|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
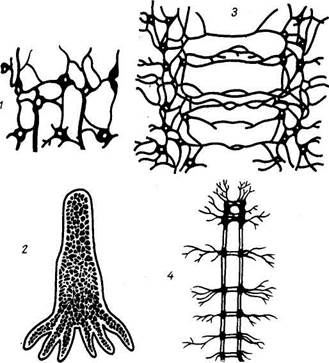
РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫФилогенез нервной системы в кратких чертах сводится к следующему. У простейших одноклеточных организмов (амеба) нервной системы еще нет, а связь с окружающей средой осуществляется при помощи жидкостей, находящихся внутри и вне организма, — гуморальная (humor — жидкость), до-нервная, форма регуляции. В дальнейшем, когда возникает нервная система, появляется и другая форма регуляции — нервная. По мере развития нервной системы нервная регуляция все больше подчиняет себе гуморальную, так что образуется единая нейрогуморальная регуляция при ведущей роли нервной системы. Последняя в процессе филогенеза проходит ряд основных этапов (рис. 265). / этап — сетевидная нервная система. На этом этапе (кишечнополостные) нервная система, например гидры, состоит из нервных клеток, многочисленные отростки которых соединяются друг с другом в разных направлениях, образуя сеть, диффузно пронизывающую все тело животного. При раздражении любой точки тела возбуждение разливается по всей нервной сети и животное реагирует движением всего тела. Отражением этого этапа у человека является сетевидное строение интрамуральнои нервной системы пищеварительного тракта. // этап — узловая нервная система. На этом этапе (беспозвоночные) нервные клетки сближаются в отдельные скопления или группы, причем из скоплений клеточных тел получаются нервные узлы — центры, а из скоплений отростков — нервные стволы — нервы. При этом в каждой клетке число отростков уменьшается и они получают определенное направление. Соответственно сегментарному строению тела животного, например у кольчатого червя, в каждом сегменте имеются сегментарные нервные узлы и нервные стволы. Последние соединяют узлы в двух направлениях: поперечные стволы связывают узлы данного сегмента, а продольные — узлы разных сегментов. Благодаря этому нервные импульсы, возникающие в какой-либо точке тела, не разливаются по всему телу, а распространяются по поперечным стволам в пределах данного сегмента. Продольные стволы связывают нервные сег-
Рис. 265. Этапы развития нервной системы. 1, 2 — диффузная нервная система гидры; 3,4 — узловая нервная система кольчатого червя. менты в одно целое. На головном конце животного, который при движении вперед соприкасается с различными предметами окружающего мира, развиваются органы чувств, в связи с чем головные узлы развиваются сильнее остальных, являясь прообразом будущего головного мозга. Отражением этого этапа является сохранение у человека примитивных черт (разбросанность на периферии узлов и микроганглиев) в строении вегетативной нервной системы. /// этап — трубчатая нервная система. На первоначальной ступени развития животных особенно большую роль играл аппарат движения, от совершенства которого зависит основное условие существования животного — питание (передвижение в поисках пищи, захватывание и поглощение ее). У низших многоклеточных развился перистальтический способ передвижения, что связано с непроизвольной мускулатурой и ее местным нервным аппаратом. На более высокой ступени перистальтический способ сменяется скелетной моторикой, т. е. передвижением с помощью системы жестких рычагов — поверх мышц (членистоногие) и внутри мышц (позвоночные). Следствием этого явилось образование произвольной (скелетной) мускулатуры и центральной нервной системы, координирующей перемещение отдельных рычагов моторного скелета. Такая центральная нервная система у хордовых (ланцетник) возникла в виде метамерно построенной нервной трубки с отходящими от нее сегментарными нервами ко всем сегментам тела, включая и аппарат движения,— туловищный мозг. У позвоночных и человека туловищный мозг становится спинным. Таким образом, появление туловищного мозга связано с усовершенствованием в первую очередь моторного вооружения животного. Наряду с этим уже у ланцетника имеются и рецепторы (обонятельный, световой). Дальнейшее развитие нервной системы и возникновение головного мозга обусловлены преимущественно усовершенствованием рецепторного вооружения. Так как большинство органов чувств возникает на том конце тела животного, который обращен в сторону движения, т. е. вперед, то для восприятия поступающих через них внешних раздражений развивается передний конец туловищного мозга и образуется головной мозг, что совпадает с обособлением переднего конца тела в виде головы — цефализация (cephal — голова). Е. К. Сепп в учебнике по нервным болезням1 дает упрощенную, но удобную для изучения схему филогенеза головного мозга, которую мы и приводим. Согласно этой схеме, на I этапе развития