|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Регуляция дыхания. Дыхательные рефлексыДыхательные рефлексы Важное биологическое значение, особенно в связи с ухудшением экологических условий и загрязнением атмосферы, имеют защитные дыхательные рефлексы – чихание и кашель. Чихание – раздражение рецепторов слизистой оболочки полости носа, например пылевыми частицами или газообразными наркотическими веществами, табачным дымом, водой вызывает сужение бронхов, брадикардию, снижение сердечного выброса, сужение просвета сосудов кожи и мышц. Различные химические и механические раздражения слизистой оболочки носа вызывают глубокий сильный выдох – чихание, способствующее стремлению избавиться от раздражителя. Афферентным путем этого рефлекса является тройничный нерв. Кашель – возникает при раздражении механо- и хеморецепторов глотки, гортани, трахеи и бронхов. При этом после вдоха сильно сокращаются мышцы выдоха, резко повышается внутригрудное и внутрилегочное давление, открывается голосовая щель и воздух из дыхательных путей под большим напором высвобождается наружу и и удаляет раздражающий агент. Кашлевый рефлекс является основным легочным рефлексом блуждающего нерва.
Дыхательный центр продолговатого мозга Дыхательный центр, совокупность нескольких групп нервных клеток (нейронов), расположенных в разных отделах центральной нервной системы, преимущественно в ретикулярной формации продолговатого мозга. Постоянная координированная ритмическая активность этих нейронов обеспечивает возникновение дыхательных движений и их регуляцию в соответствии с возникающими в организме изменениями. Импульсы от Д. ц. поступают в двигательные нейроны передних рогов шейного и грудного отделов спинного мозга, от которых возбуждение передаётся к дыхательной мускулатуре. Активность Д. ц. регулируется гуморально, т. е. составом омывающей его крови и тканевой жидкости, и рефлекторно, в ответ на импульсы, поступающие от рецепторов в дыхательной, сердечно-сосудистой, двигательной и др. системах, а также от высших отделов центральной нервной системы. Состоит из центра вдоха и центра выдоха. Дыхательный центр состоит из нервных клеток (дыхательных нейронов), для которых характерна периодическая электрическая активность в одну из фаз дыхания. Нейроны дыхательного центра локализованы двусторонне в продолговатом мозге в виде двух вытянутых столбов вблизи obex — точки, где центральный канал спинного мозга впадает в четвертый желудочек. Эти два образования дыхательных нейронов в соответствии с их положением относительно дорсальной и вентральной поверхности продолговатого мозга обозначают как дорсальная и вентральная дыхательные группы Дорсальная дыхательная группа нейронов образует вентролатеральную часть ядра одиночного тракта. Дыхательные нейроны вентральной дыхательной группы расположены в области n. ambiguus каудальнее уровня obex, n. retroambigualis непосредственно ростральнее obex и представлены комплексом Бетзингера, который находится непосредственно вблизи n. retrofacialis вентролатеральных отделов продолговатого мозга. В состав дыхательного центра входят нейроны двигательных ядер черепно-мозговых нервов (обоюдное ядро, ядро подъязычного нерва), которые иннервируют мышцы гортани и глотки.
