|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Гормони статевих залозСтатеві залози – парні органи, представлені сім'яниками (у чоловіків) та яєчниками (у жінок). Вони належать до залоз змішаної секреції, оскільки в яєчниках синтезуються яйцеклітини та стероїдні гормони – естрогени та прогестини, а в сім’яниках – сперматозоїди та андрогени. Статеві гормони, забезпечуючи репродуктивну функцію, володіють також потужним анаболічним ефектом, впливаючи на обмін речовин у шкірі, кістках і м’язах. 9.10.1. Чоловічі статеві гормони.синтез і секреція андрогенів (аndros — чоловік, грец.), основними представниками яких є тестостерон і дигідротестостерон (ДГТ), відбувається в яєчках у клітинах Лейдіга (або інтерстиційних клітинах).
Як у яєчках, так і в корі надниркових залоз попередником чоловічих статевих гормонів є холестерол, який або надходить із плазми в складі ЛПНЩ, або утворюється в самих залозах із ацетил-КоА. З нього утворюється проміжний продукт – прегнанолон, який зазнає подальших перетворень за допомогою п’яти мікросомальних ферментів. Утворення та секреція андрогенів знаходиться під контролем лютропіну гіпофіза, секреція якого, своєю чергою, регулюється гонадоліберином гіпоталамуса. Лютропін стимулює стероїдогенез і продукцію тестостерону після з’єднання з рецепторами на плазматичній мембрані клітин Лейдіга за аденілатциклазним механізмом. Утворений цАМФ стимулює ферментативне розщеплення бічного ланцюга холестеролу. Зворотній зв'язок реалізується через інгібування вивільненого гонадоліберину або шляхом гальмування секреції лютропіну. Метаболізм тестостерону здійснюються двома шляхами: через утворення 17-кетостероїдів або через утворення ДГТ, естрадіолу та андростендіолу. У печінці 17-кетостероїди утворюють кон'югати з глюкуроновою та сірчаною кислотами тавиводяться з організму. Транспорт андрогенів до органів-мішеней здійснюється специфічним глікопротеїном плазми, що отримав назву тестостерон-естрадіол-зв'язуючого глобуліну, який має спорідненість до тестостерону, ДГТ і естрадіолу. До дії тестостерону чутливі ембріональна Вольфова протока, сперматогонії, м'язи, нирки, кістки, мозок, а до ДГТ – передміхурова залоза, сім'яні міхурці, зовнішні статеві органи. шкіра. У цитоплазмі багатьох клітин-мішеней тестостерон перетворюється на ДГТ, що має вищу спорідненість до рецепторів порівняно з тестостероном. Андрогени утворюють комплекс з рецептором, який зазнавши певних конформаційних змін, проникає в ядро і взаємодіє з акцепторними білками хроматину, активуючи специфічні гени, білкові продукти котрих опосередковують біологічні ефекти андрогенів, які різняться в різні періоди життя організму. У плода чоловічої статі під впливом цих гормонів відбувається маскулінізіція мозку. Оскільки андрогени володіють потужним анаболічним ефектом, то в препубертатний період їх підвищений рівень стимулює стрімкий ріст кісток, збільшення скелетних м’язів, формування вторинних статевих ознак – розвиток хрящів гортані та формування чоловічого тембру голосу, ріст волосся на обличчі й тілі, секрецію сальних залоз. Разом з фолітропіном вони активують сперматогенез. Індукуючи синтез білків, в тому числі ферментних, андрогени впливають на енергетичні процеси, зокрема, активують ферменти циклу Кребса. Це підвищує аеробний розпад вуглеводів і розпад ліпідів у тканинах, внаслідок чого збільшується утворення енергії. Гормони підвищують синтез фосфоліпідів у різних клітинних мембранах і знижують вміст холестерину, але, порівняно з естрогенами, меншою мірою. Отже, чоловічі статеві гормони беруть участь у процесах статевого диференціювання та сперматогенезу; розвитку вторинних статевих ознак і формуванні психофізичного статусу чоловіка; стимуляції анаболічних процесів.