головной мозг состоит из трех отделов: заднего, среднего и переднего, причем из этих отделов в первую очередь (у низших рыб) особенно развивается задний, или ромбовидный, мозг (rhombencephalon). Развитие заднего мозга происходит под влиянием рецепторов акустики и гравитации (рецепторы VIII пары черепных нервов), имеющих ведущее значение для ориентации в водной среде. В дальнейшей эволюции задний мозг дифференцируется на продолговатый мозг, являющийся переходным отделом от спинного мозга к головному и потому называемый myelencephalon (myelos — спинной мозг, епсёр-halon — головной), и собственно задний мозг — metencephalon, из которого развиваются мозжечок и мост. В процессе приспособления организма к окружающей среде путем изменения обмена веществ в заднем мозге как наиболее развитом на этом этапе отделе центральной нервной системы возникают центры управления жизненно важными процессами растительной жизни, связанными, в частности, с жаберным аппаратом (дыхание, кровообращение, пищеварение и др.). Поэтому в продолговатом мозге возникают ядра жаберных нервов (группа X пары — вагуса). Эти жизненно важные центры дыхания и кровообращения остаются в продолговатом мозге человека, чем объясняется смерть, наступающая при повреждении продолговатого мозга. На II этапе (еще у рыб) под влиянием зрительного рецептора особенно развивается средний мозг, mesencephalon. На III этапе, в связи с окончательным переходом животных из водной среды в воздушную, усиленно развивается обонятельный рецептор, воспринимающий содержащиеся в воздухе химические вещества, сигнализирующие своим запахом о добыче, опасности и других жизненно важных явлениях окружающей природы. Под влиянием обонятельного рецептора развивается передний мозг — prosencephalon, вначале имеющий характер чисто обнятельного мозга. В дальнейшем передний мозг разрастается и дифференцируется на промежуточный— diencephalon и конечный — telencephalon. В конечном мозге как в высшем отделе центральной нервной системы появляются центры для всех видов чувствительности. Однако нижележащие центры не исчезают, а сохраняются, подчиняясь центрам вышележащего этажа. Следовательно, с каждым новым этапом развития головного мозга возникают новые центры, подчиняющие себе старые. Происходит как бы передвижение функциональных центров к головному концу и одновременное подчинение филогенетически старых зачатков новым. В результате центры слуха, впервые возникшие в заднем мозге, имеются также в среднем и переднем, центры зрения, возникшие в среднем, имеются и в переднем, а центры обоняния — только в переднем мозге. Под влиянием обонятельного рецептора развивается небольшая часть переднего мозга, называемая поэтому обонятельным мозгом (rhinencephalon), который покрыт корой серого вещества — старой корой (paleocortex). Совершенствование рецепторов приводит к прогрессивному развитию переднего мозга, который постепенно становится органом, управляющим всем поведением животного. Различают две формы поведения животного: инстинктивное, основанное на видовых реакциях (безусловные рефлексы), и индивидуальное, основанное на опыте индивида (условные рефлексы). Соответственно этим двум формам поведения в конечном мозге развивается две группы центров серого вещества: базальные узлы, имеющие строение ядер
(ядерные центры), и кора серого вещества, имеющая строение сплошного Необходимой формацией для осуществления высшей нервной деятель* ности является новая кора, расположенная на поверхности полушарий и приобретающая в процессе филогенеза шестислойное строение. Благодаря усиленному развитию новой коры конечный мозг у высших позвоночных превосходит все остальные отделы головного мозга, покрывая их, как плащом (pallium). Развивающийся новый мозг (neencephalon) оттесняет в глубину старый мозг (обонятельный), который как бы свертывается в виде гиппокампа (hyppocampus), остающегося по-прежнему обонятельным центром. В результате плащ, т. е. новый мозг (neencephalon), резко преобладает над остальными отделами мозга — старым мозгом (paleencephalon). Итак, развитие головного мозга совершается под влиянием развития рецепторов, чем и объясняется, что самый высший отдел головного мозга — кора (серое вещество) — представляет, как учит И. П. Павлов, совокупность корковых концов анализаторов, т. е. сплошную воспринимающую (рецептор-ную) поверхность. Дальнейшее развитие мозга у человека подчиняется иным закономерностям, связанным с его социальной природой. Кроме естественных органов тела, имеющихся и у животных, человек стал пользоваться орудиями труда. Орудия труда, ставшие искусственными органами, дополнили естественные органы тела и составили техническое вооружение человека. С