|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Механізм першого вдиху новонародженої дитини.1. Після перерізки пуповини в крові дитини накопичується вуглекислота, знижується рН крові та знижується парціальний тиск кисню – стимуляція центральних та периферичних хеморецепторів – збудження дихального центру – збудження інспіраторних нейронів – скорочення м’язів вдиху. 2. Стимулювати перший вдих новонародженого можна дією на різні рецептори, оскільки аферентна інервація дихального центру дуже різноманітна: - стимулюючи терморецептори – раніше дитину опускали то в холодну то в теплу воду; - діючи на механорецептори (поплескування, пощипування шкіри); - діючи на вестибулорецептори – дитину можна опускати головою вниз, нахиляти зі сторони в сторону. 3. Після народження дитини треба зняти “рефлекс пірнальника”, котрий блокує настання вдиху через наявність рідини в дихальних шляхах. Щоб зняти цей рефлекс цю рідину періодично аспірують.
110.Гуморальные метаболические факторы, стимулирующие внешнее дыхание. До компонентів крові, які стимулюють зовнішнє дихання, відносяться Ро2, рН, Рсо2. Найбільшого значення для регуляції зовнішнього дихання має вміст в крові СО2 (Рсо2), менше – вміст йонів Н+, саме менше – вміст О2 (Ро2). Залежність вентиляції легень від Рсо2 досліджують при диханні газовою сумішшю, в якій Ро2 залишається незмінним а Рсо2 поступово підвищується. При цьому відмічено, що підвищення Рсо2 в газовій суміші на 1 мм.рт.ст. супроводжується підвищенням ХОД на 2 – 3л/хв. При підвищенні Рсо2 артеріальної крові від 40 до 60 мм.рт.ст. ХОД зростає від 7 до 65 л/хв. Підвищення ХОД у відповідь на підвищення Рсо2 проходить лише до певної межі. Якщо Рсо2 в артеріальній крові стає вищим від 70 мм.рт.ст., ХОД знижується, що є результатом прямого пригнічуючого впливу СО2 на нейрони дихального центру. При зниженні рН артеріальної крові нижче 7,4 вентиляція легень (ХОД) також підвищується. Зменшення рН на 0,1 призводить до збільшення ХОД на 2 л/хв. Гіпервентиляція при збільшенні концентрації йонів Н+ супроводжується зниженням Рсо2 в крові. Тому, якщо дослідження проводити при постійному Рсо2 в артеріальній крові (40 мм.рт.ст.), гіпервентиляція виражена більше (заштрихована лінія). Зниження Ро2 в артеріальній крові також супроводжується гіпервентиляцією. Звертає на себе увагу той факт, що вплив зниження Ро2 впливає на ХОД, якщо величина показника стає нижчою за 50 – 60 мм.рт.ст. (норма – 100 мм.рт.ст.). тобто, гіпервентиляція розвивається у відповідь на значне зниження Ро2 і ступінь її невелика. Як і в випадку з рН, реакція стає більш вираженою, якщо при дослідженні стабілізувати Рсо2 на рівні 40 мм.рт.ст. (заштрихована лінія). Таким чином, підвищення Рсо2 призводить до гіпервентиляції відразу ж (тобто, при збільшенні показника на 1 – 2 мм.рт.ст.) і ступінь гіпервентиляції є значною. Гіпоксія стимулює зовнішнє дихання тільки при значному зниженні Ро2 (до 50 – 60 мм.рт.ст.) і ступінь гіпервентиляції при цьому не значна. Тому головним стимулятором зовнішнього дихання є підвищення Рсо2 в артеріальній крові, а зниження Ро2 в фізіологічних умовах рідко виступає як стимулятор зовнішнього дихання (наприклад, при підйомі в гори), та часто стає ним в умовах патології.
111.Регуляция внешнего дыхания при физической нагрузке. При фізичному навантаженні розвивається гіпервентиляція, ступінь якої пропорційна інтенсивності навантаження. Регуляція зовнішнього дихання в цих умовах здійснюється як по збуренню (ведучий механізм) так і по відхиленню (в умовах дуже інтенсивного або довготривалого навантаження). Головним механізмом розвитку гіпервентиляції при фізичному навантаженні є безумовні рефлекси з пропріорецепторів працюючих м’язів (керуючий пристрій (КП), яким є дихальний центр, отримує по каналу зовнішнього зв’язку інформацію від пропріорецепторів працюючих м’язів про роботу що виконується; КП аналізує цю інформацію і викликає підвищення глибини та частоти дихання для того, щоб при збільшених метаболічних потребах тканин склад альвеолярного повітря залишався незмінним). Рецептори з пропріорецепторів працюючих м’язів є ведучим, та не єдиним механізмом, який стимулює розвиток гіпервентиляції при фізичному навантаженні. Ще до початку роботи гіпервентиляція може розвиватися за механізмом умовних рефлексів (передстартові стани у спортсменів). Від моторної зони кори ГМ паралельно з передачею команди про початок роботи до скелетних м’язів, інформація надходить і до нейронів дихального центру, що також сприяє розвитку гіпервентиляції. Гіпервентиляція, яка розвивається внаслідок вмикання всіх вищеназваних механізмів, рідко точно відповідає метаболічним потребам організму, частіше вона буває дещо надмірною або недостатньою (дихальному центру “важко” точно оцінити сигнали, які до нього надходять з точки зору інтенсивності м’язевої роботи, а значить і метаболізму). Якщо рівень вентиляції не зовсім відповідає рівню метаболізма, проходить зміна газового складу артеріальної крові. Ці відхилення сприймаються хеморецепторами (центральними та периферичними – СП1) передача інформації в КП каналом зворотнього зв’язку зміна вентиляції; вона стає відповідати рівню метаболізма нормалізація газового складу артеріальної крові. Таким чином, хеморецептори виступають як детектори похибки і вносять необхідні корективи (рефлекторно) в роботу дихального центру при фізичному навантаженні. І лише при роботі дуже інтенсивній чи довготривалій, коли система зовнішнього дихання не може забезпечити підтримання постійності газового складу в альвеолярному повітрі (і в артеріальній крові), рефлекси з хеморецепторів виступають як механізми стимуляції зовнішнього дихання. Пищеварение 112.Пищеварение в ротовой полости. Регуляция слюноотделения, жевания, глотания. Ротова порожнина – початковий відділ травної системи, виконує такі функції: 1.Механічна обробка їжі – подрібнення, перемішування. 2.Зволоження їжі слиною. 3.Хімічна обробка їжі відбувається за участю ферментів слини. 4.Знезараження – знищення мікроорганізмів лізоцимом слини. 5.Аналіз смакових якостей речовин смаковим аналізатором, на підставі аналізу формуються емоції відповідно до складу їжі (солодощі задоволення, лайно огида) й приймається рішення щодо продовження чи припинення її споживання; 6.Рефлекторне надходження інформації від рецепторів, які розміщуються в даній ділянці до інших органів системи травлення, що буде змінювати їх секреторну та моторну активність – це підготовка органів до перетравлення їжі з вже встановленим складом (багата на білок чи жири), що забезпечує більш ефективне травленя. 7.Всмоктування деяких низькомолекулярних речовин – це використовується для введення деякий лікарських препаратів – валідол. 8.Формування харчової грудки – порція їжі, що готова до ковтання. Жування – рефлекторний акт, який забезпечує перемішування, подрібнення та просочення їжі слиною до консистенці, що дозволяє відбутися ковтанню. Рефлекторна дуга цього рефлекса починається рецепторами слизової оболонки чутливі волокна трійчастого нерва центр жування (довгастий мозок) рухові волокна трійчастого нерва м’язи. Свідома регуляція ковтання відбувається завдяки зв’язку кори з центром жування. Ковтання – рефлекторний акт, який забезпечує перехід харчової грудки з ротової порожнини в нижні відділи глотки, а далі в порожнину стравоходу. Рефлекторна дуга цього рефлекса починається рецепторами кореня язика, піднебіння, задньої стінки глотки чутливі волокна язикоглоткового нерва центр ковтання рухові волокна язикоглоткового, під’язикового, трійчастого, блукаючого нервів м’язи ротової порожнини, язика, глотки та стравоходу.
