|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
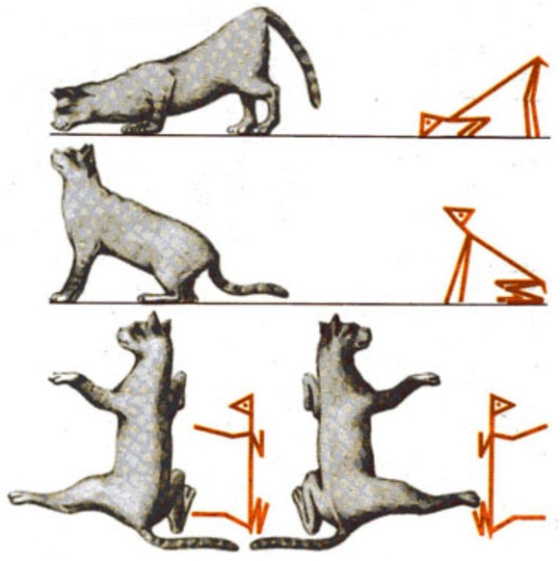
Физиология продолговатого мозга и мостаЗадний мозг, куда относят функционально близкие отделы ГМ - продолговатый мозг и мост, играет важную роль в поддержании тонических и позных рефлексов, участвует в регуляции висцеральных функций и осуществляет проводниковую деятельность. Принято выделять статические и статокинетические рефлексы заднего мозга. К статическим рефлексам относят тонические и позные рефлексы в отсутствие перемещения тела в пространстве. Значение заднего мозга в тонических и позных рефлексах определяется регулирующим влияние на эфферентные нейроны СМ со стороны нейронов вестибулярных ядер (ядра преддверно-улиткового нерва -VIII пары черепных нервов). Важную роль в статических позных рефлексах играют афферентные возбуждения от мышц шеи (отсюда специальное название этих рефлексов – тонические шейные лабиринтные рефлексы). Шейные тонические лабиринтные рефлексы впервые описаны Р. Магнусом на кошках с перерезанным мозгом на уровне шатра мозжечка. Запрокидывание головы кошки назад усиливает тонус мышц-разгибателей в передних конечностях и ослабляет в задних. Наклон головы вниз вызывает противоположенный эффект (рис. 5).
Рис. 5. Шейные тонические рефлексы кошки (по Р. Магнусу)
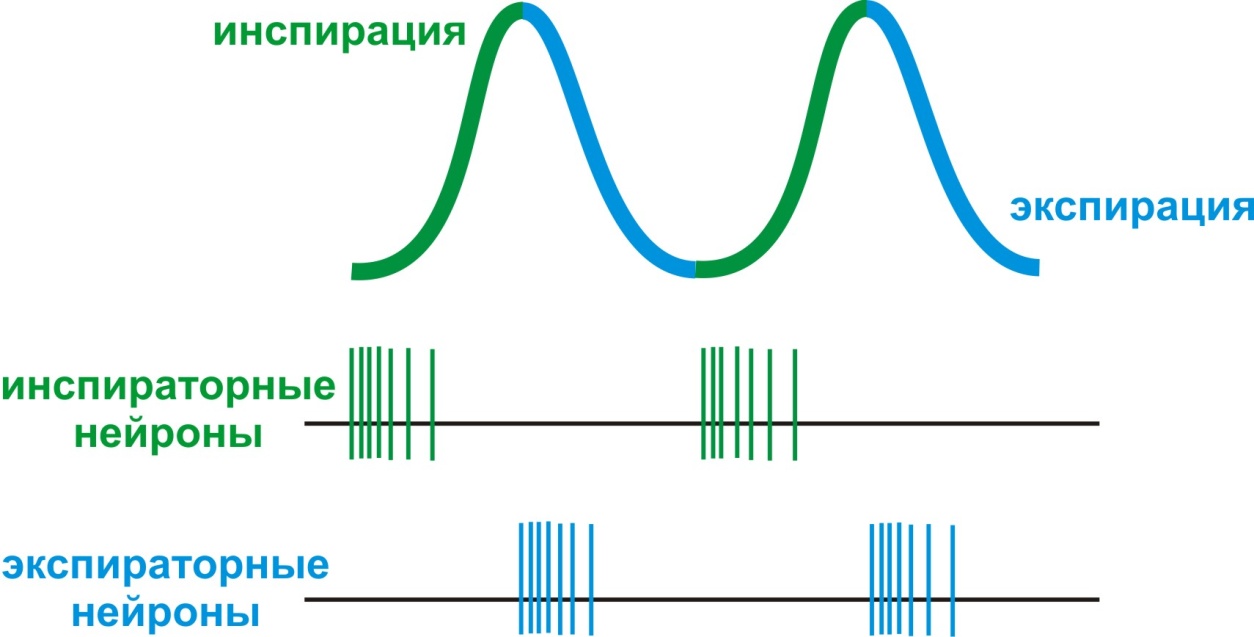
Нарушение равновесия за счет поворота головы вправо или влево относительно продольной оси вызывает компенсаторное усиление тонуса разгибателей тех конечностей, в сторону которых повернута голова (рефлекс вращения по Магнусу, рис. 5). Статокинетические рефлексы обеспечивают поддержание позы тела в пространстве в момент его перемещения. Всем знакомы лифтные рефлексы. В начальный момент движения лифта вверх может произойти сгибание в коленных суставах, но в норме мы не падаем по причине повышения тонуса мышц-разгибателей. При движении лифта вниз, напротив, тонус разгибателей снижается. Почти каждому известны головной и зрительный нистагм. Они, например, проявляются в форме продолжающегося движения головы и глазных яблок после остановки вращения человека вокруг своей оси. В заднем мозге расположены ядра 8-ми черепных нервов (ЧН): V пара - тройничный; VI пара – отводящий; VII – лицевой; VIII пара – преддверно-улитковый; IX пара – языкоглоточный; X пара – блуждающий; XI пара – добавочный; XII пара – подъязычный. На нейронах ядер ЧН замыкаются рефлекторные дуги важнейших безусловных защитных рефлексов: мигательного (роговичного; V и VII), рвотного (V, IX, X), чихательного (V, VII, IX, X, XII), кашлевого (V, VII, IX, X). В заднем мозге расположены центры рефлексов пищевого поведения: жевания, сосания, глотания. Глотательный цепной рефлекс состоит из трех последовательных фаз: ротовой, глоточной и пищеводной. При раздражении пищевым комком рецепторов корня языка и задней стенки зева возбуждение по чувствительным волокнам V, IX и X ЧН направляется к двигательным нейронам двойного ядра (общее двигательное ядро для IX и X ЧН) и тройничного нерва, затем по эфферентным волокнам указанных нервов – к мышцам языка и глотки. В ретикулярной формации продолговатого мозга расположены центры регуляции