помощью этого вооружения человек приобрел возможность не только приспосабливаться самому к природе, как это делают животные, но и приспосабливать природу к своим нуждам. Труд, как уже отмечалось, явился решающим фактором становления человека, а в процессе общественного труда возникло необходимое для общения людей средство — речь. «Сначала труд, а затем и вместе с ним членораздельная речь явились двумя самыми главными стимулами, под влиянием которых мозг обезьяны постепенно превратился в человеческий мозг, который, при всем своем сходстве с обезьяньим, далеко превосходит его по величине и совершенству» {Маркс К., Энгельс Ф. Соч., 2-е изд., т. 20, с. 490). Это совершенство обусловлено максимальным развитием конечного мозга, особенно его коры — новой коры (neocortex). Кроме анализаторов, воспринимающих различные раздражения внешнего мира и составляющих материальный субстрат конкретно-наглядного мышления, свойственного животным {первая сигнальная система действительности, по И. П. Павлову), у человека возникла способность абстрактного, отвлеченного мышления с помощью слова, сначала слышимого (устная речь) и позднее видимого (письменная речь). Это составило вторую сигнальную систему, по И. П. Павлову, которая в развивающемся животном мире явилась «чрезвычайной прибавкой к механизмам нервной деятельности» (И. П. Павлов). Материальным субстратом второй сигнальной системы стали поверхностные слои новой коры. Поэтому кора конечного мозга достигает своего наивысшего развития у человека. Таким образом, эволюция нервной системы сводится к прогрессивному развитию конечного мозга, который у высших позвоночных и особенно у человека в связи с усложнением нервных функций достигает огромных размеров. Изложенные закономерности филогенеза обусловливают эмбриогенез нервной системы человека. Нервная система происходит из наружного заро-
Рис. 266. Стадии эмбриогенеза нервной системы; поперечный схематический разрез. А — медуллярная пластинка; В, С — медуллярная бороздка; D, Е— нервная трубка; I — роговой листок (эпидермис); 2 — нейральные гребни. дышевого листка, или эктодермы (см. «Введение»). Эта последняя образует продольное утолщение, называемое медуллярной пластинкой (рис. 266). Медуллярная пластинка скоро углубляется в медуллярную бороздку, края которой (медуллярные валики) постепенно становятся выше и затем срастаются друг с другом, превращая бороздку в трубку {мозговая трубка). Мозговая трубка представляет собой зачаток центральной части нервной системы. Задний конец трубки образует зачаток спинного мозга, передний расширенный конец ее путем перетяжек расчленяется на три первичных мозговых пузыря, из которых происходит головной мозг во всей его сложности. Нервная пластинка первоначально состоит только из одного слоя эпителиальных клеток. Во время замыкания ее в мозговую трубку количество клеток в стенках последней увеличивается, так что возникает три слоя: внутренний (обращенный в полость трубки), из которого происходит эпителиальная выстилка мозговых полостей (эпендима центрального канала спинного мозга и желудочков головного); средний, из которого развивается серое вещество мозга (зародышевые нервные клетки — нейробласты); наконец, наружный, почти не содержащий клеточных ядер, развивающийся в белое вещество (отростки нервных клеток — нейриты). Пучки нейритов нейробластов распространяются или в толще мозговой трубки, образуя белое вещество мозга, или же выходят в мезодерму и затем соединяются с молодыми мышечными клетками (миобластами). Таким путем возникают двигательные нервы. Чувствительные нервы возникают из зачатков спинномозговых узлов, которые заметны уже по краям медуллярной бороздки у места перехода ее в кожную эктодерму. Когда бороздка смыкается в мозговую трубку, зачатки смещаются на ее дорсальную сторону, располагаясь по средней линии. Затем клетки этих зачатков перемещаются вентрально и располагаются вновь по бокам мозговой трубки в виде так называемых нейральных гребней. Оба нейральных гребня перешнуровываются четкообразно по сегментам дорсальной стороны зародыша, вследствие чего получается на каждой стороне ряд спинномозговых узлов, ganglia spinalia. В головной части мозговой трубки они доходят только до области заднего мозгового пузырька, где образуют зачатки узлов чувствительных черепных нервов. В ганглиозных зачатках развиваются нейробласты, принимающие вид биполярных нервных клеток, один из отростков которых врастает в мозговую трубку, другой идет на периферию, образуя чувствительный нерв. Благодаря сращению на некотором протяжении от начала обоих отростков получаются из биполярных так называемые ложные униполярные клетки с одним отростком, делящимся в форме буквы «Т», являющиеся характерными для спинномозговых узлов взрослого. Центральные отростки клеток, проникающие в спинной мозг, составляют задние корешки спинномозговых нервов, а периферические отростки, разрастаясь вентрально, образуют (вместе с вышедшими из спинного мозга эфферентными волокнами, составляющими передний корешок) сме- 17 Анатомия человека шанный спинномозговой нерв. Из нейральных гребней возникают также зачатки вегетативной нервной системы, о чем подробно см. «Вегетативная (автономная) нервная система». ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА СПИННОЙ МОЗГ Развитие спинного мозга. Как уже отмечалось, филогенетически спинной мозг (туловищный мозг ланцетника) появляется на III этапе развития нервной системы (трубчатая нервная система). В это время головного мозга еще нет, поэтому туловищный мозг имеет центры для управления всеми процессами организма, как вегетативными, так и анимальными (висцеральные и соматические центры). Соответственно сегментарному строению тела туловищный мозг имеет сегментарное строение, он состоит из связанных между собой невромеров, в пределах которых замыкается простейшая рефлекторная дуга. Метамерное строение спинного мозга сохраняется и у человека, чем и обусловливается наличие у него коротких рефлекторных дуг. С появлением головного мозга (этап кефализации) в нем возникают высшие центры управления всем организмом, а спинной мозг попадает в подчиненное положение. Спинной мозг не остается только сегментарным аппаратом, а становится и проводником импульсов от периферии к головному мозгу и обратно, в нем развиваются двусторонние связи с головным мозгом. Таким образом, в процессе эволюции спинного мозга образуется два аппарата: более старый сегментарный аппарат собственных связей спинного мозга и более новый надсегментарный аппарат двусторонних проводящих путей к головному мозгу. Такой принцип строения наблюдается и у человека. Решающим фактором образования туловищного мозга является приспособление к окружающей среде при помощи движения. Поэтому строение спинного мозга отражает способ передвижения животного. Так, например, у пресмыкающихся, не имеющих конечностей и передвигающихся с помощью туловища (например, у змеи), спинной мозг развит равномерно на всем протяжении и не имеет утолщений. У животных, пользующихся конечностями, возникает два утолщения, при этом, если более развиты передние конечности (например, крылья птиц), то преобладает переднее (шейное) утолщение спинного мозга; если более развиты задние конечности (например, ноги страуса), то увеличено заднее (поясничное) утолщение; если в ходьбе участвуют и передние, и задние конечности (четвероногие млекопитающие), то одинаково развиты оба утолщения. У человека в связи с более сложной деятельностью руки как органа труда шейное утолщение спинного мозга дифференцировалось сильнее, чем поясничное. Отмеченные факторы филогенеза играют роль в развитии спинного мозга и в онтогенезе. Спинной мозг развивается из нервной трубки, из ее заднего отрезка (из переднего возникает головной мозг). Из вентрального отдела трубки образуются передние столбы серого вещества спинного мозга (клеточные тела двигательных нейронов), прилегающие к ним пучки нервных волокон и отростки названных нейронов (двигательные корешки). Из дорсального отдела возникают задние столбы серого вещества (клеточные тела вставочных нейронов), задние канатики (отростки чувствительных нейронов). Таким образом, вентральная часть мозговой трубки является первично двигательной, а дорсальная — первично чувствительной. Деление на моторную (двигательную) и сенсорную (чувствительную) области простирается на всю нервную трубку и сохраняется в стволе головного мозга.
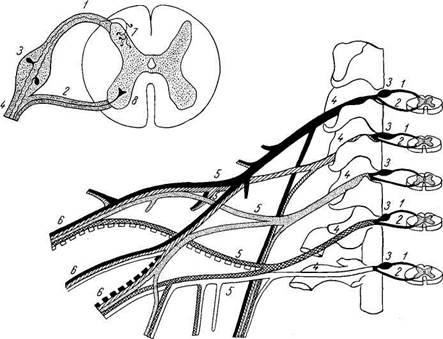
а - вид спереди; б - вид сзади. Твердая и паутинная оболочки разрезаны. Сосудистая оболочка снята. Римскими цифрами обозначен порядок расположения шейных (С), грудных (Th), поясничных (L) и крестцовых (S) спинномозговых нервов; / — intumescentia cervicalis; 2 — gangl. spinale; 3 — dura mater medullae spinalis; 4 - intumescentia lumbosacra-lis; 5 —conus medullaris; 6 — cauda equina. Из-за редукции каудальной части спинного мозга получается тонкий тяж из нервной ткани, будущая filum terminate. Первоначально, на 3-м месяце утробной жизни, спинной мозг занимает весь позвоночный канал, затем позвоночник начинает расти скорее, чем мозг, вследствие чего конец последнего постепенно перемещается кверху (краниально). При рождении конец спинного мозга уже находится на уровне III поясничного позвонка, а у взрослого достигает высоты I — II поясничного позвонка. Благодаря такому «восхождению» спинного мозга отходящие от него нервные корешки принимают косое направление (рис. 267). Строение спинного мозга
