|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Функции эритроцитовОни весьма многообразны: перенос кислорода от легких к тканям; перенос углекислого газа от тканей к легким; транспортировка питательных веществ - адсорбированных на их поверхности аминокислот - от органов пищеварения к клеткам организма; поддержание рН крови на относительно постоянном уровне благодаря наличию гемоглобина; активное участие в процессах иммунитета: эритроциты адсорбируют на своей поверхности различные яды, которые затем разрушаются клетками мононуклеарной фагоцитарной системы (МФС); осуществление процесса свертывания крови. В них найдены почти все факторы, которые содержатся в тромбоцитах. Кроме того, их форма удобна для прикрепления нитей фибрина, а их поверхность катализирует гемостаз. Г е м о л и з. Разрушение оболочки эритроцитов и выход из них гемоглобина называется гемолизом. Он может быть химический, когда их оболочка разрушается химическими веществами (кислотами, щелочами, сапонином, мылом, эфиром, хлороформом и т. д.); физический, который подразделяют на механический (при сильном встряхивании), температурный (под действием высокой и низкой температуры), лучевой (под действием рентгеновских или ультрафиолетовых лучей). Осмотический гемолиз - разрушение эритроцитов в воде или гипотонических растворах, осмотическое давление которых меньше, чем в плазме крови. Вследствие того, что давление внутри эритроцитов больше, чем в окружающей среде, вода переходит в эритроциты, их объем увеличивается и оболочки лопаются, а гемоглобин выходит наружу. Если окружающий раствор имеет достаточно низкую концентрацию соли, наступает полный гемолиз и вместо нормальной непрозрачной крови образуется относительно прозрачная «лаковая» кровь. Если раствор, в котором находятся эритроциты, менее гипотоничен, наступает частичный гемолиз. Биологический гемолиз может возникнуть при переливании крови, если кровь несовместима, при укусах некоторых змей и т.д. В организме постоянно в небольших количеств происходит гемолиз при отмирании старых эритроцитов. При этом эритроциты разрушаются в печени, селезенке, красном костном мозге, освободившийся гемоглобин поглощается клетками этих органов, а в плазме циркулирующей крови он отсутствует.
Г е м о г л о б и н. Свою основную функцию - перенос газов кровью - эритроциты выполняют благодаря наличию в них гемоглобина, который представляет сложный белок - хромопротеид, состоящий из белковой части (глобина) и небелковой пигментной группы (гема), соединенных между собой гистидиновым мостиком. В молекуле гемоглобина четыре гема. Гем построен из четырех пирроловых колец и содержит двухвалентное железо. Он является активной, или так называемой простетической, группой гемоглобина и обладает способностью присоединять и отдавать молекулы кислорода. У всех видов животных гем имеет одинаковое строение, в то время как глобин отличается по аминокислотному составу. Гемоглобин, присоединивший кислород, превращается в оксигемоглобин (НЬО) ярко-алого цвета, что и определяет цвет артериальной крови. Оксигемоглобин образуется в капиллярах легких, где напряжение кислорода высокое. В капиллярах тканей, где кислорода мало, он распадается на гемоглобин и кислород. Гемоглобин, отдавший кислород, называют восстановленным или редуцированным гемоглобином (НЬ). Он придает венозной крови вишневый цвет. И в оксигемоглобине, и в восстановленном гемоглобине атомы железа находятся в двухвалентном состоянии. Третье физиологическое соединение гемоглобина - карбогемоглобин - соединение гемоглобина с углекислым газом. Таким образом, гемоглобин участвует в переносе углекислого газа из тканей в легкие. Карбогемоглобин содержится в венозной крови. При действии на гемоглобин сильных окислителей (бертолетовая соль, перманганат калия, нитробензол, анилин, фенацетин и т. д.) железо окисляется и переходит в трехвалентное. При этом гемоглобин пре