У дитячому віці незрілі яєчники продукують незначну кількість гормонів, у пубертатний період чутливість гіпоталамо-гіпофізарної системи до впливу ЛГ і ФСГ знижується. На початку кожного менструального циклу секреція цих гормонів спричинює розвиток фолікулів, дозрівання яких супроводжується секрецією естрогенів. Концентрація естрадіолу зростає, чутливість гіпофізарних клітин до гонадоліберину підвищується і, за механізмом зворотного зв’язку зростає секреція ЛГ і ФСГ. Підвищення концентрації ЛГ спричинює овуляцію, після якої утворюється жовте тіло, яке окрім естрадіолу починає синтезувати прогестерон. Зростання концентрації цього гормона в плазмі крові за механізмом зворотного зв’язку пригнічує активність гіпоталамо-гіпофізарної системи: гальмується секреція ЛГ і ФСГ, жовте тіло руйнується, знижується продукція стероїдів яєчниками і, як наслідок, настає менструація, після чого цикл повторюється (рис. 14.15). У випадку настання вагітності жовте тіло продовжує функціонувати та продукувати прогестерон, але на пізніх етапах вагітності прогестерон більшою мірою синтезується в плаценті, яка, забезпечуючи зв'язок між системами кровоплину зародка та матері, продукує також низку інших гормонів: хоріонічний гонадотропін (подібний за активністю до лютропіну), основна функція якого полягає в підтриманні існування жовтого тіла до того часу, поки плацента не почне в необхідній кількості виробляти прогестерон; плацентарний лактоген, якому властиві лактотропний, лютеотропний і соматотропний ефекти (подібно гормону росту); тиреотропін. Метаболізм естрогенів відбувається в печінці: метаболітами естрадіолу є естрон і естріол, які інактивуються в реакціях кон’югації з УДФГК або ФАФС; прогестерон перетворюється на прегнандіол, який зв’язується з глюкуроновою кислотою.
Рис. 14.15. Циклічні зміни концентрацій жіночих статевих гормонів та гонадотропінів впродовж менструального циклу. ФСГ-фолікулостимулюючий гормон; ЛГ-лютеїнізуючий гормон
Естрогени зв’язуються зі специфічними внутрішньоклітинними рецепторами в тканинах матки, піхви, маткових труб, молочних залоз, аденогіпофіза та гіпоталамуса та, подібно до інших стероїдних гормонів, регулюють транскрипцію структурних генів, індукуючи синтез понад 50 різноманітних білків, які беруть участь у прояві їх фізіологічних ефектів. у жіночому організмі естрогени стимулюють ріст і розвиток матки та молочних залоз; стимулюють гіпертрофію та гіперплазію міометрію; стимулюють початок фази проліферації ендометрію; регулюють матково-плацентарний кровоплин; сприяють дозріванню епітеліальних клітин слизової оболонки піхви; разом з простагландином F2a вони сенсибілізують міометрій до дії окситоцину під час пологів, чим сприяють підвищенню тонусу, збудливості та скоротливості матки; викликають посилення перистальтики маткових труб у момент овуляції, що сприяє прискоренню міграції сперматозоонів; під впливом естрогенів шийковий епітелій матки посилено продукує слиз, що сприяє проникненню сперматозоїдів. Вони чинять анаболічний вплив на кістки та хрящі, підтримують структуру шкіри та кровоносних судин, сприяють утворенню оксиду азоту в судинах гладких м’язів, чим спричинюють їх розширення та посилення тепловіддачі. Естрогени стимулюють синтез транспортних білків тиреоїдних і статевих гормонів, індукують синтез факторів згортання крові (ІІ, VІІ, ІХ, Х), знижують концентрацію антитромбіну ІІІ. Ці гормони чинять вплив на обмін ліпідів: зростання швидкості синтезу ЛПВЩ і гальмування утворення ЛПНЩ, викликане естрогенами, призводить до зниження вмісту холестеролу в крові. Прогестерон, який утворюється в жовтому тілі під час менструації (лютеїнова фаза) секретується також