Спинной мозг, medulla spinalis (греч. myelos), лежит в позвоночном канале и у взрослых представляет собой длинный (45 см у мужчин и 41—42 см у женщин), несколько сплюснутый спереди назад цилиндрический тяж, который вверху (краниально) непосредственно переходит в продолговатый мозг, а внизу (каудально) оканчивается коническим заострением, conus medullaris, на уровне II поясничного позвонка (см. рис. 267). Знание этого факта имеет практическое значение (чтобы не повредить спинной мозг при поясничном проколе с целью взятия спинномозговой жидкости или с целью спинномозговой анестезии, надо вводить иглу шприца между остистыми отростками III и IV поясничных позвонков). От conus medullaris отходит книзу так называемая концевая нить, filum terminate, представляющая атрофированную нижнюю часть спинного мозга, которая внизу состоит из продолжения оболочек спинного мозга и прикрепляется ко II копчиковому позвонку. Спинной мозг на своем протяжении имеет два утолщения, соответствующих корешкам нервов верхней и нижней конечностей: верхнее из них называется шейным утолщением, intumescentia cervicalis, а нижнее — пояснично-крестцовым, intumescentia lumbosacralis. Из этих утолщений более обширно 17*
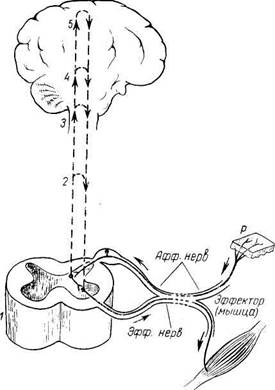
Рис. 268. Элементы периферической нервной системы (схема). /-radix posterior; 2 - radix anterior; 3 - gangl. spinale; 4 - truncus n. spinalis; 5-plexus; 6 - ветви сплетения; 7 — задний рог; 8 — передний рог. пояснично-крестцовое, но более дифференцировано шейное, что связано с более сложной иннервацией руки как органа труда. Образовавшимися вследствие утолщения боковых стенок спинномозговой трубки и проходящими по средней линии передней и задней продольными бороздами: глубокой fissura mediana anterior, и поверхностной, sulcus medianus posterior, спинной мозг делится на две симметричные половины — правую и левую; каждая из них в свою очередь имеет слабо выраженную продольную борозду, идущую по линии входа задних корешков {sulcus posterolateralis) и по линии выхода передних корешков (sulcus anterolateralis). Эти борозды делят каждую половину белого вещества спинного мозга на три продольных канатика: передний — funiculus anterior, боковой— funiculus lateralis и задний — funiculus posterior. Задний канатик в шейном и верхнегрудном отделах делится еще промежуточной бороздкой, sulcus intermedius posterior, на два пучка: fasciculus gracilis и fasciculus cuneatus. Оба эти пучка под теми же названиями переходят вверху на заднюю сторону продолговатого мозга. На той и другой стороне из спинного мозга выходят двумя продольными рядами корешки спинномозговых нервов. Передний корешок, radix ventral is s. anterior, выходящий через sulcus anterolateralis, состоит из нейритов двигательных (центробежных, или эфферентных) нейронов, клеточные тела которых лежат в спинном мозге, тогда как задний корешок, radix dorsalis s. posterior, входящий в sulcus posterolateralis, содержит отростки чувствительных (центростремительных, или афферентных) нейронов, тела которых лежат в спинномозговых узлах. На некотором расстоянии от спинного мозга двигательный корешок прилегает к чувствительному (рис. 268) и они вместе образуют ствол спинномозгового нерва, truncus n. spinalis, который невропатологи выделяют под именем канатика, funiculus. При воспалении канатика (фуникулит) возникают сегментарные расстройства одновременно двигательной и чувствительной сфер; при заболевании корешка (радикулит) наблюдаются сегментарные нарушения одной сферы — или чувствительной, или двигательной, а при воспалении ветвей нерва (неврит) расстройства соответствуют зоне распространения данного нерва. Ствол нерва обычно очень короткий, так как по выходе из межпозвоночного отверстия нерв распадается на свои основные ветви. В межпозвоночных отверстиях вблизи места соединения обоих корешков задний корешок имеет утолщение — спинномозговой узел, ganglion spinale, содержащий ложноуниполярные нервные клетки (афферентные нейроны) с одним отростком, который делится затем на две ветви: одна из них, центральная, идет в составе заднего корешка в спинной мозг, другая, периферическая, продолжается в спинномозговой нерв. Таким образом, в спинномозговых узлах отсутствуют синапсы, так как здесь лежат клеточные тела только афферентных нейронов. Этим названные узлы отличаются от вегетативных узлов периферической нервной системы, так как в последних вступают в контакты вставочные и эфферентные нейроны. Спинномозговые узлы крестцовых корешков лежат внутри крестцового канала, а узел копчикового корешка — внутри мешка твердой оболочки