|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Морфофункциональная организация спинного мозгаСпинной мозг характеризуется выраженным сегментарным строением, отражающим сегментарное строение тела позвоночных. От каждого спинномозгового сегмента отходят две пары вентральных и дорсальных корешков. Дорсальные корешки формируют афферентные входы спинного мозга. Они образованы центральными отростками волокон первичных афферентных нейронов, тела которых вынесены на периферию и находятся в спинномозговых ганглиях. Вентральные корешки образуют эфферентные выходы спинного мозга. В них проходят аксоны a и g-мотонейронов, а также преганглионарных нейронов вегетативной нервной системы. Такое распределение афферентных и эфферентных волокон было установлено еще в начале прошлого века и получило название закона Белла – Мажанди. После перерезки передних корешков на одной стороне наблюдается полное выключение двигательных реакций; но чувствительность этой стороны тела сохраняется. Перерезка задних корешков выключает чувствительность, но не приводит к утрате двигательных реакций мускулатуры.
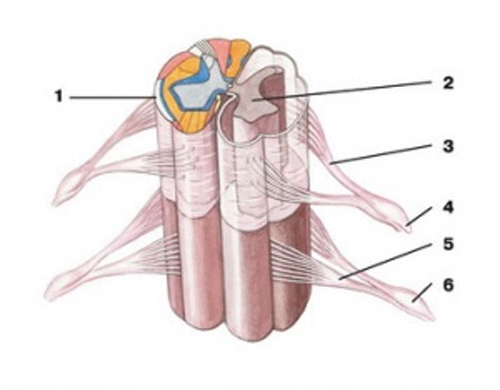
Объемная реконструкция спинного мозга: 1 — белое вещество; 2 — серое вещество; 3 — задний (чувствительный) корешок; 4 — спинно-мозговые нервы; 5 — передний (двигательный) корешок; 6 — спинно-мозговой ганглий Нейроны спинномозговых ганглиев относятся к простым униполярным, или псевдоуниполярным, нейронам. Название «псевдоуниполярный» объясняется тем, что в эмбриональном периоде первичные афферентные нейроны происходят от биполярных клеток, отростки которых затем сливаются. Нейроны спинномозговых ганглиев можно подразделить на клетки малых и больших размеров. Тело крупных нейронов имеет диаметр порядка 60–120 мкм, в то время как у мелких нейронов он колеблется от 14 до 30 мкм. Крупные нейроны дают начало толстым миелинизированным волокнам. От мелких начинаются как тонкие миелинизированные, так и немиелинизированные волокна. После бифуркации оба отростка направляются в противоположные направления: центральный входит в дорсальный корешок и в его составе – в спинной мозг, периферический – в различные соматические и висцеральные нервы, подходящие к рецепторным образованиям кожи, мышц и внутренних органов. Иногда центральные отростки первичных афферентных нейронов заходят в вентральной корешок. Это происходит при трифуркации аксона первичного афферентного нейрона, в результате которой его отростки проецируются в спинной мозг и через дорсальный и вентральный корешки. Из всей популяции клеток дорсальных ганглиев примерно 60–70% относится к мелким нейронам. Это соответствует тому, что число немиелинизированных волокон в дорсальном корешке превышает число миелинизированных волокон. Тела нейронов спинномозговых ганглиев не имеют дендритных отростков и не получают синоптических входов. Их возбуждение происходит в результате прихода потенциала действия по периферическому отростку, контактирующему с рецепторами. Клетки спинномозговых ганглиев содержат высокие концентрации глутаминовой кислоты – одного из предполагаемых медиаторов. Их поверхностная мембрана содержит рецепторы, специфически чувствительные к g-аминомасляной кислоте, что совпадает с высокой чувствительностью к g-аминомасляной кислоте центральных окончаний первичных афферентных волокон. Малые нейроны ганглиев содержат вещество Р или соматостатин. Оба этих полипептида также являются вероятными медиаторами, высвобождаемыми окончаниями первичных афферентных волокон. Каждая пара корешков соответствует одному из позвонков и покидает позвоночный канал через отверстие между ними. Поэтому сегменты спинного мозга принято обозначать по тому позвонку, возле которого из спинного мозга выходят соответствующие корешки. Спинной мозг принято также разделять на несколько отделов: шейный, грудной, поясничный и крестцовый, каждый из которых содержит по нескольку сегментов. В связи с развитием конечностей нейронный аппарат тех сегментов спинного мозга, которые их иннервируют, получил наибольшее развитие. Это нашло свое отражение в образовании шейного и поясничного утолщений. В области утолщений спинного мозга корешки содержат наибольшее количество волокон и имеют наибольшую толщину. На поперечном срезе спинного мозга ясно выделяется центрально расположенное серое вещество, образованное скоплением нервных клеток, и окаймляющее его белое вещество, образованное нервными волокнами. В сером веществе различают вентральные и дорсальные рога, между которыми лежит промежуточная зона. Кроме того, в грудных сегментах различают также боковое выпячивание серого вещества – боковые рога. Все нейронные элементы спинного мозга могут быть подразделены на 4 основные группы: эфферентные нейроны, вставочные нейроны, нейроны восходящих трактов и интраспинальные волокна чувствительных афферентных нейронов. Моторные нейроны сосредоточены в передних рогах, где они образуют специфические ядра, все клетки