Взаимодействие нейронов инспираторных и экспираторных зон Дыхательные нейроны, активность которых вызывает инспирацию или экспирацию, называются соответственно инспираторными или экспираторными. Между группами нейронов, управляющими вдохом и выдохом, существуют реципрокные отношения. Возбуждение экспираторного центра сопровождается торможением в инспираторном центре и наоборот. Инспираторные и экспираторные нейроны в свою очередь делятся на «ранние» и «поздние». Каждый дыхательный цикл начинается с активизации «ранних» инспираторных нейронов, затем возбуждаются «поздние» инспираторные нейроны. Также последовательно возбуждаются экспираторные нейроны, которые тормозят инспираторные нейроны и прекращают вдох. Современные исследователи показали, что нет четкого разделения на инспираторный и экспираторный отделы, а есть скопления дыхательных нейронов с определенной функцией
Представление об ауторитме дыхания. Влияние ph крови на процесс дыхания. Если происходит снижение pH артериальной крови по сравнению с нормальным уровнем, равным 7,4, вентиляция легких увеличивается. При возрастании pH выше нормы вентиляция уменьшается, хотя и в несколько меньшей степени. Ауторитмия – это волны возбуждения и соответствующие им «шевеления» животного, происходящие с определённой периодичностью. ауторитмия - самопроизвольная активность центральной нервной системы, которая осуществляется без какого бы то ни было воздействия афферентной стимуляции и проявляются в ритмизированных и скоординированных движениях организма. Пневмотоксический центр варолиева мота. Взаимодействие с дыхательным центром продолговатого мозга В варолиевом мосту находятся ядра дыхательных нейронов образующих пневмотаксический центр. Считается, что дыхательные нейроны моста участвуют в механизме смены вдоха и выдоха и регулируют величину дыхательного объема. Дыхательные нейроны продолговатого мозга и варолиева моста связаны между собой восходящими и нисходящими нервными путями и функционируют согласованно. Получив импульсы от инспираторного центра продолговатого мозга, пневмотаксический центр посылает их и экспираторному центру продолговатого мозга, возбуждая последний. Инспираторные нейроны тормозятся. Разрушения мозга между продолговатым мозгом и мостом удлиняет фазу вдоха. Спинной мозг; мотонейроны ядер межреберных нервов и ядра диафрагмального нерва, взаимодействие с дыхательным центром продолговатого мозга. В передних рогах спинного мозга на уровне Мотонейроны, иннервирующие межреберные мышцы, находятся в передних рогах на уровнях Роль коры больших полушарий в регуляции дыхания. Определенные зоны коры больших полушарий осуществляют произвольную регуляцию дыхания в соответствии с особенностями влияния на организм факторов внешней среды и связанными с этим гомеостатическими сдвигами. Помимо дыхательного центра, расположенного в стволе мозга, на состояние функции дыхания влияют и корковые зоны, обеспечивающие его произвольную регуляцию. Расположены они в коре соматомоторных отделов и медиобазальных структур головного мозга. Есть мнение, что моторные и премоторные области коры по воле человека облегчают, активируют дыхание, а кора медиобазальных отделов больших полушарий тормозит, сдерживает дыхательные движения, влияя и на состояние эмоциональной сферы, а также степень сбалансированности вегетативных функций. Эти отделы коры больших полушарий влияют и на адаптацию функции дыхания к сложным движениям, связанным с поведенческими реакциями, и приспосабливают дыхание к текущим ожидаемым метаболическим сдвигам.
Регуляция кровяного давления, кровотока В вентролатеральных отделах продолговатого мозга сосредоточены образования, соответствующие по своим характеристикам тем представлениям, которые вкладывают в понятие «вазомоторный центр». Здесь сконцентрированы нервные элементы, играющие ключевую роль в тонической и рефлекторной регуляции кровообращения. В вентральных отделах продолговатого мозга расположены нейроны, изменение тонической активности которых ведет к активации симпатических преганглионарных нейронов. Структуры этих отделов мозга контролируют выброс вазопрессина клетками супраоптического и паравентрикулярного ядер гипоталамуса.
Доказаны проекции нейронов каудальной части вентральных отделов продолговатого мозга к клеткам его ростральной части, что свидетельствует о возможности тонического угнетения активности этих клеток. Функционально значимы связи структур вентральных отделов продолговатого мозга с ядром солитарного тракта, которое играет ключевую роль в обработке афферентации от хемо- и барорецепторов сосудов.