вращается в метгемоглобин и приобретает коричневую окраску. Являясь продуктом истинного окисления гемоглобина, последний прочно удерживает кислород и поэтому не может служить в качестве его переносчика. Образование значительного количества метгемоглобина резко ухудшает дыхательные функции крови. Это может случиться после введения в организм лекарств, обладающих окислительными свойствами. Метгемоглобин - патологическое соединение гемоглобина. Гемоглобин очень легко соединяется с угарным газом, при этом образуется карбоксигемоглобин (НЬСО) Химическое сродство окиси углерода к гемоглобину примерно в 200 раз больше, чем кислорода. Поэтому достаточно примеси небольшого количества СО к воздуху, чтобы образовалось значительное число молекул этого соединения. Оно весьма прочное, и гемоглобин, блокированный СО, не может быть переносчиком кислорода. Поэтому угарный газ очень ядовит. При вдыхании воздуха, содержащего 0,1 % СО, через 30-60 мин развиваются тяжелые последствия кислородного голодания (рвота, потеря сознания). При содержании в воздухе 1 % СО через несколько минут наступает смерть. Пострадавших людей и животных необходимо вывести на чистый воздух или дать вдохнуть кислород. Под влиянием высокого давления кисло рода происходит медленное расщепление карбоксигемоглобина. При действии соляной кислоты на гемоглобин образуется гемин. В этом соединении железо находится в окисленной трехвалентной форме. Для его получения каплю высушен ной крови нагревают на предметном стекле с кристалликами поваренной соли и 1-2 каплями ледяной уксусной кислоты. Коричневые ромбические кристаллы гемина рассматривают в микроскоп. Кристаллы гемина разных видов животных отличаются по своей форме. Это обусловлено видовыми различиями в структуре глобина. Данную реакцию, получившую название геминовой пробы, можно использовать для обнаружения следов крови. При рассматривании в спектроскоп разведенного раствора оксигемоглобина видны две характерные темные полосы поглощения в желто-зеленой части спектра, между фраунгоферовыми линиями Д и Е. Для восстановленного гемоглобина характерна одна широкая полоса поглощения в желто-зеленой части спектра. Спектр карбоксигемоглобина очень похож на спектр оксигемоглобина. Их можно различить добавлением восстанавливающего вещества. Карбоксигемоглобин и после этого имеет две полосы поглощения. Метгемоглобин имеет характерный спектр: одна узкая полоса поглощения находится слева, на границе красной и желтой частей спектра, другая узкая полоса на границе желтой и зеленой зон и широкая темная полоса в зеленой части.
Количество гемоглобина определяют колориметрическим методом и выражают в грамм-процентах (г%), а затем с помощью коэффициента пересчета по Международной системе единиц (СИ), который равен 10, находят количество гемоглобина в граммах на литр (г/л). Оно зависит от вида животных. На содержание эритроцитов и гемоглобина влияют возраст, пол, порода, высота над уровнем моря, работа, кормление. Так, новорожденные животные имеют более высокое содержание эритроцитов и гемоглобина, чем взрослые; у самцов количество эритроцитов на 5-10 % выше, чем у самок. Количество эритроцитов у скаковых лошадей больше, чем у тяжеловозов, и доходит до 10-10,5 млн/мкл крови, или по системе СИ 10- 10,5.1012 л, а у тяжеловозов составляет 7,4-7,б млн/мкл Уменьшение давления кислорода на большой высоте над уровнем моря стимулирует образование эритроцитов. Поэтому у овец, коров на горных пастбищах количество эритроцитов и гемоглобина повышено. Интенсивная физическая на грузка вызывает такое же действие. Количество гемоглобина в крови рысаков, равное до бега в среднем 12,6 г% (126 г/л), после бега увеличивается до 16-18 г% (160- 180 г/л). Ухудшение кормления ведет к уменьшению содержания эритроцитов и гемоглобина. Особенно большое влияние оказывает недостаток микроэлементов и витаминов (цианкобаламина, фолиевой кислоты и др.).