113.Слюна – состав, свойства, функции. Механизм секреции слюны. Регуляция слюноотделения. Слина виділяється: І. Трьома парами великих слинних залоз: 1. Привушні (серозні). 2. Підщелепні (змішані). 3. Під’язикові (змішані). ІІ. Власними залозами слизової оболонки: 1. Серозні (на бокових поверхнях язика) 2.Слизові (корінь язика) 3.Змішані Слина – змішаний секрет всіх слинних залоз. Склад слини різних слинних залоз неоднаковий (привушні – серозна – найрідкіша, під’язикові – слизова – найгустіша). Змішана слина має 99,4 – 99,5% води, решта – органічні та неорганічні речовини, які забезпечують оптимум рН для дії ферментів. Органічні: - амілаза (секретується переважно привушними залозами), ліпаза слизової оболонки язика, кисла та лужна фосфатази, РНК-ази, ДНК-ази, муцин (глікопротеїн), захисні речовини (лізоцим, імуноглобуліни), поодинокі сперматозоїди; неорганічні – К+, Na+, Ca2+, Cl-, HCO3- та ін. В стані спокою рН слини складає 5,5-6,0, а при стимуляції підвищується до 7,8. Слина розчиняє речовини, що діють на смакові рецептори, зволожує ротову порожнину, змочує їжу, формує та покириває харчову грудку, сприяє ковтанню. Розпочинає гідроліз вуглеводів (- амілаза найбільш активна при рН 6,9 й розчеплює крохмаль до олігоцукридів (декстринів), мальтаза розчеплює дицукрид мальтозу до глюкози – тому, якщо потримати довго в роті ломтик хліба, відчувається солодкий присмак). Захисна функція полягає в тому, що в слині містяться бактерицидні речовини (лізоцим, імуноглобуліни тому собака зализує рани), вони сприяють заживленню мікротравм рота. При надходженні разом з харчовою грудкою в шлунок продовжує травлення в середині грудки та частково нейтралізує кислотність шлункового соку. Для дослідження слини у тварин вирізають слизову разом з протоком привушної залози й виводять її на зовнішню поверхню щоки. Коли рана загоїться, з цього протоку буде капати слина. Через цей свищ (фістула) збирають слину й досліджують її якість та склад при дії різних подразників. У людини слину збирають за допомогою капсули Лешлі-Красногорського. Її прикріплюють всередині щоки, де відкривається проток та збирають слину.
3. Механізми утворення слини, первинна та вторинна слина. В дольках (ацинусах) слинних залоз утворюється первинний секрет, який є ізотонічним (однаковим за йонним складом) по відношенню до плазми крові, і містить ферменти, що секретуються ацинарними клітинами. При проходженні цієї (первинної слини) через протоки, вона зазнає змін: більша частина натрію реабсорбується, обмінюючись на калій; хлор поглинається пасивно – в обмін на гідрокарбонат, до неї додається слиз, що секретується протоковими клітинами. Тому в кінцевій (вторинній) слині йонів Na+ та Cl- стає менше в 7-10 разів, а йонів К+ та HCO3- в 2-3 рази більше. Протоки майже непроникні для води, тому при проходженні через протоки слина стає гіпоосмічною. Електролітний склад слини істотно залежить від швидкості секреції – при високій швидкості концентрації натрію та хлору зростають, оскільки не встигає відбуватися реабсорбція. Утворення ферментів та слизу – активний процес, тобто відбувається з затратами енергії. Секреторний цикл – це послідовна зміна станів секреторних клітин, які виділяють ферменти та слиз. Для клітин, які виділяють ферменти можна визначити такі фази секреторного циклу: 1.Надходження з крові в секреторну клітину низькомолекулярних речовин, які необхідні для синтезу ферментів. 2.Синтез просекрету, іде на рибосомах, далі просекрет переходить в апарат Гольджі. 3.Дозрівання секрету в апараті Гольджі. 4.Упакування секрету в гранули. В цих гранулах секрет зберігається до тих пір, доки не надійде сигнал клітині про необхідність виділення секрету. 5.Через апікальний полюс відбувається виділення секрета (найчастіше це кальцій залежний процес). У людини слина виділяється постійно, а у тварин лише під час їжі.
4. Регуляція слинновиділення. Вплив властивостей подразника на кількість і якість слини. Слиновиділення має нервову та гуморальну регуляцію, проте основну роль у регуляції виконує нервова система. Слиновиділення знаходиться під контролем як симпатичної, так і парасимпатичної нервових систем. Індукують слиновиділення секреторні центри довгастого мозку, аферентні активуючі імпульси до якого надходять від рецепторів язика, ротової порожнини та піднебіння (смакові та тактильні), від носової порожнини (нюхові рецептори) та від вищих відділів мозку (думка про їжу). Від всіх цих рецепторів інформація про характер їжі надходить до довгастого мозку по V, VII, IX, X парах ЧМН, та від зорових та слухових рецепторів по відповідних нервах спочатку до вищих відділів мозку, а потім вже до центрів слиновиділення в довгастому мозку. Свої аферентні впливи парасимпатична НС здійснює за допомогою VII та IX пар ЧМН і тим самим ініціює виділення великої кількості слини з високим вмістом електролітів та з низьким вмістом білкових сполук (ферментів). На противагу їй, симпатична НС стимулює під’язикові та підщелепні слинні залози, які виділяють невелику кількість густої слини, багатої на білкові сполуки (саме тому деяки лектори носять з собою склянку води: погана поведінка студентів дратування викладача активація симпато-адреналової системи збільшення виділення густої слини (цей ефект усувається ковтком води) успішна здача державного іспиту студентами армія або КПІ). дуга парасимпатичного рефлексу дуга симпатичного рефлексу При відсутності стимуляції слинні залози секретують слину зі швидкістю 0,5 мл/хв. При появі певних стимулів (вигляд їжі, її запах, смак і навіть звук побрязкування кришки кастрюлі в якій вариться смачненький борщ), слиновиділення може зрости в декілька разів. Наприклад, при появі нюхового подразника слиновиділення збільшується вдвічі, при жуванні – в 2,5 рази, а деякі смакові стимулятори такі як лимонна кислота можуть збільшити швидкість слиновиділення до 7,4 мл/хв.
114.Состав и свойства желудочного сока. Механизмы секреции соляной к-ты и ее функции. На тваринах секреторну функцію шлунка досліджують такими методами: 1. Свищ (fistula) шлунка – трубка, що вставляється в порожнину шлунка. З зовнішнього боку свищ закривають пробкою і під час того, коли досліди не проводяться, тварина нормально живе і в її шлунку проходить нормальний процес травлення. Під час проведення досліду свищ відкривають, завдяки чому можна отримати шлунковий сік. Свищ шлунка можна комбінувати з езофаготомією (операцією по вирізанню стравоходу). У такої тварини можна проводити дослід “уявного годування”, який полягає в тому, що їжа, яку тварина (в основному для цього використовують собак) проковтує, не проникає в шлунок а виходить з отвору стравоходу. Завдяки цьому можна отримати чистий шлунковий сік без домішок їжі. Добутий таким чином шлунковий сік відноситься до того, який виділяється під час 1-ої фази шлункової секреції (бо їжа не подразнила слизову оболонку шлунка), тому в даному соці буде менше ферментів і більше електролітів. 2. Операції “маленьких шлуночків”. Даний метод полягає в тому, що в піддослідної тварини вирізають шматок тканини шлунка, з’єднують її із зовнішнім середовищем і пророблюють фістулу, утворюючи цим самим так званий “маленький шлуночок”. Є дві методики даних операцій: а) За Гейденгайном, вирізаючи частину шлунка та утворюючи з нього “маленький шлуночок”, перерізають при цьому гілки блукаючого нерва, залишаючи нормальним його кровопостачання (шлуночок стає денервованим). За допомогою цього методу можливо дослідити тільки гуморальну регуляцію шлункової секреції. б) За Павловим, утворюючи “маленький шлуночок”, залишали нормальним як його кровопостачання, так і інервацію. Саме завдяки цьому можна досліджувати і гуморальну, і нервову регуляцію шлункової секреції. У людини для дослідження шлункової секреції проводять такі дослідження: 1. Метод шлункового зондування полягає в введенні в порожнину шлунка зонд через який ми і отримуємо шлунковий сік для досліджень його якісного та кількісного складу. Найчастіше отримують 9 порцій шлункового соку через 15 хв. кожна. 1-у порцію отримують одразу ж після введення зонда в шлунок (ця порція береться вранці та натще). Потім, через одну годину збирають 4 порції соку в умовах базальної секреції. Потім, ще 4 порції в умовах стимуляції секреції шлунка. При цьому натщесерце отримують невелику кількість лужного соку; в умовах базальної секреції – багато кислого соку. В кожній порції визначають показники кислотності шлункового соку, його лужний компонент (наявність слизу), кількість і активність ферментів й на основі цих даних роблять висновок про секреторну діяльність шлунку. 2. Внутрішньошлункова ендоскопія – за допомогою введення в порожнину шлунка спеціальної оптичної техніки (ендоскопа), лікар наочно може вивчати стан слизової оболонки шлунка. 3. Метод ацидотеста на кислотність шлункового соку. Людині дають проковтнути спеціальну розчинну капсулу всередині якої знаходиться барвник, який при розчиненні капсули потрапляє на слизову шлунка, а потім в кров і виділяється з сечею. Далі збирають 3 порції сечі через 2-4-8 годин і визначають, яка порція буде більше зафарбована барвником. 4. Визначення активності пепсину в крові та сечі. Через те, що основна маса пепсину, виділяючись через апікальний полюс секреторної клітини, іде в просвіт залози, а її менша частина іде через базальний полюс в кров, то визначивши кількість пепсину в крові чи в сечі, можна дати оцінку ферментовидільній функції головних залоз шлунка.