фетоплацентарним комплексом під час вагітності. Під час фолікулярної фази його концентрація незначна, тому що дія цього гормона спрямована на забезпечення репродуктивної функції організму жінки: він відповідає за підвищення температури тіла на 0,2 – 0,5 °C, яке відбувається зразу після овуляції та зберігається впродовж лютеїнової фази менструального циклу; сприяє виникненню фази секреції в ендометрії та утворенню децидуальної оболонки під час вагітності; зменшує збудливість та скоротливу активність матки шляхом зниження чутливості міометрію до речовин, що викликають його скорочення; підтримує тонус істміко-цервікального відділу матки; стимулює гіпертрофію м’язових волокон під час вагітності; готує молочні залози до секреції молока; зменшує продукцію шийкового слизу, робить його непрохідним для сперматозоїдів; знижує скоротливу активність маткових труб. Крім зазначених ефектів, у великій концентрації прогестерон може взаємодіяти з рецепторами клітин ниркових канальців, конкуруючи в такий спосіб з альдостероном, у результаті чого останній втрачає здатність стимулювати реабсорбцію води. Цей гормон також чинить вплив і на ЦНС, викликаючи певні особливості поведінки жінки в передменструальний період. Стан, викликаний зниженням рівня синтезу та секреції жіночих статевих гормонів, називається гіпогонадизмом. Він може бути зумовлений процесами, які безпосередньо ушкоджують яєчники і призводять до їх недостатності (первинний гіпогонадизм), або зниженням гонадотропної функції гіпофіза (вторинний гіпогонадизм). При дефіциті естрогенів до періоду статевого дозрівання відбувається затримка розвитку органів статевої системи та формування вторинних статевих ознак з порушенням перебігу статевих циклів; спостерігаються зміни в метаболічних процесах - знижується рівень кальцію і фосфату, має місце гіперліпемія, негативний азотистий баланс. Дефіцит прогестерону порушує перебіг статевого циклу та є причиною викиднів. Зміна концентрації естрогенів і прогестерону в слині жінок репродуктивного віку може певною мірою відображати період овуляції та слугувати для моніторингу впливу циркадіанних (добових) ритмів на адреналову активність (вміст кортизолу). Існує залежність між дефіцитом естрогенів та розвитком дифузного остеопорозу в міжзубних перетинках та альвеолярних відростках, особливо в період менопаузи та після тотальної оваріектомії.
9.11. Ейкозаноїди Ейкозаноїди – це біологічно активні речовини, які синтезуються в багатьох клітинах із полієнових жирних кислот, що містять 20 атомів карбону («ейкоза» – 20, грец). Вони включають у себе простагландини, тромбоксани та лейкотрієни тощо – високоактивні регулятори клітинних функцій. Їх період півжиття надзвичайно короткий, тому свої ефекти вони чинять як «гормони місцевої дії», впливаючи на метаболізм тих клітин, що їх синтезують за автокринним механізмом, а на оточуючі клітини – за паракринним механізмом.
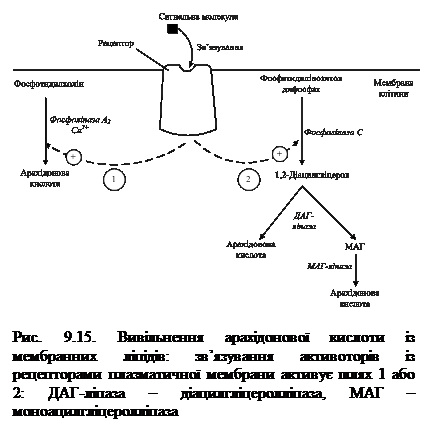
Головними субстратами для синтезу ейкозаноїдів слугують арахідонова, ейкозапентаєнова та ейкозатрієнова кислоти. Вони входять до складу гліцерофосфоліпідів мембран (рис. 9.15). Під впливом асоційованої з мембраною фосфоліпази А2 (рідше фосфоліпази С) жирна кислота відщеплюється від гліцерофосфоліпіда та використовується для синтезу ейкозаноїдів. У різних тканинах арахідонова та інші ейкозанові кислоти можуть використовуватися в трьох основних напрямках: циклооксигеназний шлях призводить до утворення простагландинів, простациклінів і тромбоксанів; ліпооксигеназний шлях забезпечує синтез лейкотрієнів, ліпоксинів і гідроксіейкозатетраєноатів (ГЕТЄ); система окиснення за участі цитохрому Р-450 бере участь у синтезі епоксидів.