которых посылают свои аксоны к определенной мышце. Каждое двигательное ядро обычно тянется на несколько сегментов. Поэтому и аксоны мотонейронов, иннервирующих одну и ту же мышцу, покидают спинной мозг в составе нескольких вентральных корешков. Кроме моторных ядер, расположенных в вентральных рогах, выделяются большие скопления нервных клеток в промежуточной зоне серого вещества. Это основное ядро вставочных нейронов спинного мозга. Аксоны вставочных нейронов распространяются как внутри сегмента, так и в ближайшие соседние сегменты. Характерное скопление нервных клеток занимает также дорсальную часть дорсального рога. Эти клетки образуют густые переплетения, а указанная зона получила название желатинозной субстанции Роланда. Наиболее точное и систематизированное представление о топографии нервных клеток серого вещества спинного мозга дает разделение его на последовательные слои, или пластины, в каждой из которых группируются главным образом однотипные нейроны. Хотя послойная типография серого вещества была первоначально выявлена в спинном мозге кошки, она оказалась достаточно универсальной и вполне применима к спинному мозгу, как других позвоночных, так и человека. Согласно этим данным, все серое вещество можно разделить на 10 пластин. Самая первая дорсальная пластина содержит главным образом, так называемые краевые нейроны. Их аксоны проецируются рострально, давая начало спиноталамическому тракту. На краевых нейронах оканчиваются волокна тракта Лиссауэра, который образован смесью первичных афферентных волокон и аксонами проприоспинальных нейронов. Вторая и третья пластины образуют желатинозную субстанцию. Здесь локализуются два основных типа нейронов: более мелкие и относительно крупные нейроны. Хотя тела нейронов второй пластины имеют небольшой диаметр, их дендритные разветвления весьма многочисленны. Аксоны нейронов второй пластины проецируются на тракт Лиссауэра и собственный дорсолатеральный пучок спинного мозга, но многие остаются в пределах желатинозной субстанции. На клетках второй и третьей пластин оканчиваются волокна первичных афферентных нейронов, преимущественно кожной и болевой чувствительности. Четвертая пластина занимает примерно центр дорсального рога. Дендрита нейронов IV слоя проникают в желатинозную субстанцию, а их аксоны проецируются в таламус и боковое цервикальное ядро. Синаптические входы они получают от нейронов желатинозной субстанции, а их аксоны проецируются в таламус и боковое цервикальное ядро. Синаптические входы они получают от нейронов желатинозной субстанции и первичных афферентных нейронов. В целом нервные клетки первой-четвертой пластин захватывают всю вершину дорсального рога и образуют первичную сенсорную область спинного мозга. Сюда проецируются волокна большей частя дорсально-корешковых афферентов от экстерорецепторов, включая кожную и болевую чувствительность. В этой же зоне локализованы нервные клетки, дающие начало нескольким восходящим трактам. В пятой и шестой пластинах локализуются многочисленные типы вставочных нейронов, получающие синаптические входы от волокон заднего корешка и нисходящих путей, в особенности кортико-спинального и руброспинального тракта. В седьмой и восьмой пластинах локализуются проприоспинальные вставочные нейроны, дающие начало длинным аксонам, достигающим нейронов отдаленных сегментов. Здесь заканчиваются афферентные волокна от проприорецепторов, волокна вестибулоспинального и ретикулоспинального трактов, аксоны проприоспинальных нейронов. В девятой пластине располагаются тела a- и g-мотонейронов. Этой области достигают также пресинаптические окончания первичных афферентных волокон от мышечных рецепторов растяжения, окончания волокон нисходящих трактов, кортико-спинальных волокон, терминали аксонов возбуждающих и тормозящих вставочных нейронов. Десятая пластина окружает спинномозговой канал и содержит наряду с нейронами значительное количество глиальных клеток и комиссуральных волокон. Клетки нейроглии спинного мозга на значительном протяжении покрывают поверхность нейронов, причем отростки глиальной клетки направлены, с одной стороны, к телам нейронов, а с другой, часто контактируют с кровеносными капиллярами, являясь посредниками между нервными элементами и источниками их питания. Спинной мозг по восходящим путям передает сигналы в надсегментарные уровни головного мозга, а по нисходящим получает оттуда команды к действию. Восходящие пути передают импульсы от проприоцепторов по волокнам спинобульбарных пучков Голля и Бурдаха и спинномозжечковых путей Говерса и Флексиго, от болевых и температурных рецепторов по латеральному спиноталамическому тракту, от тактильных рецепторов по вентральному спиноталамическому пути и частично по пучкам Голля и Бурдаха. Нисходящие пути проходят в составе кортикоспинальных, или пирамидных, трактов и экстракортикоспинальных, или экстрапирамидных. Дата добавления: 2015-02-05 | Просмотры: 1711 | Нарушение авторских прав |