|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Физиология возбудимых тканей. 1.Физиология как наука. Понятия о функции1.Физиология как наука. Понятия о функции. Методы физиол. исследований. ПП, механизм возникновения, физиологическая роль. Физиология - наука об объективной закономерности функций живого организма, про их взаимодействие и изменения под влиянием внешней среды. Функция- деятельность и свойство клеток, органа, системы, организма, что проявляется как физиологический процесс или совокупность процессов. Методы – наблюдение, эксперимент (острый, хронический), инструментальные или лабораторные исследования, моделирование. ПП – разница потенциалов между внутренней и наружной поверхностями мембран, существующих в состояние покоя. Равен -50,- 90 мВ, существует до момента действия раздражителя. Механизм возникновения: 1.разность концентрации ионов, 2.различная проницаемость для различных ионов, 3.работа K/Na – АТФазы. Физиол. роль – для создания градиента концентрации.
Возбудимость – это частный случай раздражимости (Раздражимость – свойство живой материи активно изменять характер своей жизнедеятельности при действии раздражителя). Возбудимыми называют ткани, клетки которых способны генерировать потенциал действия, а невозбудимыми – клетки которых не способны к генерации потенциала действия. Порог деполяризации – min сдвиг мембранного потенциала, необходимого для того, что бы исходная его величина достигла критического значения деполяризации.(ПД)
Блокаторы Na каналов – тетродотоксин, тетраметиламоний.
3.ПП, механизмы его происхождения, методы регистрации и параметры. Понятие о деполяризации и гиперболизации мембран клеток. ПП – разница потенциалов между внутренней и наружной поверхностями мембран, существующих в состояние покоя. Равен -50,- 90 мВ. Механизм возникновения: 1.разность концентрации ионов, 2.различная проницаемость для различных ионов, 3.работа K/Na – АТФазы. Методы регистрации-при помощи техники микро электродов. Деполяризация - уменьшение ПП, лежит в основе ПД.(активная- изменение K/Na проницаемости, пасивная-под катодом -) Гиперполяризация - увеличение разницы потенциалов между внешней и внутренней поверхностями мембраны возбудимой клетки.(активная- изменение K/Na проницаемости,пасивная-под анодом +)
4.ПД, механизм происхождения, методы регистрации и параметры. ПД – быстрое изменение ПП, в ответ на действие пороговых и сверхпороговых раздражителей. Механизм-в ответ на пороговый раздражитель возникает ЛП, что достигая КУД переходит в деполяризацию мембраны за счет повышения проницаемости для Na,на овершуте Na каналы инактивируются, и из-за активации K каналов происходит реполяризация, из-за следового выхода Na- следовая деполяризация, а из-за выхода K – следовая гиперполяризация. Методы регистрации- при помощи микроэлектродов. Параметры +30,70, 100,120 длительность:нейрон- 1-3мсек, миокард 300мсек, гладкая мускулатура до 10сек. 5.Сравнительная характеристика ЛП и ПД. Изменения возбудимости клеток во время развития ЛП и ПД. ЛП-ответная реакция на раздражения до пороговое но близкое к пороговому, и не распространяется.
6.Механизм раздражения клеток электрическим током. КУД и порог деполяризации. Значение параметров электрических импульсов для возникновения возбуждения. Электроды помещают в следующем порядке: анод+ внутрь клетки (он повышает уровень деполяризации, снижая порог возбудимости) катод – наружу. КУД-величина мембранного потенциала, при достижение которой возникает ПД Порог деполяризации-min значение ПП, необходимое для того, что бы исходная его величина достигла КУД. Закон Пфлюгера – при замыкание под катодом -, при размыкание под анодом+ Катодическая депрессия Вериго - снижение возбудимости ткани под катодом – при длительном действии на нее постоянного электрического тока.
Реобаза- min сила раздражителя, вызывающая ПД за неопределенное время. Полезное время- min время действия раздражителя силой в 1 реобазу. Хронаксия- min время действия раздражителя силой в 2 реобазы.