Біосинтез ПГ і тромбоксану починається із перетворення арахідонової кислоти на ендопероксидні проміжні продукти (ПГG2 і ПГН2) (рис. 9.16). Обидві реакції каталізуються поліферментним комплексом простагландинсинтазою, який складається із двох компонентів — циклооксигенази (ЦОГ) і пероксидази. В активному центрі ЦОГ міститься тирозин, а пероксидази – гем. В організмі присутні два типи ЦОГ: ЦОГ-1 конститутивний фермент, який синтезується з постійною швидкістю, тоді як синтез ЦОГ-2 зростає під час запалення та індукується відповідними медіаторами – цитокінами. Обидва типи циклооксигеназ каталізують включення 4 атомів кисню в арахідонову кислоту з утворенням п’ятичленного кільця. До утворення ПГН2 шлях синтезу різних ПГ однаковий, подальші перетворення специфічні для кожного типу клітин. так, у тромбоцитах під впливом тромбоксансинтази з ПГН2 утворюється тромбоксан А2 (ТхА2), який володіє потужною судинозвужувальною дією, а в клітинах ендотелію – простациклін (ПГІ2), який розширює судини. 9.11.2. Біосинтез лейкотрієнів. Лейкотрієни також синтезуються з ейкозанових кислот, однак, на відміну від ПГ, у їх структурі відсутні цикли і вони мають 3 спряжені подвійні зв’язки (звідки назва «лейкотрієн»). Утворення лейкотрієнів відбувається переважно у клітинах крові – лейкоцитах різних класів, тромбоцитах, макрофагах, що відображає провідну роль лейкотрієнів у реакціях запалення, згортанні крові, алергійних реакціях і імунних процесах.
Синтезовані ейкозаноїди надходять із клітин у міжклітинне середовище і реалізують свої функції, взаємодіючи із специфічними рецепторами мембран клітин-мішеней. Інактивація цих речовин відбувається дуже швидко (від кількох секунд до кількох хвилин) під дією відповідних ферментів. Продукти перетворень ейкозаноїдів виводяться з сечею. 9.11.3. Основні біологічні ефекти ейкозаноїдів. Всі ейкозаноїди, незважаючи на короткий період піврозпаду, мають надзвичайно широкий спектр дії та високу біологічну активність. Вони діють на клітини через спеціальні рецептори. Деякі з них зв’язані з аденілатциклазною системою та протеїнкіназою А (ПГЕ, ПГD, ПГІ2); інші діють через зростання рівня кальцію в цитозолі клітин мішеней (ПГF, ТхА2, лейкотрієни). У різних тканинах вони виконують різні, іноді протилежні функції (табл. 9.4). Таблиця 9.4. Біологічні функції ейкозаноїдів
Біологічні функції ПГ пов'язані з впливом на скорочувальну функцію гладких м'язів, проте окремі ПГ мають різні фізіологічні ефекти у певних тканинах-мішенях. Так, наприклад, ПГ А та Е знижують артеріальний тиск при гіпертонічній хворобі; ПГЕ спричинює розслаблення гладеньких м'язів бронхів і трахеї, тоді як ПГF, навпаки, викликає їх скорочення. Характерною особливістю ПГЕ2 та ПГF2 (містяться в сім'яній рідині) є їх стимулюючий вплив стосовно м'язів матки, що сприяє переміщенню сперматозоїдів у порожнину фаллопієвих труб, де відбувається запліднення. ПГ Е1 гальмує базальну та стимульовану секрецію хлоридної кислоти, захищає клітини слизової оболонки від ушкоджуючих хімічних подразників. Препарати ПГЕ1 мізопростол, цитотек є ефективними засобами сучасної терапії виразкової хвороби. Протилежну дію мають простациклін і тромбоксан. Простациклін синтезується у клітинах ендотелію судин і перешкоджає згортанню крові, гальмуючи агрегацію тромбоцитів і розширюючи судини, а тромбоксан А2 утворюється у тромбоцитах і сприяє їх агрегації, а також скорочує гладкі м'язи судин. Звільнення тромбоксану із агрегатів тромбоцитів і дифузія в стінку судин зумовлюють їх звуження. Таким чином, відносна активність тромбоксану і простацикліну визначає ймовірність утворення тромбів і місцевого спазму судин. Дисбаланс цих сполуку бік тромбоксанів відіграє важливу роль в утворенні атеросклеротичних бляшок. Механізм дії тромбоксану полягає у підвищенні виходу Са2+ із внутрішньоклітинних депо в цитозоль тромбоцитів. Іони кальцію, в свою чергу, стимулюють скорочувальні білки тромбоцитів, а також вивільнення із тромбоцитів вмісту їх гранул (серотоніну, катехоламінів, АДФ). Простациклін підвищує рівень цАМФ у тромбоцитах, що перешкоджає мобілізації Са2+ і синтезу тромбоксану. Простагландини також можуть впливати на рівень цАМФ у клітинах, зокрема простагландини групи Е активують аденілатциклазу в ендокринних залозах, але гальмують у жировій тканині. Концентрація цАМФ у клітині, своєю чергою, впливає на синтез ПГ.
