заболеваниям и избирательно снижать содержание лимфоцитов в крови и лимфоидных органах. Радиационный иммунодефицит развивается сразу после облучения организма. Действие радиации обусловлено преимущественно двумя эффектами: нарушением естественных барьеров, прежде всего слизистых оболочек, что приводит к усилению доступа в организм патогенов; избирательным повреждением лимфоцитов, а также всех делящихся клеток, включая предшественники клеток иммунной системы и клетки, вовлекаемые в иммунный ответ. Предметом изучения радиационной иммунологии является, главным образом, второй эффект. Радиационная гибель клеток реализуется по двум механизмам — митотическому и интерфазному. Причина митотической гибели — нерепарируемые повреждения ДНК и хромосомного аппарата, препятствующие осуществлению митозов. Интерфазная гибель затрагивает покоящиеся клетки. Ее причиной служит развитие апоптоза по р53/АТМзависимому механизму Если чувствительность всех типов клеток к митозу примерно одинакова (D0 — около 1 Гр), то по чувствительности к интерфазной гибели лимфоциты значительно превосходят все остальные клетки: большинство их погибает при облучении в дозах 1–3 Гр, тогда как клетки других типов погибают при дозах, превышающих 10 Гр. Высокая радиочувствительность лимфоцитов обусловлена, как уже сказано, низким уровнем экспрессии антиапоптотических факторов Bcl-2 и Bcl-XL. Различные популяции и субпопуляции лимфоцитов несущественно различаются по чувствительности к апоптозу (В-клетки несколько чувствительнее Т-лимфоцитов; D0 для них составляет соответственно 1,7–2,2 и 2,5–3,0 Гр). В процессе лимфопоэза чувствительность к цитотоксическим воздействиям изменяется в соответствии с уровнем экспрессии в клетках антиапоптотических факторов: она наиболее высока в периоды селекции клеток (для Т-лифмоцитов — стадия кортикальных CD4+ CD8+ тимоцитов, D0 — 0,5–1,0 Гр). Радиочувствительность высока у покоящихся клеток, она дополнительно возрастает на начальных этапах активации, а затем резко снижается. Высокой радиочувствительностью характеризуется процесс пролиферативной экспансии лимфоцитов, причем при вступлении в пролиферацию могут погибнуть клетки, подвергшиеся действию излучения ранее и несущие нерепарированные разрывы ДНК. Сформировавшиеся эффекторные клетки, особенно плазматические, устойчивы к действию радиации (D0 — десятки Гр). В то же время клетки памяти радиочувствительны примерно в той же степени, что и наивные лимфоциты. Клетки врожденного иммунитета радиорезистентны. Радиочувствительны только периоды их пролиферации во время развития. Исключение составляют NK-клетки, а также дендритные клетки (погибают при дозах 6–7 Гр), которые по радиочувствительности занимают промежуточное положение между другими лимфоидными и миелоидными клетками. Хотя зрелые миелоидные клетки и опосредуемые ими реакции радиорезистентны, в ранние сроки после облучения максимально проявляется именно недостаточность миелоидных клеток, в первую очередь нейтрофилов, вызванная радиационным нарушением гемопоэза. Его последствия раньше и тяжелее всего сказываются на нейтрофильных гранулоцитах как популяции клеток с наиболее быстрым обменом пула зрелых клеток. Это обусловливает резкое ослабление первой линии защиты, нагрузка на которую именно в этот период значительно возрастает в связи с нарушением барьеров и бесконтрольным поступлением в организм патогенов и других чужеродных агентов. Ослабление этого звена иммунитета служит главной причиной радиационной гибели в ранние сроки после облучения. В более поздние сроки последствия поражения факторов врожденного иммунитета сказываются значительно слабее. Функциональные проявления врожденного иммунитета сами по себе устойчивы к действию ионизирующих излучений. Через 3–4 сут после облучения в дозах 4–6 Гр у мышей погибает более 90% лимфоидных клеток и происходит опустошение лимфоидных органов. Функциональная активность выживших клеток снижается. Резко нарушается хоминг лимфоцитов — их способность мигрировать в процессе рециркуляции во вторичные лимфоидные органы. Реакции адаптивного иммунитета при действии этих доз ослабляются в соответствии со степенью радиочувствительности клеток, которые опосредуют эти реакции. В наибольшей степени от действия радиации страдают те формы иммунного ответа, развитие которых нуждается во взаимодействиях радиочувствительных клеток. Поэтому клеточный иммунный ответ более радиорезистентен, чем гуморальный, а тимуснезависимое антителообразование более радиорезистентно, чем тимусзависимый гуморальный ответ. Дозы радиации в интервале 0,1–0,5 Гр не вызывают повреждения периферических лимфоцитов и нередко оказывают стимулирующее действие на иммунный ответ, обусловленный прямой способностью квантов излучения, генерирующих активные формы кислорода, активировать в лимфоцитах сигнальные пути. Иммуностимулирующее действие радиации, особенно в отношении IgE-ответа, закономерно проявляется при облучении после иммунизации. Полагают, что в этом случае стимулирующий эффект обусловлен относительно более высокой радиочувствительностью регуляторных Т-клеток, контролирующих эту форму иммунного ответа, по сравнению с эффекторными клетками. Стимулирующее действие радиации на клетки врожденного иммунитета