вегетативных функций: дыхательный центр и сосудодвигательный центр. В XIX веке М. Флуранс обнаружил, что повреждение каудальной части дна IV желудочка приводит к остановке дыхания (укол Флуранса). Позднее Н. А. Миславский (1885) выделил в дыхательном центре две функционально различные части: инспираторную и экспираторную, активность нейронов которых коррелирует с фазами дыхательного цикла (вдохом – инспирацией, и выдохом – экспирацией) Нейроны ДЦ обладают автоматией, поэтому дыхательный центр является ритмоводителем дыхания (задает ритм дыхания частотой 15-16 дыхательных циклов в минуту). По этой причине травмы основания черепа опасны и могут привести к остановке дыхания в результате нарушения функций нейронов ДЦ. Эфферентные возбуждения нейронов ДЦ по ретикуло-спинномозговому пути переключаются на дыхательные мотонейроны СМ и в составе диафрагмального и межреберных нервов направляются к одноименным мышцам. Еще одна особенность ДЦ заключается в реципрокном характере активности инспираторных и экспираторных нейронов (рис. 6).
Рис. 6. Реципрокные взаимосвязи нейронов дыхательного центра Вторым центром регуляции дыхания является пневмотаксический центр (ПТЦ), расположенный в мосту. ПТЦ не является ритмоводителем, но играет важную роль в регуляции глубины и частоты дыхания. Другим жизненно важным центром ретикулярной формации продолговатого мозга является сосудодвигательный центр (СДЦ), локализация которого впервые была определена Ф. В. Овсянниковым (1871). Сосудодвигательный или вазомоторный центр расположен в дорсолатеральном направлении от дна IV мозгового желудочка до пирамид. Установлено, что стимуляция ростральных частей СДЦ увеличивает тонус гладких мышц кровеносных сосудов – вазоконстрикцию (тем самым повышает артериальное давление - АД) и вызывает тахикардию (увеличение частоты сердечных сокращений). Стимуляция каудального отдела СДЦ, напротив, вызывает расширение сосудов (вазодилатацию), снижение АД и брадикардию (снижение частоты сердцебиения). Нейроны, активность которых приводит к вазоконстрикции, получили название прессорных нейронов, а нейроны, вызывающие дилатацию – депрессорных. Активность прессорных нейронов определяется тоническим влиянием симпатической нервной системы. Задний мозг участвует в регуляции вегетативных функций. Это связано с расположением парасимпатических нейронов в составе некоторых черепных нервов: лицевой и языкоглоточный ЧН имеют слюноотделительные ядра (верхнее и нижнее, соответственно). Основным парасимпатическим нервом является блуждающий нерв (n. vagus), который регулирует работу органов грудной и верхней части брюшной полостей. Подробнее о парасимпатических регуляциях мы поговорим в разделе «физиология вегетативной нервной системы». Животные, у которых произведено пересечение ГМ по переднему краю варолиева моста, получили название – бульбарные животные. Такие животные самостоятельно дышат, поддерживают постоянный уровень кровяного давления, но не могут поддерживать постоянную температуру тела и передвигаться (но лягушка, кролик способны осуществлять некоторые виды движений). Дата добавления: 2014-09-03 | Просмотры: 1477 | Нарушение авторских прав |