Для определения насыщенности каждого эритроцита гемоглобином служит цветной показатель или индекс I
В норме цветной показатель равен 1. Если он меньше 1, содержание гемоглобина в эритроцитах понижено (гипохромия), если больше 1 - повышено (гиперхромия). Миоглобин. В скелетных и сердечных мышцах находится мышечный гемоглобин (миоглобин). Он имеет сходство и различие с гемоглобином крови. Сходство этих двух веществ выражается наличии одной и той же простетической группы, одинакового количества железа и в способности давать обратимые соединения с О и СО. Однако масса миоглобина гораздо меньше, и он обладает значительно большим сродством к кислороду, чем гемоглобин крови, а поэтому приспособлен к функции депонирования (связывания) кислорода, что имеет большое значение для снабжения кислородом сокращающихся мышц. Когда мышцы сокращаются, их кровоснабжение временно ухудшается из- за сжатия капилляров. И в этот момент миоглобин служит важным источником кислорода. Он «запасает» кислород во время расслабления и отдает его во время сокращения. Содержание миоглобина увеличивается под влиянием мышечных грузок. Скорость оседания эритроцитов (СОЭ). Для определения СОЭ кровь смешивают раствором лимоннокислого натрия, собирают в стеклянную трубочку пробирку с миллиметровыми делениями. Через некоторое время отсчитывают высоту верхнего прозрачного слоя. СОЭ различна у животных разного вида. Очень быстро оседают эритроциты лошади, весьма медленнно - жвачных. На величину СОЭ влияет физиологическое состояние организма. Усиленная тренировка замедляет эту реакцию. У спортивных лошадей, отобранных для олимпийских соревнований, при средней нагрузке СОЭ за первые 15 мин равнялась 9,6 мм. Через 2 мес напряженных тренировок за те же первые 15 мин она равнялась 2,6 мм. СОЭ сильно увеличивается во время беременности, а также при хронических воспалительных процессах, инфекционных заболеваниях, злокачественных опухолях. Это свзывают с увеличением в плазме количества крупномолекулярных белков - глобулинов и особенно фибриногена. Вероятно, крупномолекулярные белки уменьшают электрический заряд и электроотталкивание эритроцитов, что способствует большей скорости их оседания.
Продолжительность жизни эритроцитов. У разных животных она неодинакова. Эритроциты у лошади находятся в сосудистом русле в среднем 100 дн., у крупного рогатого скота - 120-16, у овцы – 130, у северного оленя - у кролика - 45-60 дн.
В 1951 г. А. Л. Чижевский в результате экспериментальных исследований и математических расчётов пришел к выводу, что в артериальных сосудах у здоровых людей и животных эритроциты движутся в системе, состоящих из монетных столбиков. Причем монетные столбики из эритроцитов крупного диаметра примыкают к медленному пристеночному слою крови, а монетные столбики из эритроцитов малого диаметра несутся в быстром осевом потоке крови. Кроме поступательного движения, эритроциты совершают и вращательные движения вокруг собственной оси. При заболеваниях происходит нарушение пространственного расположения эритроцитов в сосудах. Лейкоциты. Белые кровяные летки имеют цитоплазму и ядро. Их подразделяют на две большие группы: зернистые (гранулоциты) и незернистые (агранулоциты). В цитоплазме зернистых лейкоцитов содержатся зернышки (гранулы), в цитоплазме незернистых гранулы отсутствуют.
Зернистые лейкоциты зависимости от окраски гранул различают эозинофильные (гранулы окрашиваются в розовый цвет кислыми красками, например эозином), базофильные (в синий цвет основными красками) и нейтрофильные теми и другими красками в розово-фиолетовый цвет), У юных гранулоцитов ядро округлое, у молодых оно в виде подковы или палочки (палочкоядерные); по мере развития ядро перешнуровывается и разделяется на несколько сегментов. Сегментоядерные, нейтрофилы составляют основную массу гранулоцитов. У птиц вместо сегментоядерных нейтрофилов присутствуют псевдоэозинофилы, в цитоплазме которых содержатся палочкообразные и веретеновидные гранулы. Незернистые лейкоциты делятся на лимфоциты и моноциты. Лимфоциты имеют крупное ядро, окруженное узким поясом цитоплазмы. В зависимости от размера различают большие, средние и малые лимфоциты. Лимфоциты составляют большую часть белых кровяных клеток: у крупного рогатого скота 50--60 % всех лейкоцитов, У свиней - 45--б0, у овец - 55--65, у коз - 40--50, у кроликов – 50--65, у кур - 45-65 %. Этим видам животных присущ так называемый лимфоцитарный профиль крови. У лошадей и плотоядных преобладают сегментоядерные нейтрофилы - нейтрофильный профиль крови. Однако и у этих животных количество лимфоцитов значительное -- 20-40 % всех лейкоцитов, Моноциты - самые большие клетки крови, в основном округлой формы, с хорошо выраженной цитоплазмой. В крови птиц, кроме того, имеются клетки Тюрка - крупные, с эксцентрично расположенным ядром и значительным количеством цитоплазмы. Процентное соотношение различных форм лейкоцитов называют лейкоцитарной формулой (лейкоформулой) или лейкограммой. Она имеет видовые отличия и характерно изменяется при инфекционных и паразитарных заболеваниях, поэтому ее изучению придается большое значение в клинике. Общее количество лейкоцитов в крови значительно меньше, чем эритроцитов. У млекопитающих оно составляет около 0,1--0,2 % от числа эритроцитов, у птиц - несколько больше (около 0,5--1 %). Увеличение количества лейкоцитов называют лейкоцитозом, а уменьшение - лейкопенией. Различают два вида лейкоцитозов: физиологический и реактивный. Физиологический, в свою очередь, делят на: 1) пищеварительный (значительное увеличение количества лейкоцитов происходит после приема корма; особенно выражен у лошадей, свиней, собак и кроликов); 2) миогенный (развивается после тяжелой мышечной работы); 3) эмоциональный; 4) при болевых воздействиях; 5) при беременности. Физиологвческие лейкоцитозы по своей природе являются перераспределительными, то есть лейкоциты в этих случаях выходят из депо (селезенка, костный мозг, лимфатические узлы). Они характеризуются быстрым развитием, кратковременностью, отсутствием изменений лейкоцитарной формулы.