В продолговатом мозге расположены нервные центры, тормозящие деятельность сердца (ядра блуждающего нерва). В ретикулярной формации продолговатого мозга находится сосудодвигательный центр, состоящий из двух зон: прессорной и депрессорной. Возбуждение прессорной зоны приводит к сужению сосудов, а возбуждение депрессорной зоны - к их расширению. Сосудодвигательный центр и ядра блуждающего нерва постоянно посылают импульсы, благодаря которым поддерживается постоянный тонус: артерии и артериолы постоянно несколько сужены, а сердечная деятельность замедлена. В. Ф. Овсянниковым (1871) было установлено, что нервный центр, обеспечивающий определенную степень сужения артериального русла — сосудодвигательный центр — находится в продолговатом мозге. Локализация этого центра определена путем перерезки ствола мозга на разных уровнях. Если перерезка произведена у собаки или кошки выше четверохолмия, то АД не изменяется. Если перерезать мозг между продолговатым и спинным мозгом, то максимальное давление крови в сонной артерии понижается до 60—70 мм рт.ст. Отсюда следует, что сосудодвигательный центр локализован в продолговатом мозге и находится в состоянии тонической активности, т. е. длительного постоянного возбуждения. Устранение его влияния вызывает расширение сосудов и падение АД. Более детальный анализ показал, что сосудодвигательный центр продолговатого мозга расположен на дне IV желудочка и состоит из двух отделов — прессорного и депрессорного. Раздражение прессорного отдела сосудодвигательного центра вызывает сужение артерий и подъем, а раздражение второго — расширение артерий и падение АД. Считают, что депрессорный отдел сосудодвигательного центра вызывает расширение сосудов, понижая тонус прессорного отдела и снижая, таким образом, эффект сосудосуживающих нервов. Влияния, идущие от сосудосуживающего центра продолговатого мозга, приходят к нервным центрам симпатической части вегетативной нервной системы, расположенным в боковых рогах грудных сегментов спинного мозга, регулирующих тонус сосудов отдельных участков тела. Спинномозговые центры способны через некоторое время после выключения сосудосуживающего центра продолговатого мозга немного повысить давление крови, снизившееся вследствие расширения артерий и артериол. Кроме сосудодвигательных центров продолговатого и спинного мозга, на состояние сосудов оказывают влияние нервные центры промежуточного мозга и больших полушарий.
Гипоталамическая регуляция висцеральных функций Если стимулировать электрическим током различные зоны гипоталамуса, то можно вызвать и сужение и расширение сосудов. Импульс передается по волокнам заднего продольного пучка. Часть волокон проходят через области, не переключаются и идут к вазомоторным нейронам. Информация поступает от осморецепторов, они улавливают состояние воды внутри и внеклетки, содержащейся в гипоталамусе. Активация осморецепторов вызывает гормональный эффект – выброс вазопрессина, а это вещество обладает сильным сосудосуживающим действием, он обладает удерживающим свойством. Особое значение НЭС (нейроэндокринной регуляции) имеет в регуляции висцеральных (“отно-сящихся к внутренним органам”) функций организма. Установлено, что эфферентные влияния ЦНС на висцеральные функции реализуются в норме и при патологии как вегетативными, так и эндокринными аппаратами (Speckmann, 1985). В отличие же от коры гипоталамус, очевидно, постоянно участвует в управлении работой висцеральных систем организма. Обеспечивает постоянство внутренней среды. Контроль за действием симпатической и парасимпатической систем, иннервирующих внутренние органы, сосуды, гладкую мускулатуру, железы внутренней и внешней секреции, осуществляет “висцеральный мозг”, который представлен центральными вегетативными аппаратами (вегетативные ядра) гипоталамической области (О.Г.