спинного мозга. Вследствие того что спинной мозг короче позвоночного канала, место выхода нервных корешков не соответствует уровню межпозвоночных отверстий. Чтобы попасть в последние, корешки направляются не только в стороны от мозга, но еще и вниз, при этом тем отвеснее, чем ниже они отходят от спинного мозга. В поясничной части последнего нервные корешки спускаются к соответствующим межпозвоночным отверстиям параллельно filum terminate, облекая ее и conus medullaris густым пучком, который носит название конского хвоста, cauda equina (см. рис. 267). Внутреннее строение спинного мозга. Спинной мозг состоит из серого вещества, содержащего нервные клетки, и белого вещества, слагающегося из миелиновых нервных волокон. А. Серое вещество, substantia grisea, заложено внутри спинного мозга и окружено со всех сторон белым веществом. Серое вещество образует две вертикальные колонны, помещенные в правой и левой половинах спинного мозга. В середине его заложен узкий центральный канал, canal is central is, спинного мозга, проходящий во всю длину последнего и содержащий спинномозговую жидкость. Центральный канал является остатком полости первичной нервной трубки. Поэтому вверху он сообщается с IV желудочком головного мозга, а в области conus medullaris заканчивается расширением — концевым желудочком, ventriculus terminalis. Серое вещество, окружающее центральный канал, носит название промежуточного, substantia intermedia central is. В каждой колонне серого вещества два столба: передний, columna anterior, и задний, columna posterior. На поперечных разрезах спинного мозга эти столбы имеют вид рогов: переднего, расширенного, cornu anterius, и заднего, заостренного, cornu posterius. Поэтому общий вид серого вещества на фоне белого напоминает букву «Н». Серое вещество состоит из нервных клеток, группирующихся в ядра, расположение которых в основном соответствует сегментарному строению спинного мозга и его первичной трехчленной рефлекторной дуге. Первый, чувствительный, нейрон этой дуги лежит в спинномозговых узлах, периферический отросток которого начинается рецепторами в органах и тканях, а центральный в составе задних чувствительных корешков проникает через sulcus posterolateralis в спинной мозг. Вокруг верхушки заднего рога образуется пограничная зона белого вещества, представляющая собой совокупность центральных отростков клеток спинномозговых узлов, заканчивающихся в спинном мозге. Клетки задних рогов образуют отдельные группы или ядра, воспринимающие из сомы различные виды чувствительности, — соматически-чувствительные ядра. Среди них выделяются: грудное ядро, nucleus thoracicus (coliimna thoracica), наиболее выраженное в грудных сегментах мозга; находящееся на верхушке рога студенистое вещество, substantia gelatinosa, а также так называемые собственные ядра, nuclei proprii. Заложенные в заднем роге клетки образуют вторые, вставочные, нейроны. В сером веществе задних рогов разбросаны также рассеянные клетки, так называемые пучковые клетки, аксоны которых проходят в белом веществе обособленными пучками волокон. Эти волокна несут нервные импульсы от определенных ядер спинного мозга в его другие сегменты или служат для связи с третьими нейронами рефлекторной дуги, заложенными в передних рогах того же сегмента. Отростки этих клеток, идущие от задних рогов к передним, располагаются вблизи серого вещества, по его периферии, образуя узкую кайму белого вещества, окружающего серое со всех сторон. Это собственные пучки спинного мозга, fasciculi proprii. Вследствие этого раздражение, идущее из определенной области тела, может передаваться не только на соответствующий ей сегмент спинного мозга, но захватывать и другие. В результате простой рефлекс может вовлекать в ответную реакцию целую группу мышц, обеспечивая сложное координированное движение, остающееся, однако, безусловнорефлекторным. Передние рога содержат третьи, двигательные, нейроны, аксоны которых, выходя из спинного мозга, составляют передние, двигательные, корешки. Эти клетки образуют ядра эфферентных соматических нервов, иннервирующих скелетную мускулатуру, — соматически-двигательные ядра. Последние имеют вид коротких колонок и лежат в виде двух групп — медиальной и латеральной. Нейроны медиальной группы иннервируют мышцы, развившиеся из дорсальной части миотомов (аутохтонная мускулатура спины), а латеральной — мышцы, происходящие из вентральной части миотомов (вентролатеральные мышцы туловища и мышцы конечностей); чем дисталь-нее иннервируемые мышцы, тем латеральнее лежат иннервирующие их клетки. Наибольшее число ядер содержится в передних рогах шейного утолщения спинного мозга, откуда иннервируются верхние конечности, что