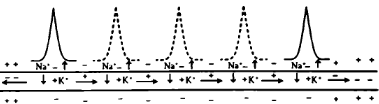
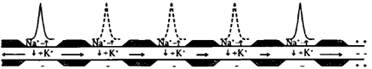
7.Механизмы проведения возбуждения по НВ. Схема проведения возбуждения по миелиновому и без миелиновому НВ. Согласно «кабельной» теории А. Германа, возбуждение проводится непрерывно по без миелиновым и прерывисто(сальтаторно) по миелиновым волокнам. Без миелиновые волокна на всем протяжении имеют одинаковую электропроводность и сопротивление, вследствие чего ЛП распространяется на рядом расположенные не возбужденные участки, идет последовательно, медленно. Миелиновые волокна имеют изолирующий слой что дает возможность «сократить» длину волокна, ПД перепрыгивает с одного безмиелинового участка на другой.
8.Нервно-мышечный синапс, его структура. Механизм и закономерности нервно- мышечной передачи возбуждения. Блокаторы нервно- мышечных синапсов. Синапс- это специализированная структура между НК и мышечной, где нервный импульс может влиять на деятельность постсинаптической клетки, возбуждая или тормозя ее.(электрические щель-2Нм, химическая – 50 Нм) Состоит из: пресинаптической и постсинаптической мембран,синаптическая щель между ними(синаптическая задержка 0.5-1мс) ПД деполяризует пресинаптическое окончание(аксон мот нейрона) ионы Ca поступая из внеклеточной жидкости, связываются с кальмодулином, комплекс Ca-кальмодулин толкают визикулы с медиатором(Ах) к синаптической щели. В синаптической щели Ах дифундирует 0.1-0.2мс к пост синаптической мембране(Ах-эстераза 40% Ах разрушает, 60% захватывает пре-СМ) Ах взаимодействуя с N-холино рецепторами активирует Na каналы и возникает деполяризация концевой пластинки(ЛП) что затем вызывает генерацию ПД на близлежащем мышечном волокне. АХ-ВПСП N-холин рецепторы(Na)/Глицин-ТПСП М-холин рецепторы(K) N-блокаторы пахикарин, пирилен, бензогексоний. М-блокаторы атропин, скополамин, платифилин, метацил. 1блок Ca каналов, 2блок синтеза медиатора, 3блок рецептора,4инактивация Na каналов, 5 Ах-эстераза.
9.Виды и типы мышечных сокращений, их механизмы. Утомление мышц. Изотоническое-длинна мышц меняется а напряжение нет, Изометрическое-напряжение нарастает а длинна нет. Ауксотопическое-сочетанное Одиночное-при однократном раздражении, Тетаническое-длительное сокращение мышц,под действием ритмического раздражителя *гладкий-если последующее раздражение попадает в момент укорочения *зубчатый- если последующее раздражение попадает в момент расслабления Тоническое-длительное сокращение мышц неритмической природы, небольшое по силе. ПД в мышечных волокнах непрерывен (как в без миелиновом волокне) >активация Ca каналов СПР>Са +тропонин (кальмодулин в гладкой мускулатуре) > смещают тропомиозин оголяя активный участок актина>миозиновая головка прикрепляется к активному центру энергия АТФ тратится на «гребневое движение» шейкой 1шаг= 20Нм. Затем закрываются Са каналы, тропомиозин обратно конформируется закрывая активный участок актина, и обратное скольжение под силой гравитации и эластичности возвращает саркомер к исходному размеру. Мышечное утомление-снижение силы мышечно работы со временем до полной ее остановки, зависит от ритма работы, величины груза и тренированности. Но изначально истощаются синапсы.
Латентный-10мс Сокрашение-50мс Расслабление-50мс
Смотри пред ведущий.
ЦНС, ВНС Дата добавления: 2015-12-15 | Просмотры: 746 | Нарушение авторских прав |
 2.Возбудимость. Порог деполяризации мембраны. Механизмы изменения возбудимости клеток под влиянием разных факторов.
2.Возбудимость. Порог деполяризации мембраны. Механизмы изменения возбудимости клеток под влиянием разных факторов.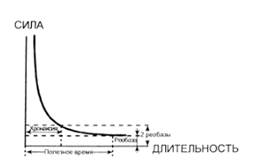
 10.Электрохимическое сопряжение возбуждения в скелетных мышцах и механизм мышечного сокращения. Роль ионов Са в механизмах мышечного сокращения.
10.Электрохимическое сопряжение возбуждения в скелетных мышцах и механизм мышечного сокращения. Роль ионов Са в механизмах мышечного сокращения.