Газенко и соавт., 1987). В свою очередь, гипоталамус находится под контролем определенных областей коры (в частности, лимбической) больших полушарий. Координация деятельности всех трех частей автономной нервной системы осуществляется сегментарными и надсегментарными центрами (аппаратами) при участии коры большого мозга. В сложноорганизованном отделе промежуточного мозга -- гипоталамической области, находятся ядра, имеющие непосредственное отношение к регуляции висцеральных функций. Хемо и барорецепторы кровеносных сосудов Афферентные импульсы от барорецепторов поступают к сосудодвигательному центру продолговатого мозга. Эти импульсы оказывают тормозное влияние на симпатические центры и возбуждающее на парасимпатические. В результате снижается тонус симпатических сосудосуживающих волокон (или так называемый вазомоторный тонус), а также частота и сила сердечных сокращений. Поскольку импульсация от барорецепторов наблюдается в широком диапазоне значений артериального давления, их тормозные влияния проявляются даже при «нормальном» давлении. Иными словами барорецепторы оказывают постоянное депрессорное действие. При повышении давления импульсация от барорецепторов возрастает, и сосудодвигательный центр затормаживается сильнее; это приводит к еще большему расширению сосудов, причем сосуды в разных областях расширяются в разной степени. При падении давления импульсация от барорецепторов уменьшается и развиваются обратные процессы, приводящие, в конечном счете, к повышению давления. Возбуждение хеморецепторов приводит к снижению частоты сокращений сердца и сужению сосудов в результате прямого действия на циркуляторные центры продолговатого мозга. При этом эффекты, связанные с сужением сосудов, преобладают над последствиями снижения сердечного выброса, и вследствие этого артериальное давление повышается. барорецепторы расположены в стенках артерий. Увеличение артериального давления приводит к растяжению барорецепторов, сигналы от которых поступают в центральную нервную систему. Затем сигналы обратной связи направляются к центрам автономной нервной системы, а от них — к сосудам. В результате давление понижается до нормального уровня. Барорецепторы чрезвычайно быстро реагируют на изменения артериального давления. Хеморецепторы чувствительны к химическим компонентам крови. артериальные хеморецепторы реагируют на изменения концентрации в крови кислорода, углекислоты, водородных ионов, питательных веществ и гормонов, уровня осмотического давления; благодаря хеморецепторам поддерживается гомеостаз. Дата добавления: 2015-12-15 | Просмотры: 1318 | Нарушение авторских прав |
 -
-  располагаются мотонейроны, образующие диафрагмальный нерв. Диафрагмальный нерв - смешанный нерв, который осуществляет чувствительную иннервацию плевры и перикарда, - входит в состав шейного сплетения; образуется передними ветвями нервов СЗ-С5. Отходит с обеих сторон шеи от шейного сплетения третьего, четвертого (и иногда пятого) шейных спинномозговых нервов и направляется вниз к диафрагме, проходя между легкими и сердцем (между средостенной плеврой и перикардом). Проходящие по этим нервам от головного мозга импульсы вызывают периодические сокращения диафрагмы во время дыхания.
располагаются мотонейроны, образующие диафрагмальный нерв. Диафрагмальный нерв - смешанный нерв, который осуществляет чувствительную иннервацию плевры и перикарда, - входит в состав шейного сплетения; образуется передними ветвями нервов СЗ-С5. Отходит с обеих сторон шеи от шейного сплетения третьего, четвертого (и иногда пятого) шейных спинномозговых нервов и направляется вниз к диафрагме, проходя между легкими и сердцем (между средостенной плеврой и перикардом). Проходящие по этим нервам от головного мозга импульсы вызывают периодические сокращения диафрагмы во время дыхания.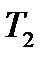 -
-  (
( -
- 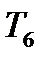 - мотонейроны инспираторных мышц,
- мотонейроны инспираторных мышц,  -
-  - экспираторных). Двигательные ветви межреберных нервов иннервируют аутохтонные мышцы (вдоха) груди и мышцы живота. Установлено, что одни регулируют преимущественно дыхательную, а другие познотоническую активность межреберных мышц.
- экспираторных). Двигательные ветви межреберных нервов иннервируют аутохтонные мышцы (вдоха) груди и мышцы живота. Установлено, что одни регулируют преимущественно дыхательную, а другие познотоническую активность межреберных мышц.