Склад і властивості шлункового соку. В шлунку секреторну функцію виконують такі клітини: 1. Клітини шлункових залоз: - головні клітини – секретують ферменти; - парієтальні (обкладові) – соляну кислоту; - додаткові (мукоцити) – розчинний слиз. 2. Клітини покривного епітелію – нерозчинний слиз. До складу шлункового соку входять: 1. Ферменти: а) протеолітичні ферменти, які відносяться до ендопептидаз: - пепсин – гідролізує білки в дуже кислому середовищі (pH = 1,5-2); - гастриксин – розщеплює білки в менш кислому середовищі (pH = 3-3,5); Значення протеолітичних ферментів: В шлунку існує радіальний та повздовжній градієнт pH. Значення радіального градієнту заключається в тому, що чим ближче до стінки, тим pH нижчий (висока кислотність), а чим ближче до центру шлунка, тим він вищий (низька кислотність). Це явище пояснюється тим, що оскільки низьку pH забезпечує соляна кислота, а в їжі, що надходить до шлунку, завжди є лужні продукти які її нейтралізують. Саме тому, ближче до стінки білки перетравлюють пепсини, а далі від стінки – гастриксини. Повздовжній градієнт pH збільшується по напрямку до пілоричного відділу шлунку, так як залози цього відділу не виділяють соляну кислоту, але виділяють багато слизу, реакція якого лужна і тому він нейтралізує соляну кислоту, зв’язуючи йони водню. Через це, в області тіла та дна шлунку їжу перетравлюють пепсини, а в пілоричному відділі – гастриксини. Протеолітичні ферменти шлункового соку виділяються шлунковими залозами в неактивному стані. Активуються вони тільки в порожнині шлунка за рахунок соляної кислоти. Неактивна форма пепсину називається пепсиноген. Виділення ферментів в неактивному вигляді попереджує самоперетравлювання головних клітин залоз. б) ліпаза – розщеплює емульгований жир, який є в рідких молочних продуктах. Шлункова ліпаза забезпечує гідроліз цього жиру до жирних кислот та гліцерину. При цьому утворюються також дигліцериди та моногліцериди. Оптимум pH для дії шлункової ліпази = 3,3-5,4. В дорослої людини цей фермент не має великого значення для перетравлення їжі, але він є дуже важливим для дітей 1-го року життя. У маленьких дітей pH в шлунку = 3-4. Ферменти виділяють залози всіх відділів шлунка. 2. Соляна кислота, що являється одним із найважливіших компонентів шлункового соку, виділяється парієтальними клітинами, яких багато в залозах тіла та дна шлунка, та мало в залозах пілоричного відділу шлунка. Роль соляної кислоти в травленні: 1)забезпечує оптимальний рівень pH для активації та дії ферментів шлункового соку; 2)забезпечує набухання білків, що полегшує їх гідроліз; 3)викликає перетворення нормального молока на кисле, що полегшує його перетравлення ліпазою; 4)впливає на виділення багатьох гормонів ШКТ (гастрин, секретин, соматостатин); 5)впливає на рухову функцію шлунка; 6)руйнує патогенні мікроорганізми (антибактерицидна дія).
3. Слиз (розчинний та нерозчинний) Розчинний слиз виділяється додатковими клітинами залоз шлунка, змішується з їжею і полегшує її перетравлення. Нерозчинний слиз виділяється клітинами покрівного епітелію і покриває тонким шаром (близько 0,6 мм) всю слизову оболонку шлунка. Він виконує захисну функцію і захищає слизову шлунка від дії соляної кислоти та самоперетравлення протеолітичними ферментами.
Слиз має лужну реакцію (pH=7) і тому називається лужним компонентом шлункового соку. До складу слизу входить внутрішній фактор Касла – це глікопротеїд, який разом з R-білком виконує важливу роль у подальшому всмоктуванні вітаміну В12 в кишківнику. Механізм секреції HCl.
Гідрокарбонат через базолатеральні мембрани парієтальних клітин поступає в кров, де він входить до складу буферних систем крові, а замість нього з крові в клітину надходять йони Cl- (по градієнту концентрації). Йони H+ через апікальну мембрану цих же клітин, в якій знаходиться H+/K+-насос (АТФ-залежний процес), виходить в порожнину залози і далі в шлунок, а замість нього в клітину надходить K+. Шляхом активного транспорту через апікальну мембрану парієтальних клітин в порожнину шлунка виводяться йони Cl-, які надійшли з крові. Внаслідок всіх цих процесів в порожнину шлунка надходять йони H+ та Cl-, які і утворюють соляну кислоту.
115. Двигательная функция желудка и ее регуляция. Механизм перехода желудочного содержимого в ДПК. В регуляції моторики шлунку беруть участь нервові та гуморальні механізми. Серед нервових механізмів виділяють місцеві та центральні. Деякі м’язові клітини внутрішнього шару м’язової оболонки шлунку мають пейсмейкерну активність, тобто періодично генерують ПД – з частотою 3-2 на секунду, що спричиняє періодичне підвищення внутрішньошлункового тиску. Впливи парасимпатичної нервової системи, котрі досягають шлунку волокнами блукаючого нерву, теж стимулюють моторику. Через деякий час після їди шлунок починає скорочуватися. У процесі таких скорочень, що повторюються з інтервалом близько 20 с, їжа, що знаходилась біля стінок, пересувається до антрального відділу – таким чином здійснюється її перемішування. Протягом першої години після вживання їжі перистальтичні хвилі слабкі, потім посилюються, при цьому швидше просувається та частина їжі, що містилася біля стінок. Гуморальні стимулятори шлункової моторики ХЦК-ПЗ, гастрин, мотилін, інсулін серотонін, ацетилхолін. Інгібітори секретин, ВІП, ШІП. Перехід хімусу з шлунку до дванадцятипалої кишки визначається, в основному градієнтом тиску між ними. Також при надходженні до duodenum кислого хімусу, продуктів гідролізу жирів стимулюється секреція інгібіторів шлункової моторики. Посилення моторики починається з впливу n.vagus, потім до нього приєднуються гуморальні механізми, після виходу хімусу, припиняється подразнення нервових закінчень, потім зникають гуморальні впливи – таким чином відновлюється рівень базальної секреції.