Аспірин – протизапальний препарат, механізм дії якого полягає в інгібуванні циклооксигенази і, як наслідок, пригніченні синтезу медіаторів запалення. Циклооксигеназа незворотно інгібується шляхом ацетилування серину в активному центрі фермента (рис. 9.18). Однак, ефект аспірину нетривалий, оскільки експресія гена цього фермента не порушується і синтезуються його нові молекули. Інші нестероїдні препарати (ібупрофен, ацетамінофен тощо) теж знижують синтез ПГ за конкурентним механізмом, зв’язуючись з активним центром фермента. Стероїдні препарати володіють значно потужнішою протизапальною дією, оскільки індукують синтез білків-ліпокортинів, котрі інгібують активність фосфоліпази А2 і, запобігаючи вивільненню субстрату для синтезу ейкозаноїдів – арахідонової кислоти, зменшують їх синтез.
Дата добавления: 2015-11-28 | Просмотры: 1625 | Нарушение авторских прав |
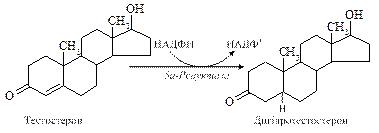
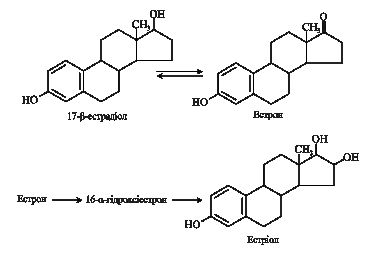 9.10.2. Жіночі статеві гормони. У яєчниках синтезуються естрогени (oistros – пристрасний потяг, грец.) – С18-стероїди та прогестини – С21-стероїди, серед яких найактивнішими є 17b-естрадіол і прогестерон. У людини основними джерелами естрогенів є яєчники і плацента, частина гормонів утворюється також у корі надниркових залоз, сім'яниках і деяких інших тканинах. Їх синтез відбувається шляхом ароматизації андрогенів внаслідок складного процесу, що включає три етапи гідроксилування, кожний з яких вимагає НАДФН2 і молекулярного кисню. Основний представник прогестинів – прогестерон утворюється з холестеролу в жовтому тілі та, незначною мірою, у плаценті та корі надниркових залоз.