определяется участием последних в трудовой деятельности человека. У последнего в связи с усложнением движений руки как органа труда этих ядер значительно больше, чем у животных, включая антропоидов. Таким образом, задние и передние рога серого вещества имеют отношение к иннервации органов животной жизни, особенно аппарата движения, в связи с усовершенствованием которого в процессе эволюции и развивался спинной мозг. Передний и задний рога в каждой половине спинного мозга связаны между собой промежуточной зоной серого вещества, которая в грудном и поясничном отделах спинного мозга, на протяжении от I грудного до II — III поясничных сегментов особенно выражена и выступает в виде бокового рога, cornu late rale. Вследствие этого в названных отделах серое вещество на поперечном разрезе приобретает вид бабочки. В боковых рогах заложены клетки, иннервирующие вегетативные органы и группирующиеся в ядро, которое носит название columna intermediolaterdlis. Нейриты клеток этого ядра выходят из спинного мозга в составе передних корешков. Б. Белое вещество, substantia alba, спинного мозга состоит из нервных отростков, которые составляют три системы нервных волокон: 1. Короткие пучки ассоциативных волокон, соединяющих участки спинного мозга на различных уровнях (афферентные и вставочные нейроны). 2. Длинные центростремительные (чувствительные, афферентные). 3. Длинные центробежные (двигательные, эфферентные). Первая система (коротких волокон) относится к собственному аппарату спинного мозга, а остальные две (длинных волокон) составляют проводниковый аппарат двусторонних связей с головным мозгом. Собственный аппарат включает серое вещество спинного мозга с задними и передними корешками и собственными пучками белого вещества (fasciculi proprii), окаймляющими серое в виде узкой полосы. По развитию собственный аппарат является образованием филогенетически более старым и потому сохраняет примитивные черты строения — сегментарность, отчего его называют также сегментарным аппаратом спинного мозга в отличие от остального несегментированного аппарата двусторонних связей с головным мозгом. Таким образом, нервный сегмент — это поперечный отрезок спинного мозга и связанных с ним правого и левого спинномозговых нервов, развившихся из одного невротома (невромера). Он состоит из горизонтального слоя белого и серого вещества (задние, передние и боковые рога), содержащего нейроны, отростки которых проходят в одном парном (правом и левом) спинномозговом нерве и его корешках (см. рис. 268). В спинном мозге различают 31 сегмент, которые топографически делятся на 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1 копчиковый. В пределах нервного сегмента замыкается короткая рефлекторная дуга (см. рис. 264). Так как собственный сегментарный аппарат спинного мозга возник тогда, когда еще не было головного, то функция его — это осуществление тех реакций в ответ на внешнее и внутреннее раздражения, которые в процессе эволюции возникли раньше, т. е. врожденных реакций. Аппарат двусторонних связей с головным мозгом филогенетически более молодой, так как возник лишь тогда, когда появился головной мозг. По мере развития последнего разрастались кнаружи и проводящие пути, связывающие спинной мозг с головным (рис. 269). Этим объясняется тот факт, что белое вещество спинного мозга как бы окружило со всех сторон серое вещество. Благодаря проводниковому аппарату собственный аппарат спинного мозга связан с аппаратом головного мозга, который объединяет работу всей нервной системы. Нервные волокна группируются в пучки, а из пучков составляются видимые невооруженным глазом канатики: задний, боковой и передний. В заднем канатике (рис. 270), прилежащем к заднему (чувствительному) рогу, лежат пучки восходящих нервных волокон; в переднем канатике, прилежащем к переднему (двигательному) рогу, лежат пучки нисходящих нервных волокон; наконец, в боковом канатике находятся и те и другие. Кроме канатиков, белое вещество находится в белой спайке, comissura alba, образующейся вследствие перекреста волокон спереди от substantia intermedia centralis; сзади белая спайка отсутствует. Задние канатики содержат волокна задних корешков спинномозговых нервов, слагающиеся в две системы: 1. Медиально расположенный тонкий пучок, fasciculus grdcilis. 2. Латерально расположенный клиновидный пучок, fasciculus cuneatus. к коре головного мозга сознательную проприоцептивную (мышечно-суставное чувство) и кожную (чувство стереогноза — узнавание предметов на ощупь) чувствительность, имеющую отношение к определению положения тела в пространстве, а также тактильную чувствительность. Боковые канатики содержат следующие пучки: А. Восходящие.