116.Фазы регуляции желудочной секреции. Энтеральные стимуляторы и ингибиторы желудочной секреции. Кількість та склад шлункового соку змінюється особливо після вживання їжі. В значній мірі кількість та склад соку залежить від характеру подразника (кількість та склад їжі). Натще секретується невелика кількість шлункового соку (до 10 мл на годину). Він містить в основному слиз. Після прийому їжі виділення шлункового соку значно збільшується, росте його кислотність та вміст ферментів. За головними механізмами регуляції шлункову секрецію поділяють на декілька фаз: І фаза – складно рефлекторна. Секреція в цій фазі пов’язана з реалізацією умовних рефлексів (на вигляд, запах їжі, думка про якусь смакоту). Другий механізм – безумовні рефлекси, що виникають при подразненні їжею рецепторів ротової порожнини блукаючий нерв вплив на головні та G-клітини. Особливості секреції в першій фазі (вивчають ці особливості при постановці досліду “уявного годування”): 1. Короткий латентний період (5-8 хвилин). Після початку досліду перша крапля соку з свища з’явиться через 5-8 хвилин. Цей період короткий тому, що секреція викликана нервовими механізмами регуляції. 2. Кількість соку невелика – 20% від всього об’єму, який виділяється під час їди. 3. Склад соку – в соці багато ферментів тому, що блукаючий нерв передає інформацію до головних клітин шлунка. Багато ферментів швидке переварювання білків їжі. 4. Тривалість секреції невелика – 1-1,5 години тому, що нервові механізми регуляції. ІІ фаза – нейрогуморальна. В цій фазі секреція пов’язана з реалізацією нервових та гуморальних механізмів регуляції. Нервові – це рефлекси. Гуморальні – шлунково-кишкові гормони гастроінтестинальні гормони). Реалізується цей механізм таким чином: при розтягненні шлунку подразнюються чутливі волокна блукаючого нерва, ця інформація надходить в довгастий мозок, і парасимпатичними волокнами блукаючого нерва назад до шлунку, стимулюючи секрецію – ваго-вагальний рефлекс; в нервовій регуляції також беруть участь інтрамуральні нервові сплетення. Хімічна стимуляція секреції здійснюється посередництвом гастрину, що виділяється G-клітинами. Гастрин з током крові надходить до парієтальних клітин – стимулює виділення соку з високою концентрацією соляної кислоти. Основні стимулятори секреції гастрину – продукти переварювання білків – пептиди, олігопептиди, амінокислоти – особливо триптофан і фенілаланін, а також кальцій, магній, алклголь та кофеїн. При надходженні хімусу до дванадцятипалої кишки, здійснюється дуоденогастральний рефлекс – продукти гідролізу поживних речовин, соляна кислота, а також розтягнення дванадцятипалої кишки хімусом викликають підвищення секреції тканинних гормонів – секретину S-клітинами, котрий гальмує шлункову секрецію, і холецистокініну-панкреозиміну І-клітинами, що має подібну дію. Стимулятори шлункової секреції: -ацетилхолін; -гастрин; -гістамін; -бомбезин; -інсулін; Блокатори шлункової секреції: - вагогастрон виділяється вагусними закінченнями; - секретин; - глюкагон - -клітини pancreas; - вазоактивний інтестинальний пептид – ВІП – метасимпатична нервова система; - гастроінгібуючий пептид – ГІП – D-клітини кишково-шлункового тракту; - нейротензин – N-клітини тонкого кишківника; - соматостатин - -клітини pancreas; - вазопрасин, окситицин; - простагландини; - катехоламіни.
117.Секретоная функция поджелудочной ж-зы, ее регуляция. Состав и функции поджелудочного сока. Серед методів дослідження зовнішньосекреторної функції підшлункової залози виділяють гострі та хронічні. У людини цю функцію досліджують такими методами: -зондування; -УЗД; -томографія; -визначення ферментів у крові та сечі; -копрографія. Склад та властивості підшлункового соку. У людини виділяється 1,5 – 2 літри соку на добу. Кислотність його – 7,8 – 8,4. Електроліти соку. Основні аніони підшлункового соку – Cl- та HCO3-, катіони – Na+ та K+. На відміну від слини, сік ізотонічний плазмі крові незалежно від ступеня стимуляції. Концентрація катіонів при стимуляції лишається сталою, аніонів – змінюється в протилежному напрямку. Карбонат утворюється в ацинусах у більш високій концентрації, а при проходженні через протоки частково обмінюється на хлор – тому при посиленні підшлункової секреції концентрація карбонату підвищується не встигає відбуватися обмін йонів. Окрім цих йонів, в підшлунковому соці містяться також йони кальцію, магнію, цинку, сульфат та фосфат. Ферменти підшлункового соку: 1.Протеолітичні аендопептидази – розщеплюють внутрішні пептидні зв’язки у білковій молекулі – трипсин, хемотрипсин, еластаза. бекзопептидази – відщеплюють кінцеві амінокислоти: карбоксипептидази відщеплює амінокислоти від СООН-кінця, амінопептидази відщеплюють амінокислоти від NH2 - кінця 2.Амілолітичні: -амілаза – розщеплює -1,4-глікозидні зв’язки в полімерах глюкози. 3.Ліполітичні: ліпаза – розщеплює ефірні звязки в положеннях 1 і 3 тригліцеридів, фосфоліпаза А2 – ефірні зв’язки в положенні 2 фосфоліпідів, холестеролаза – ефіри холестеролу. 4.Нуклеолітичні: рибонуклеаза – фосфодиефірні зв’язки між сусідніми нуклеотидами в рибонуклеїнових кислотах. Фази регуляції секреторної функції підшлункової залози. В регуляції секреції підшлункової залози виділяють 2 фази: Перша фаза – складнорефлекторна (цефалічна). Секреція підшлункового соку під час цієї фази пов’язана з реалізацією складнорефлекторних механізмів регуляції, тобто умовних рефлексів (вигляд, запах їжі) і з реалізацією безумовних рефлексів (подразнення їжею рецепторів ротової порожнини). Для цієї фази характерні такі показники: 1.Короткий латентний період (1-2хв.); 2.Об’єм соку невеликий (20% від загального об’єму соку, який виділяється при їді); 3.Сік містить багато ферментів і має високу перетравлюючу силу; 4.Невелика тривалість секреції (1-2год.); 5. Переважають нервові механізми регуляції. Друга фаза – нейрогуморальна (кишкова). Ця фаза називається нейрогуморальною, тому що вона зв’язана з реалізацією вагальних безумовних рефлексів у відповідь на подразнення їжею чи хімусом механо- та хеморецепторів слизової оболонки шлунка та тонкого кишківника. Метасимпатичні рефлекси під час цієї фази мають менше значення, ніж в регуляції шлункової секреції. Схема нервової регуляції під час 2-ї фази: Для реалізації 2-ї фази підшлункової секреції великого значення набувають гуморальні механізми регуляції (за допомогою шлунково-кишкових гормонів). На початкових етапах травлення виділяються гормони, які стимулюють підшлункову секрецію, а на пізніх етапах навпаки – які її гальмують. Гормони, що стимулюють підшлункову секрецію: 1. Секретин. Виділяється S-клітинами стінки дванадцятипалої кишки під впливом соляної кислоти, яка надійшла з шлунку разом з хімусом. Він викликає виділення великої кількості підшлункового соку, багатого бікарбонатами, але бідного на ферменти, так як основна його дія спрямована на протокові клітини, що виділяють бікарбонати і він майже не діє на клітини ацинусів, що секретують ферменти. 2. Холецистокінін-панкреозимін (ХЦК-ПЗ). Виділяється І-клітинами стінки дванадцятипалої кишки під впливом продуктів початкового гідролізу харчового білка та жиру, деякі амінокислоти, а також соляна кислота та вуглеводи. Він діє на ацинозні клітини, а тому стимулює виділення невеликої кількості підшлункового соку багатого на ферменти. Виділяючись одночасно ХЦК-ПЗ та секретин посилюють дію один одного. ХЦК-ПЗ також стимулює скорочення жовчного міхура та вихід жовчі в дванадцятипалу кишку. 3. Гастрин. Виділяється G-клітинами шлунка та проксимального відділу тонкої кишки. Окрім посилення секреції підшлункової залози, гастрин також посилює перистальтику тонкого кишківника та жовчного міхура. 4. Бомбезин. Виділяється Р-клітинами шлунка та проксимальних відділів тонкого кишківника. Стимулює секрецію ферментів підшлунковою залозою через стимуляцію вивільнення ХЦК-ПЗ. 5. Субстанція Р. 6. Серотонін. 7. Інсулін. Гормони, що гальмують підшлункову секрецію: 1. Соматостатин. Виділяється D-клітинами дванадцятипалої кишки та паренхіми підшлункової залози за умови підвищення вмісту поживних речовин в крові. Спричиняє зниження виділення підшлункового соку через гальмування секреції шлунково-кишкових гормонів. 2. Нейротензин. Виділяється N-клітинами дистальних відділів тонкої кишки. Гальмує виділення соляної кислоти шлунком зменшення, або повне припинення секреції секретину та ХЦК-ПЗ відсутність стимулюючих гуморальних впливів на секреторні клітини підшлункової залози пониження її секреторної діяльності. Посилює виділення глюкагону. 3. Глюкагон. 4. Енкефаліни. 5. Шлунковий інгібуючий пептид (ШІП). 6. Кальцитонін. Характеристика секреції в 2-їй фазі: 1.латентний період довший, ніж в першій фазі; 2.об’єм соку = 80% від загального об’єму соку, що виділяється під час їжі; 3.до складу соку входить багато ферментів, води та бікарбонатів; 4.довготривалість секреції (декілька годин).