9.10.2. Жіночі статеві гормони. У яєчниках синтезуються естрогени (oistros – пристрасний потяг, грец.) – С18-стероїди та прогестини – С21-стероїди, серед яких найактивнішими є 17b-естрадіол і прогестерон. У людини основними джерелами естрогенів є яєчники і плацента, частина гормонів утворюється також у корі надниркових залоз, сім'яниках і деяких інших тканинах. Їх синтез відбувається шляхом ароматизації андрогенів внаслідок складного процесу, що включає три етапи гідроксилування, кожний з яких вимагає НАДФН2 і молекулярного кисню. Основний представник прогестинів – прогестерон утворюється з холестеролу в жовтому тілі та, незначною мірою, у плаценті та корі надниркових залоз.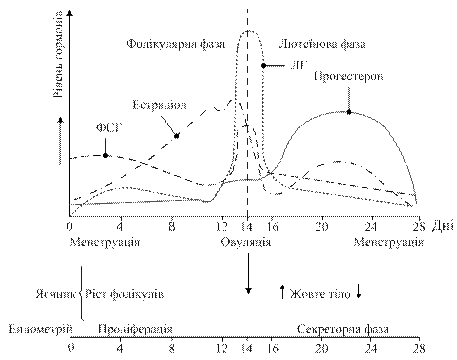
 Вони беруть участь у багатьох процесах: регулюють тонус гладком’язевих клітин, впливаючи в такий спосіб на регуляцію артеріального тиску, стан бронхів, кишок і матки; секрецію води та натрію нирками, впливають на утворення тромбів. Різні типи ейкозаноїдів беруть участь у розвитку запального процесу, адже такі його ознаки як біль, набряк, гарячка значною мірою обумовлені дією саме ейкозаноїдів. Надлишок ейкозаноїдів призводить до низки захворювань (алергійні реакції, бронхіальна астма).
Вони беруть участь у багатьох процесах: регулюють тонус гладком’язевих клітин, впливаючи в такий спосіб на регуляцію артеріального тиску, стан бронхів, кишок і матки; секрецію води та натрію нирками, впливають на утворення тромбів. Різні типи ейкозаноїдів беруть участь у розвитку запального процесу, адже такі його ознаки як біль, набряк, гарячка значною мірою обумовлені дією саме ейкозаноїдів. Надлишок ейкозаноїдів призводить до низки захворювань (алергійні реакції, бронхіальна астма).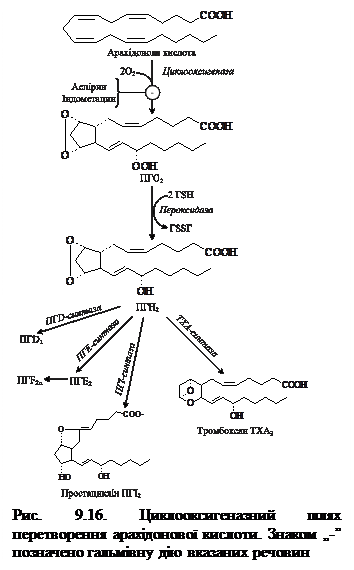 9.11.1. Синтез простагландинів і тромбоксанів. Простагландини (ПГ) вперше були виявлені У. Ейлером у передміхуровій залозі (рrostate gland – передміхурова залоза, англ), тому й одержали свою назву. згодом було досліджено, що ПГ знаходяться у всіх органах і тканинах, за винятком еритроцитів, впливають на низку фізіологічних функцій. Вони містять у своїй молекулі 20 атомів вуглецю, п'ять з яких утворюють циклопентанове кільце. За особливостями хімічної будови ПГ поділяють на 4 групи: ПГЕ, ПГF, ПГА, ПГВ, у кожній групі розрізняють підгрупи, що позначаються цифрою, яка вказує на число подвійних зв'язків у молекулі (ПГЕ1, ПГЕ2 тощо).