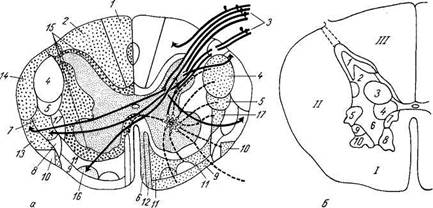
Рис. 270. Внутреннее строение спинного мозга; поперечный разрез. а — схема проводящих путей спинного мозга: слева изображено местоположение восходящих, справа — нисходящих систем волокон: 1 — fasc. gracilis; 2 — fasc. cuneatus; 3 — radix posterior; 4 — tr. corti-cospinalis lateralis; 5 — tr. rubrospinalis; 6 — tr. tectospinalis; 7 — tr. spinothalamicus lateralis; 8 — tr. spinote-ctalis; 9 — tr. vestibulospinalis; 10 — tr. olivospinalis; 11 — tr. reticulospinalis; 12 — tr. corticospinalis anterior; 13 — tr. spinocerebellaris anterior; 14 — tr. spinocerebellaris posterior; 15 — fascc. proprii; 16 — tr. spinothalamicus anterior; 17 —tr. thalamospinalis; б — ядра серого вещества (в грудном отделе); 1 — substantia gelatinosa; 2 — nucl. proprius cornu posterioris; 3 — nucl. thoracicus; 4 — nucl, interrnedjome-dialis; 5 — columna intermediolateralis; 6, 7, 8, 9, 10 — пять двигательных ядер переднего рога; /, //, III — соответственно передний, боковой и задний канатики белого вещества. К заднему мозгу: 1) tractus spinocerebellaris posterior, задний спинно-мозжечковый путь, располагается в задней части бокового канатика по его периферии; 2) tractus spinocerebellaris anterior, передний спинно-мозжеч-ковый путь, лежит вентральнее предыдущего. Оба спинно-мозжечковых тракта проводят бессознательные проприоцеп-тивные импульсы (бессознательная координация движений). К среднему мозгу: 3) tractus spinotectalis, спинно-покрыщечный путь, прилегает к медиальной стороне и передней части tractus spinocerebellaris anterior. К промежуточному мозгу: 4) tractus spinothalamicus laterdlis прилегает с медиальной стороны к tractus spinocerebellaris anterior, тотчас позади tractus spinotectalis. Он проводит в дорсальной части тракта тем-' пературные раздражения, а в вентральной — болевые; 5) tractus spinothalamicus anteriror s. ventrdlis аналогичен предыдущему, но располагается кпереди от соименного латерального и является путем проведения импульсов осязания, прикосновения (тактильная чувствительность). По последним данным, этот тракт располагается в переднем канатике. Б. Нисходящие. От коры большого мозга: 1) латеральный корково-спинномозго-вой (пирамидный) путь, tractus corticospinalis (pyramidalis) lateralis. Этот тракт является сознательным эфферентным двигательным путем. От среднего мозга: 2) tractus rubrospinalis. Он является бессознательным эфферентным двигательным путем. От заднего мозга: 3) tractus olivospindlis, лежит вентральнее tractus spinocerebellaris anterior, вблизи переднего канатика. Передние канатики содержат нисходящие пути. От коры головного мозга: 1) передний корково-спинномозговой (пирамидный) путь, tractus corticospinalis (pyramidalis) anterior, составляет с латеральным пирамидным пучком общую пирамидную систему. От среднего мозга: 2) tractus tectospinalis, лежит медиальнее пирамидного пучка, ограничивая fissura mediana anterior. Благодаря ему осуществляются рефлекторные защитные движения при зрительных и слуховых раздражениях — зрительно-слуховой рефлекторный тракт. Ряд пучков идет к передним рогам спинного мозга от различных ядер продолговатого мозга, имеющих отношение к равновесию и координации движений, а именно: 3) от ядер вестибулярного нерва — tractus vestibulospinalis — лежит на гра 4) от formatio reticularis — tractus reticulospindlis anterior, лежит в средней 5) собственно пучки, fasciculi prbprii, непосредственно прилегают к се Дата добавления: 2015-12-16 | Просмотры: 789 | Нарушение авторских прав |
 1 Сепп Е. К., Цукер М. Б., Шмид Е. В. Нервные болезни.-М.: Медгиз, 1954.
1 Сепп Е. К., Цукер М. Б., Шмид Е. В. Нервные болезни.-М.: Медгиз, 1954.
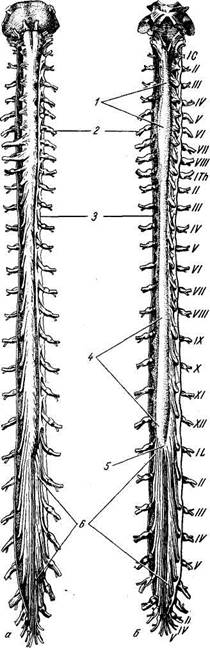 Рис. 267. Спинной мозг.
Рис. 267. Спинной мозг.