118.Состав и функции желчи. Желчные кислоты, их виды и функции. Желчные пигленты. У людини жовчовиділеня досліджується за допомогою таких методів: 1) Дуоденальне зондування. Зонд вводять в дванадцятипалу кишку і збирають її вміст. Для збільшення надходження жовчі в дуоденум, підшкірно можна вводити гормон ХЦК-ПЗ, або через зонд – концентрований розчин MgSO2, яєчний жовток чи глюкозу, які стимулюють секрецію та виділення жовчі. В жовчі досліджують вміст жовчних кислот та інших компонентів. 2) Дослідження вмісту в крові, сечі та калі жовчних пігментів (білірубіну, білівербину та стеркобіліну). Даний метод базується на тому, що у випадку порушення функції виділення жовчі, в крові та в сечі концентрація жовчних пігментів підвищується, а в калі навпаки – понижується.
3) Ультразвукове дослідження (УЗД) печінки та жовчного міхура. Цей метод дозволяє вивчити секреторну функцію печінки та евакуацію жовчі із жовчного міхура. Состав желчи Жовч має такі властивості: 1. Емульгує жири, збільшуючи поверхню, на якій здійснюється їх гідроліз ліпазою; 2. Розчиняє продукти гідролізу жирів, чим сприяє їх гідролізу; 3. Підвищує активність панкреатичних та кишкових ферментів, особливо ліпази; 4. За участю жовчних солей проходить утворення тонкодисперсних частинок жирів, внаслідок чого вони в невеликій кількості можуть всмоктуватись із тонкої кишки навіть без проходження попереднього гідролізу; 5. Стимулює моторну та секреторну діяльність тонкої кишки; 6. Здатна припинити шлункове травлення не тільки нейтралізацією соляної кислоти, яка поступила в дванадцятипалу кишку, а й шляхом інактивації пепсину; 7. Має бактеріостатичні властивості; 8. Приймає участь у процесах всмоктування з кишківника жиророзчинних вітамінів, холестерину, амінокислот та солей кальцію.
119.Регуляция образования и выделения желчи. Влияние состава пищи на желчеобразование и желчевыведение. Механізм жовчоутворення та жовчовиділення: Утворення жовчі іде постійно, але збільшується під час травлення, під впливом складно-рефлекторних механізмів, які відносяться до 1-ї фази жовчоутворення (вигляд, запах їжі, звуки, що супроводжують їду), а також нейрогуморальних впливів, які діють під час 2-ї фази жовчоутворення та жовчовиділення. До нервових впливів відноситься стимулюючий вплив парасимпатичної нервової системи (Vagus), а до гуморальних – стимуляція жовчоутворення та жовчовиділення такими гормонами як гастрин, секретин, ХЦК-ПЗ та глюкагон. Механізм надходження жовчі в дванадцятипалу кишку: Вихід жовчі з жовчного міхура, та її рух по жовчовивідних шляхах зумовлений різницею тисків в вищезгаданих частинах, та в дванадцятипалій кишці. По-за процесу травлення, тиск в жовчному міхурі становить 60 – 185 мм.вод.ст., а під час травлення, за рахунок скорочення стінок жовчного міхура, тиск в ньому піднімається до 150 – 260 мм.вод.ст., забезпечуючи вихід жовчі в дванадцятипалу кишку по градієнту тисків (за даних умов тиск в дванадцятипалій кишці стає меншим) через сфінктер Одді, який в цей момент відкривається. Такі зміни стану жовчного міхура та сфінктера викликають парасимпатичні рефлекси (умовні та безумовні) і гормон ХЦК-ПЗ.
120.Секреторные функции тонкой кишки. Состав и свойства кишечного сока. Регуляция его секреции. Полостное и пристеночное пищеварение. За добу виділяється близько 1,8 л. кишкового соку (КС), pH якого приблизно рівний 7,8 – 8,0. В його секреції приймають участь Бруннерові залози, які розміщені в дванадцятипалій кишці і виділяють багато слизу, а також Ліберк’юнові крипти, в яких розміщені різні за будовою та функціями епітеліальні клітини (слизові, недиференційовані ентероцити, ендокринні, клітини Панета). За рахунок складок слизової оболонки кишки (складки Керкрінга), ворсинок, крипт та мікроворсинок площа поверхні слизової оболонки тонкого кишківника досягає 200 м2. Після центрифугування КС видно його поділ на рідку та щільну частини: 1. Рідка частина містить в собі воду та електроліти (хлориди, бікарбонати і фосфати натрію, калію, кальцію). В ній майже відсутні ферменти; 2. Щільна частина вміщує близько 22-ох ферментів, які завершують гідроліз пептидів до амінокислот; жирів – до гліцерину та жирних кислот; вуглеводів – до моноцукрів. В цій частині КС наявні також злущені клітини епітелію. Ферменти кишкового соку: 1) Протеолітичні: - поліпептидази (лейцинамінопептидаза) – розщеплюють пептиди різної довжини; - трипептидази (амінополіпептидаза); - дипептидази; - катепсини (розщеплюють нуклеїнові кислоти, нуклео-зиди). 2) Амілолітичні: - лактаза; - сахараза; - мальтаза; - амілаза; - інвертаза (розщеплює сахарозу до глюкози та фруктози). 3) Ліполітичні: - лужна фосфатаза – розщеплює фосфорні ефіри; - ліпаза; - фосфоліпаза; - холестеролестераза. Функції кишкового соку: а) кінцевий гідроліз харчових продуктів; б) захист слизової оболонки кишки від механічних та хімічних пошкоджень; в) підтримання рідкого стану хімусу; г) участь в олужненні кислого хімусу. Регуляція секреції кишкового соку: 1. Місцеві механізми регуляції здійснюються за допомогою метасимпатичної нервової системи, яка реагує на тактильні та хімічні стимули, що ідуть від хімусу, який і подразнює слизову оболонку кишки вмикаючи цим метасимпатичні рефлекси підсилення секреції КС. 2. Нервові механізми регуляції здійснюються за посередництвом пара-симпатичних (стимулюють) та симпатичних (гальмують) нервів. 3. Гуморальні механізми регуляції здійснюються за допомогою стимулю-ючих (секретин, ентерокіназа, ВІП, ентерокінін) та пригнічуючих (серотонін) секрецію КС гормонів. Поняття про порожнинне та пристінкове травлення: Порожнинне травлення проходить в порожнині кишкового каналу за рахунок ферментів. Це гідроліз поживних речовин, який іде в порожнині в: 1. Розчині, де контактують розчинені ферменти та розчинені субстрати; 2. На границі поділу фаз – це поверхні твердих частинок їжі, волокна сполучної тканини, згустки слизу. Порожнинне травлення може забезпечити гідроліз до кінцевих продуктів, але його тривалість дуже велика. Пристінкове травлення проходить на мембрані глікокалікса мікроворсинок ентероцитів за допомогою фіксованих ферментів, активні центри яких направлені на субстрат. Під час цього виду травлення дуже швидко проходить гідроліз близько 80% вуглеводів, 60% жирів та пептонів. Перевагою цього травлення є також його стерильність (структура ворсинок та глікокалікса не дає можливості мікроорганізмам пройти в кров та в лімфу).