9.11.1. Синтез простагландинів і тромбоксанів. Простагландини (ПГ) вперше були виявлені У. Ейлером у передміхуровій залозі (рrostate gland – передміхурова залоза, англ), тому й одержали свою назву. згодом було досліджено, що ПГ знаходяться у всіх органах і тканинах, за винятком еритроцитів, впливають на низку фізіологічних функцій. Вони містять у своїй молекулі 20 атомів вуглецю, п'ять з яких утворюють циклопентанове кільце. За особливостями хімічної будови ПГ поділяють на 4 групи: ПГЕ, ПГF, ПГА, ПГВ, у кожній групі розрізняють підгрупи, що позначаються цифрою, яка вказує на число подвійних зв'язків у молекулі (ПГЕ1, ПГЕ2 тощо).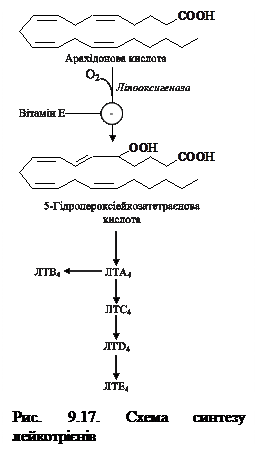 Під дією ферменту ліпооксигенази та молекули кисню арахідонова кислота окиснюється до 5-гідропероксіейкозатетраєнової кислоти (5-ГПЕТК) – метаболічного попередника біологічно активних лейкотрієнів. У залежності від типу тканини ліпоксигенази діють на арахідонову кислоту в 5-ому, 12-ому та 15-ому положеннях. Так, наприклад, у поліморфоноядерних лейкоцитах міститься в основному 5-ліпоксигеназа, у тромбоцитах – 12-ліпоксигеназа, в еозинофілах – 15-ліпоксигеназа. У лейкоцитах і опасистих клітинах 5-ГПЕТК перетворюється на епоксид-лейкотрієн А4 (ЛТА4), де нижній індекс означає кількість подвійних зв’язків (рис. 9.17). Інші типи лейкотрієнів утворюються з ЛТА4: ЛТА4 шляхом конденсації із трипептидом глутатіоном утворює ЛТС4, а із нього після відщеплення однієї та двох амінокислот синтезуються ЛТD4 і ЛТЕ4; ЛТВ4 утворюється під дією епоксидгідроксилази в лейкоцитах і клітинах епітелію судин.
Під дією ферменту ліпооксигенази та молекули кисню арахідонова кислота окиснюється до 5-гідропероксіейкозатетраєнової кислоти (5-ГПЕТК) – метаболічного попередника біологічно активних лейкотрієнів. У залежності від типу тканини ліпоксигенази діють на арахідонову кислоту в 5-ому, 12-ому та 15-ому положеннях. Так, наприклад, у поліморфоноядерних лейкоцитах міститься в основному 5-ліпоксигеназа, у тромбоцитах – 12-ліпоксигеназа, в еозинофілах – 15-ліпоксигеназа. У лейкоцитах і опасистих клітинах 5-ГПЕТК перетворюється на епоксид-лейкотрієн А4 (ЛТА4), де нижній індекс означає кількість подвійних зв’язків (рис. 9.17). Інші типи лейкотрієнів утворюються з ЛТА4: ЛТА4 шляхом конденсації із трипептидом глутатіоном утворює ЛТС4, а із нього після відщеплення однієї та двох амінокислот синтезуються ЛТD4 і ЛТЕ4; ЛТВ4 утворюється під дією епоксидгідроксилази в лейкоцитах і клітинах епітелію судин.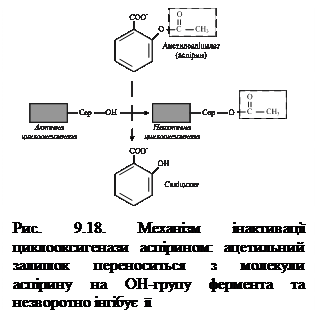 Лейкотрієни С4, D4 і Е4 секретуються тканинними базофілами у відповідь на дію антигенів і викликають сильне скорочення гладких м'язів бронхів і трахеї. Із дією лейкотрієнів пов'язують утруднене дихання у хворих на бронхіальну астму. Також вони впливають на гладкі м'язи травного тракту, судин, знижують силу скорочень міокарда, збільшують проникність стінок судин. Ейкозаноїди також виконують функцію медіаторів запалення і діють на всіх етапах цього процесу, внаслідок чого збільшується проникність капілярів, транссудат і лейкоцити проникають крізь судинну стінку, що обумовлює ознаки запалення – почервоніння, біль, гарячку, набряк.
Лейкотрієни С4, D4 і Е4 секретуються тканинними базофілами у відповідь на дію антигенів і викликають сильне скорочення гладких м'язів бронхів і трахеї. Із дією лейкотрієнів пов'язують утруднене дихання у хворих на бронхіальну астму. Також вони впливають на гладкі м'язи травного тракту, судин, знижують силу скорочень міокарда, збільшують проникність стінок судин. Ейкозаноїди також виконують функцію медіаторів запалення і діють на всіх етапах цього процесу, внаслідок чого збільшується проникність капілярів, транссудат і лейкоцити проникають крізь судинну стінку, що обумовлює ознаки запалення – почервоніння, біль, гарячку, набряк.