проявляется даже при действии высоких доз, особенно в отношении способности клеток продуцировать цитокины (IL-1, TNF α и др.). Помимо прямого стимулирующего действия радиации на клетки, проявлению усиливающего эффекта способствует стимуляция этих клеток продуктами патогенов, поступающих в организм через поврежденные барьеры. Однако повышение активности клеток врожденного иммунитета под действием ионизирующей радиации не является адаптивным и не обеспечивает адекватной защиты. В связи с этим превалирует отрицательное действие облучения, проявляющееся в подавлении (при дозах, превышающих 1 Гр) адаптивного антигенспецифичекого иммунного ответа (рис. 4.50). Уже в период развивающегося опустошения лимфоидной ткани включаются восстановительные процессы. Восстановление происходит двумя основными путями. С одной стороны, активизируются процессы лимфопоэза за счет дифференцировки всех разновидностей лимфоцитов из кроветворных стволовых клеток. В случае Т-лимфопоэза к этому добавляется развитие Т-лимфоцитов из внутритимусных предшественников. При этом в определенной степени повторяется последовательность событий, свойственных Т-лимфопоэзу в эмбриональном периоде: сначала образуются γδТ-клетки, затем— αβТ-клетки. Процессу восстановления предшествует омоложение эпителиальных клеток тимуса, сопровождающееся повышением выработки ими пептидных гормонов. Численность тимоцитов быстро возрастает, достигая максимума к 15-м суткам, после чего происходит вторичная атрофия органа вследствие исчерпания популяции внутритимусных клеток-предшественников. Эта атрофия мало сказывается на численности периферических Т-лимфоцитов, поскольку к этому времени включается второй источник восстановления популяции лимфоцитов.
Этот источник — гомеостатическая пролиферация выживших зрелых лимфоцитов. Стимул к реализации этого механизма регенерации лимфоидных клеток — выработка IL-7, IL-15 и BAFF, служащих гомеостатическими цитокинами соответственно для Т-, NK- и В-клеток. Восстановление Т-лимфоцитов происходит наиболее медленно, поскольку для реализации гомеостатической пролиферации необходим контакт Т-лимфоцитов с дендритными клетками, экспресирующими молекулы MHC. Численность дендритных клеток и экспрессия на них молекул MHC (особенно класса II) после облучения снижены. Эти изменения можно трактовать как индуцированные радиацией изменениия микроокружения лимфоцитов — лимфоцитарных ниш. С этим связана задержка восстановления пула лимфоидных клеток, особенно существенная для CD4+ Т-клеток, которая реализуется в неполном объеме. Т-клетки, формирующиеся в процессе гомеостатической пролиферации, имеют фенотипические признаки клеток. Для них характерны пути рециркуляции, свойственные этим клеткам (миграция в барьерные ткани и нелимфоидные органы, ослабление миграции в Т-зоны вторичных лимфоидных органов). Именно поэтому численность Т-лимфоцитов в лимфоузлах практически не восстанавливается до нормы, в то время как в селезенке она восстанавливается полностью. Иммунный ответ, развивающийся в лимфатических узлах, также не достигает нормального уровня при его полной нормализации в селезенке. Таким образом, под влиянием ионизирующей радиации изменяется пространственная организация иммунной системы. Другое следствие конверсии фенотипа Т-лимфоцитов в процессе гомеостатической пролиферации — учащение аутоиммунных процессов вследствие повышения вероятности распознавания аутоантигенов при миграции в нелимфоидные органы, облегчения активации Т-клеток памяти и отставания регенерации регуляторных Т-клеток по сравнению с остальными субпопуляциями. Многие изменения в иммунной системе, индуцированные радиацией, напоминают следствия обычного старения; особенно наглядно это проявляется в тимусе, возрастное снижение активности которого ускоряется облучением. Варьирование дозы облучения, его мощности, применение фракционированного, местного, внутреннего облучения (инкорпорированных радионуклидов) придает определенную специфику иммунологическим нарушениям в пострадиационном периоде. Однако принципиальные основы радиационного поражения и пострадиационного восстановления во всех этих случаях не отличаются от рассмотренных выше. Особую практическую значимость действие умеренных и малых доз радиации приобрело в связи с радиационными катастрофами, особенно в Чернобыле. Сложно точно оценить эффекты малых доз радиации и дифференцировать влияние радиации от роли привходящих факторов (особенно таких, как стресс). В этом случае могут проявляться уже упоминавшееся стимулирующее действие радиации как часть эффекта гормезиса. Радиационную иммуностимуляцию нельзя рассматривать как положительное явление, поскольку оно, во-первых, не адаптивно, во-вторых сопряжено с разбалансировкой иммунных процессов. Пока затруднительно объективно оценить влияние на иммунную систему человека того незначительного повышения естественного фона радиации, которое наблюдается в местностях, прилегающих к зонам катастроф или связанных с особенностями производственной деятельности. В подобных случаях радиация становится одним из неблагоприятных факторов среды и ситуацию следует анализировать в контексте экологической медицины. Дата добавления: 2016-06-05 | Просмотры: 504 | Нарушение авторских прав |