121.Двигательная функция кишечника, методы ее изучения. Виды сокращений и их регуляция. Механизмы перехода химуса из тонкой кишки в толстую. Рух тонкої та товстої кишок принципово не відрізняються, хоча рухи товстої кишки складніші, так як в ній хімус знаходиться протягом більш тривалого часу. За механізмами виникнення всі види скорочень гладких м’язів кишківника і всі його рухи поділяються на 2 групи: 1. Міогенні – м’язові види скорочень та рухів. В їх основі лежать особливі властивості гладких м’язів товстої та тонкої кишок: а) Здатність до автоматії – здатність збуджуватись без зовнішніх подразників. Це пояснюється наявністю в кишківнику двох видів пейсмекерів (збудників ритму) – це місце, де знаходиться багато нервових клітин, що здатні до автоматії. Один знаходиться в дванадцятипалій кишці, другий – посередині довжини товстого кишківника. Саме в цих місцях виникають м’язеві скорочення, що рухаються вздовж кишки в дистальному напрямку. б) Наявність з’єднань між клітинами, через які ПД поширюється з одної клітини на іншу. Види міогенних скорочень: 1. Тонічні – зумовлені тонусом гладких м’язів – тривалі скорочення невеликої сили. Тонічні скорочення тривають близько 10 хв, такі скорочення займають великі ділянки кишки. За рахунок тонусу зростає внутрішньокишковий тиск, що покращує всмоктування і контакт хімусу та стінок кишки. 2. Ритмічна сегментація – періодичні скорочення циркулярного шару м’язової оболонки в певних ділянках кишки. В результаті кишка та її вміст ділиться на сегменти. Потім ці м’язи розслаблюються, інші скорочуються – виникають інші сегменти. В результаті здійснюється перемішування хімусу і покращуються умови його контакту зі стінкою кишки.
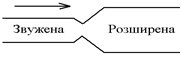
3. Маятникоподібні скорочення – періодичні скорочення поздовжнього шару м’язової оболонки. Просвіт кишки при цьму стає ширшим, а сама кишка – коротшою. Допоміжне значення мають скорочення м’язів циркулярного шару. При таких скороченнях хімус в просвіті кишки виконує коливальні рухи, тобто йде перемішування хімуса, що покращує пристінкове травлення. Послідовність розвитку міогенних скорочень така: 1. Тонічні скорочення; 2. Ритмічна сегментація; 3. Маятникоподібні скорочення. Ці види скорочень змінюються при зміні кількості хімусу в кишківнику і його параметрів. Наприклад, розтягнення кишківника хімусом посилює ці скорочення. 2. Нейрогенні. В основі регуляції цих рухів лежать рефлекси: 1. Перистальтика – рух вздовж кишки звуженої ділянки, а перед нею – розширеної. Даний вид скорочень забезпечує рух хімуса вздовж кишки в дистального напрямку.
Подразники, що стимулюють метасимпатичний рефлекс: 1) Механічні: а) розтягнення кишки хімусом; б) подразнення стінки кишки грубою їжею, рослинною клітковиною (овочі, фрукти, чорний хліб). 2) Хімічні: а) нейтральні рідкі жири – це всі види рослинних масел. Вони стимулюють перистальтичний рефлекс; б) молочнокислі продукти; в) екстрактивні речовини (соки, бульйони); г) жовчні кислоти. Рухову функцію кишківника також посилюють гастрин, гістамін, серотонін. Кініни, простагландини посилюють цю функцію в патології. В звичайних умовах центральні вегетативні рефлекси (симпатичні та парасимпатичні) не впливають на перистальтику, але в умовах різкого збудження цих відділів ВНС, вони можуть дещо вплинути на перистальтику (як правило парасимпатичні посилюють перистальтику а симпатичні – гальмують).
122. Механизмы и регуляция всасывания белков, жиров, жирорастворимых витаминов и углеводов в различных отделах ЖКТ. Всмоктування – це процес транспорту речовин із порожнини травного каналу у внутрішні середовища організму (кров та лімфу). Більшість речовин всмоктується в кров (продукти гідролізу білків, вуглеводів та вода), в лімфу всмоктуються продукти гідролізу ліпідів. Найінтенсивніше процеси всмоктування проходять в верхніх відділах тонкого кишківника. Всмоктування в шлунку. Об’єм всмоктування на даному відрізку травного каналу дуже малий. Тут всмоктуються моносахариди, деякі солі, вода та алкоголь. Всмоктування в тонкому кишківнику. Здатність до всмоктування практично однакова у всіх відділах тонкого кишківника. У здорової людини всмоктування поживних речовин, солей та води проходить в верхніх відділах тонкого кишківника, а нижні відділи являються резервом всмоктування. У дорослої людини при нормальному харчуванні за добу всмоктується приблизно 100 г білків у вигляді амінокислот, 100 г жирів у вигляді гліцерину та жирних кислот, 500 г вуглеводів у вигляді моносахаридів. При великих навантаженнях може всмоктуватись до 500 г білків, до 100 г жирів та до 4000-5000 г вуглеводів. Загальні механізми всмоктування: Переважають активні механізми транспорту (з затратами енергії проти градієнта концентрації). Так всмоктуються багато йонів, більшість амінокислот та моносахаридів. Існує і пасивний транспорт. Так в епітеліоцити з кишківника надходять жирні кислоти. Вода всмоктується також пасивно за механізмом осмосу, або за допомогою ультрафільтрації. Покращують всмоктування такі умови: - pH=7,4 - осмотичний тиск, що = 7,6 Атм. - гідростатичний тиск хімусу, що = 6 – 8 см.вод.ст. Всмоктування проводять ворсинки, до складу яких входять кровоносні судини та лімфатичні капіляри. До складу ворсинок також входять гладком’язеві клітини, які при всмоктуванні ритмічно скорочуються і цим викликають періодичне зменшення чи збільшення об’єму та площі поверхні ворсинок, що посилює всмоктування. Механізм всмоктування йонів Na+:
Механізм всмоктування глюкози: Вуглеводи всмоктуються тільки у вигляді моноцукрів, переважно за механізмом вторинного активного транспорту в комплексі з йонами Na. Na- насос з затратами енергії АТФ створює градієнт концентрації йонів Na. На апікальній мембрані є білки-переносники, які мають 2 активних центри. Один для зв’язування йонів Na, другий – для зв’язування моноцукрів (наприклад, глюкози). Комплекс білок-переносник – йон Na – глюкоза, рухається до внутрішньої поверхні мембрани клітини, цей рух викликає градієнт концентрації йонів Na в клітині та в порожнині кишки (цей градієнт створюється за допомогою Na-го насосу про дію якого було згадано вище). На внутрішній поверхні мембран клітин комплекс розпадається і в цитоплазму надходять йони Na та глюкоза. Далі йони Na видаляються із клітини Na-насосом, а глюкоза переходить в кров пасивно за механізмом дифузії. Білок-переносник стає вільним і цикл повторюється знову. Механізм всмоктування білків: Білки всмоктуються переважно у вигляді амінокислот (АК) за механізмом активного транспорту разом з йонами Na. Виділяють 5 білків-переносників для різних АК. Невелика частина білків проходить в епітеліоцити у вигляді поліпептидів за механізмом третинного активного транспорту також в комплексі з йонами Na. В епітеліоцитах ці поліпептиди гідролізуються до АК, які далі пасивно надходять в кров. У маленьких дітей всмоктування можливе за механізмом піноцитоза (мікровезикулярний транспорт), який має для дітей велике значення, так як забезпечує надходження в організм дитини продуктів гідролізу молока. Механізм всмоктування жирів має такі особливості: 1) жири всмоктуються переважно у вигляді жирних кислот та гліцерину, хоча можуть всмоктуватись і моногліцериди; 2) жирні кислоти з довгими ланцюгами і гліцерин всмоктуються тільки в комплексі з жовчними кислотами; 3) оскільки жири погано розчиняються в воді, то вони транспорту-ються до епітеліоцитів в комплексі з жовчними кислотами. Жовчні кислоти разом з ліпідами утворюють міцели циліндричної форми. Всередині міцели розміщується жирна кислота. Міцели вільно переміщуються в воді і підходять до мембран епітеліоцитів. Тут міцели розпадаються на жирні кислоти та гліцерин, які надходять в клітину пасивно за механізмом дифузії; 4) в епітеліоцитах проходить ресинтез нейтральних жирів (три-гліцеридів), які характерні для даного організму з жирних кислот та гліцерину, що надійшли з їжею; 5) синтезований нейтральний жир в епітеліоцитах з’єднується з білками утворюються хіломікрони, які збільшують водорозчинність жиру; 6) хіломікрони транспортуються переважно в лімфу.
123.Согласование различных этапов пищеварения и его нейро- гуморальные механизмы. Подразники, що стимулюють метасимпатичний рефлекс: 1) Механічні: а) розтягнення кишки хімусом; б) подразнення стінки кишки грубою їжею, рослинною клітковиною (овочі, фрукти, чорний хліб). 2) Хімічні: а) нейтральні рідкі жири – це всі види рослинних масел. Вони стимулюють перистальтичний рефлекс; б) молочнокислі продукти; в) екстрактивні речовини (соки, бульйони); г) жовчні кислоти. Рухову функцію кишківника також посилюють гастрин, гістамін, серотонін. Кініни, простагландини посилюють цю функцію в патології. В звичайних умовах центральні вегетативні рефлекси (симпатичні та парасимпатичні) не впливають на перистальтику, але в умовах різкого збудження цих відділів ВНС, вони можуть дещо вплинути на перистальтику (як правило парасимпатичні посилюють перистальтику а симпатичні – гальмують).
Обмен вещ-в, терморегуляция, выделение 124. Источники и пути использования энергии в организме. Методы опредления энергозатрат организмом человека. Дыхательный коэффициент. Энергия тратится в организме на поддержание и обеспечение целосности клеточных структур, поддержание ионных градиентов, на биосинтетические процессы, обеспечение специальных форм клеточной активности (сокращение, проведение нервных импульсов, секреция).Часть энергии выделяется в виде тепла(обогрев), ж(9,3ккал),б,у(4,1ккал). Методы определения энергозатрат: - прямые(камера Этуотера - Бенедикта) - Непрямые Дихательный коэффициент – соотношение обьема выделенного СО2 к обьему потребленного О2. (окисление 1г белка ДК=0.8, жира=0.7, углеводов=1, при употреблении смешанной пищи = 0.85-0.9) По таблицам Гарриса и Бенедикта,
125.Основной обмен и методы его определения. Факторы что влияют на его величину. Основной обмен минимальный расход энергии, обеспечивающий гомеостазис в стандартных условиях: при бодрствовании, максимальном мышечном и эмоциональном покое, натощак (12 – 16 часов без еды), при температуре комфорта (18 – 20С). энергия основного обмена расходуется на обеспечение жизнедеятельности всех органов и тканей организма, клеточный синтез, на поддержание температуры тела. Определяют в стандартных условиях:максимального мышечного и эмоционального покоя, натощак, при температуре комфорта Потому что физическая нагрузка, эмоциональное напряжение, прием пищи и изменение температуры окружающей среды увеличивают интенсивность метаболических процессов в организме (расход энергии). Определяют по таблицам, по формулам, по номограммам. Метод Крога (неполный газовый анализ). У мужчин 1500 – 1700 ккал (6300 – 7140 кДж), или 21 – 24 ккал (88 – 101 кДж)/кг/сутки. У женщин примерно на 10% меньше этой величины. Факторы определяющие величину должного основного обмена Пол, возраст, рост и масса тела (вес). Условия жизнедеятельности, к которым организм адаптирован: постоянное проживание в холодной климатической зоне увеличивает основной обмен; длительное вегетарианское питание – уменьшает
126.Физиологические принципы составления пищевого рациона. 1 соотношение Б:Ж:У = 1:1:4 2 в составе белковой пищи не мение 30% белков должны быть животного происхождения. 3 в составе жиров не мение 30% должны быть растительног происхождения. 4 в сутки в организм должно поступать оптимальное количество минеральных вещ-в, 5 -|-|-|- витаминов, 6 -|-|-|- клетчатки, 7 -|-|-|- воды, 8 питание должно бить дробным (завтрак 25%, 2завтрак/полдник 15%, обед 35%, ужин 25%), 9 пища должна обладать удовлетворительными органолептичискими свойствами, 10 в рацион должны входить продукты соответствующие национальным традициям.
127.Температура тела человека. Физиологическое значение гомойотермии. Терморегуляция и центр терморегуляции. Пойкилотермные животные (холоднокровные) – с непостоянной температурой тела, зависящей от температуры окружающей среды; гомойотермные (теплокровные) – животные с постоянной температурой тела, не зависящей от температуры окружающей среды. Значение гомойотермии обеспечение высокого уровня жизнедеятельности относительно независимо от температуры окружающей среды. Основная выроботка теплп идет в мышцах, легких, печени, почках.Центром терморегуляции является Гипоталамус.
128.Механизмы физической терморегуляци. Тепловая адаптация. физическая терморегуляция – с помощью изменения интенсивности теплоотдачи. Теплоизлучение (радиация), теплоиспарение, теплопроведение, конвекция При повышении температуры окружающей среды сосуды кожи расширяются. Смысл в том, что изменение ширины просвета сосудов(расширяются), регулируя теплоотдачу, способствует поддержанию постоянной температуры тела.
129. Механизмы химической терморегуляци. Холодовая адаптация. Химическая терморегуляция – регуляция температуры тела с помощью изменения интенсивности теплопродукции. При понижении температуры сосуды кожи суживаются Смысл в том, что изменение ширины просвета сосудов(суживаются), регулируя теплоотдачу, способствует поддержанию постоянной температуры тела.
130.Общая характеристика системы выделения. Функции почек, как основного органа этой системы. Особенности кровоснабжения почек. Головним виконавчим органом системи виділення є нирки, бо лише за їх участі можливе підтримання параметрів гомеостазу. Всі інші виконавчі органи відіграють допоміжну роль і виводять з організму: -легені – леткі (газоподібні) речовини: СО2, ацетонові тіла, пари алкоголю, ефіру; -шкіра та її залози – продукти азотистого обміну, деякі іони; об’єм виділення невеликий, але стає більш значним при порушенні видільної функції нирок; -травний канал та його залози – солі важких металів (при отруєнні ними), солі йоду (при їх надмірному вмісті в організмі), продукти азотистого обміну (їх виділення стає суттєвим при порушенні видільної функції нирок). Нирки являються основним органом системи виділення, так як тільки він виділяючи з організму в великій кількості продукти азотистого обміну, підтримують їх концентрацію в крові на певному рівні. Участь в цьому процесі шкіри, травного каналу та їх залоз недостатньо. Саме тому, при недостатності видільної функції нирок організм страждає від уремії – підвищеної концентрації в крові продуктів азотистого обміну. Тільки нирки можуть довго і в самих різноманітних умовах підтримувати параметри водно-сольового гомеостазу організму. Особливості кровопостачання нирок: 1.Дуже високий рівень кровотоку навіть в стані спокою – 1250 мл/хв (тобто, 25% ХОК, а маса нирок складає 0,43% від маси тіла). Це пов’язане з тим, що кровотік в нирках йде не тільки на живлення нирок як органа, а й на виконання ними видільної функції; 2.Кровотік в нирках нерівномірний – 92,5% крові направляється в кіркову речовину, де в ниркових клубочках проходить процес фільтрації; 3.Ниркова артерія відходить безпосередньо від аорти, приносні артеріоли (vas afferens) ширші від виносних (vas efferens), що створює в капілярах ниркового клубочка досить високи тиск (60 – 70 мм.рт.ст.); 4.В нирках існує чудова сітка капілярів – виносна артеріола знову розпадається на сітку капілярів, які оплітають канальці нефрона. Паралельно з петлями Генле проходять прямі судини (довгі прямі капіляри). Тиск крові в вторинній сітці капілярів становить 15 – 20 мм.рт.ст.; 5.Нирковий кровотік характеризується відносною постійністю та незалежністю від величини системного артеріального тиску (точніше від змін САТ). В значному ступені це являється наслідком хорошої вираженості в судинах нирок міогенних мехпнізмів регуляції. 6.Механізми регуляції можуть змінювати величину ниркового кровотоку через зміну тонусу приносної та виносної артеріол (тобто, змінюючи співвідношення пре- та посткапілярного опору). В основі процесу сечоутворення лежать три процеси: -фільтрація в нирковому тільці – в результаті цього процесу утворюється первинна сеча (близько 180 л/добу); -реабсорбція речовин; -секреція речовин. Функции почек: 1 водный баланс организма, 2 ионный баланс, 3 постоянство осмотического давления, 4 кислотно-основного баланса, 5 метаболизм Б, Ж, У, нуклеиновых кислот, 6 обмен электролитов, 7 эритропоез(эритропоэтин), 8 гемостаз (урокиназа, тромбопластин, гепарин), 9 защитная, 10 регуляция величины АД.
131.Клубочковая фильтрация а почках, механизм, регуляция. Состав первичной мочи. Сечоутворення починається з процесу клубочкової фільтрації, яка проходить в ниркових тільцях. В результаті цього процесу плазма крові фільтрується в просвіт капсули Шумлянського-Боумена і утворюється первинна сеча – ультрафільтрат плазми крові, який за складом відрізняється від неї тільки відсутністю білків. Фільтрація – пасивний процес, що здійснюється під впливом сили, яка носить назву ефективного фільтраційного тиску (Ре.ф.) і розраховується за формулою: Ре.ф. = Рг.к. – (Ро.к. + Рг.капс.), де: Рг.к. – гідростатичний тиск крові в капілярах ниркового тільця (близько 70 мм.рт.ст.); Ро.к. – онкотичний тиск плазми крові (близько 30 мм.рт.ст.); Рг.капс. – гідростатичний тиск рідини (первинної сечі) в капсулі Шумлянського-Боумена (близько 20 мм.рт.ст.). Таким чином: Ре.ф. = 70 – (30 + 20) = 20 мм.рт.ст. Плазма крові фільтрується в просвіт капсули через нирковий фільтр, який складається з трьох шарів: -шар ендотеліоцитів капілярів (1); -базальна мембрана (2); -шар подоцитів (епітелій капсули - 3); Ендотелій капілярів плоский фенестрований, лежить на базальній мембрані. З другого боку до неї за допомогою цитоподій прикріплюються подоцити. Базальна мембрана ниркового фільтра трьохшарова, найбільш щільним являється її середній шар, який має пори діаметром до 7 мкм. Проникність ниркового фільтру визначається, перш за все, станом базальної мембрани: діаметр, кількість пор, негативний заряд всередині пор в значному ступені визначають рух складових компонентів крові в просвіт капсули. В нормі через нирковий фільтр не проходять форменні елементи крові, білки (можлива фільтрація тільки невеликої кількості низькомолекулярних білків – альбумінів). Тому, ультрафільтрат плазми крові в капсулі Шумлянського-Боумена (первинна сеча) відрізняється за складом від плазми крові відсутністю білків. Проникність ниркового фільтру може змінюватись за фізіологічних умов під впливом речовин, що виробляються самою ниркою. При цьому змінюється так званий коефіцієнт фільтрації (КФ). В патологічних умовах проникність ниркового фільтру (при його ураженні) може значно зростати в сечі з’являються форменні елементи, а саме – еритроцити (гематурія), білок (протеїнурія). Факторами, які впливають на інтенсивність клубочкової фільтрації, являються: -гідростатичний тиск крові в капілярах (Рг.к.); -онкотичний тиск крові (Ро.к.); -гідростатичний тиск первинної сечі в капсулі Шумлянського-Боумена (Рг.капс.); -КФ, величина якого визначається станом ниркового фільтру (його проникністю); -величина ефективного ниркового кровотоку. Регуляція процесу клубочкової фільтрації звідиться до зміни: -вуличини ефективного ниркового кровотоку; -величини тиску крові в капілярах клубочка. Ефективний нирковий кровотік змінюється без зміни тиску крові в капілярах, якщо тонус приносної та виносної артеріол змінюється однаково. Виражене в різній ступені звуження (розширення) приносних та виносних артеріол в нирках веде до зміни тиску в капілярах зміна Ре.ф. зміна процесу клубочкової фільтрації зміна процесів сечоутворення. Таким чином, зміна Ре.ф. змінює рівень клубочкової фільтрації в кожному окремому нефроні. Зміна ефективного ниркового кровотоку змінює кількість функціональних клубочків. Симпатична нервова система (при високій ступені активності) і катехоламіни (при високій концентрації) звужують і приносну, і виносну артеріоли значне зниження ниркового кровотоку зменшення діурезу. Ангіотензин-ІІ звужує виносні артеріоли зниження ниркового кровотоку при підвищенні тиску в капілярах клубочків. Дата добавления: 2015-12-15 | Просмотры: 840 | Нарушение авторских прав |
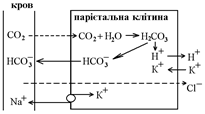 Соляна кислота секретується парієтальними клітинами залоз шлунка, в яких дуже активний фермент карбоангідраза (КА), який каталізує утворення вугільної кислоти H2CO3 із води та вуглекислого газу (H2O + CO2), яка далі розщеплюється на йони водню та гідрокарбонату (H+ та HCO3-).
Соляна кислота секретується парієтальними клітинами залоз шлунка, в яких дуже активний фермент карбоангідраза (КА), який каталізує утворення вугільної кислоти H2CO3 із води та вуглекислого газу (H2O + CO2), яка далі розщеплюється на йони водню та гідрокарбонату (H+ та HCO3-).
 2. Антиперистальтика забезпечує рух кишки в проксимальному на-прямку. В нормі у здорової людини антиперистальтика є тільки в товстому кишківнику. В основі антиперистальтики лежить метасимпатичний рефлекс, який починається з подразнення хімусом механо- та хеморецепторів кишки.
2. Антиперистальтика забезпечує рух кишки в проксимальному на-прямку. В нормі у здорової людини антиперистальтика є тільки в товстому кишківнику. В основі антиперистальтики лежить метасимпатичний рефлекс, який починається з подразнення хімусом механо- та хеморецепторів кишки.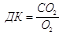 В базолатеральних відділах мембран епітеліоцитів працюють Na-насоси, які активно (з затратою енергії АТФ) транспортують йони Na із епітеліоцитів в інтерстицій (міжклітинну рідину), а потім він пасивно надходить в кров. За рахунок роботи натрієвих насосів в епітеліоцитах створюється низька концентрація Na і внаслідок цього він по градієнту концентрації пасивно входить в епітеліоцити з порожнини кишківника, тобто в кінцевому результаті всмоктування Na проходить за механізмом первинного пасивного активного транспорту. За добу всмоктується 25 – 35г. Na, головним чином в тонкому кишківнику, хоча цей процес проходить також і в товстому кишківнику.
В базолатеральних відділах мембран епітеліоцитів працюють Na-насоси, які активно (з затратою енергії АТФ) транспортують йони Na із епітеліоцитів в інтерстицій (міжклітинну рідину), а потім він пасивно надходить в кров. За рахунок роботи натрієвих насосів в епітеліоцитах створюється низька концентрація Na і внаслідок цього він по градієнту концентрації пасивно входить в епітеліоцити з порожнини кишківника, тобто в кінцевому результаті всмоктування Na проходить за механізмом первинного пасивного активного транспорту. За добу всмоктується 25 – 35г. Na, головним чином в тонкому кишківнику, хоча цей процес проходить також і в товстому кишківнику.