Реактивные, или истинные, лейкоцитозы бывают при воспалительных процессах, инфекционных заболеваниях. При этом резко усиливается образование белых кровяных телец в органах кроветворения и количество лейкоцитов в крови увеличивается значительнее, чем при перераспределительном лейкоцитозе. Но главное отличие заключается в том, что при реактивных лейкоцитозах меняется лейкоцитарная формула: в крови увеличивается количество молодых форм нейтрофилов - миелоцитов, юных, палочкоядерных. По ядерному сдвигу влево оценивают тяжесть заболевания и реактивность организма.
В последнее время лейкопении встречаются чаще, чем раньше. Это объясняется повышением фоновой радиоактивности и другими причинами, связанными с техническим прогрессом. Особенно тяжелые лейкопении, вызванные поражением костного мозга, наблюдают при лучевой болезни. Лейкопению выявляют и при некоторых инфекционных заболеваниях (паратиф телят, чума свиней). Функции лейкоцитов. Лейкоциты играют важную роль в защитных и восстановительных процессах организма. Моноциты и нейтрофилы способны к амебеоидному движению. Скорость движения последних может доходить до 40 мкм/м что равно расстоянию, в З-4 раза превышающему диаметр этих клеток. Данные виды лейкоцитов проходят через эндотелий капилляров и движутся в тканях к месту скопления микробов, инородных частиц или разрушающихся клеток самого организма. Один нейтрофил может захватить до 20-30 бактерий, а моноцит фагоцитирует до 100 микробов. Кроме протеолитических ферментов, эти формы лейкоцитов выделяют, а так же адсорбируют на своей поверхности и переносят вещества, обезвреживаюшие микробы и чужеродные белки - антитела. Эозинофилы участвуют в разрушении и обезвреживании чужеродных белков и токсинов белкового происхождения. Предполагают, что эозинофилы адсорбируют и расщепляют гистамин благодаря образованию фермента гистаминазы. Гистамин является продуктом промежуточного обмена белков и обладает сильным биологическим действием. Количество эозинофилов в крови повышается при паразитарных заболеваниях, аллергических состояниях и болезнях кожи. Базофилы имеют слабовыраженную способность к фагоцитозу или совсем ее не обнаруживают. Как и тучные клетки соединительной ткани, они синтезируют гепарин - вещество, препятствующее свертыванию крови. Кроме того, базофилы способны образовывать гистамин. Гепарин предотвращает свертывание крови, а гистамин расширяет капилляры в очаге воспаления, что ускоряет процесс рассасывания и заживления. Лимфоциты принимают участие в выработке антител, поэтому имеют большое значение в создании невосприимчивости к инфекционным заболеваниям (инфекционный иммунитет), а также ответственны за реакции на введение чужеродных белков и отторжение чужеродных тканей при пересадке органов (трансплантационный иммунитет). Ведущую роль в иммунитете, особенно трансплантационном, играют так называемые Т-лимфоциты. Они образуются из клеток-предшественников в костном мозге, проходят дифференцировку в тимусе (зобной железе), а затем переходят в лимфатические узлы, селезенку или циркулирующую кровь, где на их долю приходится 40-70 % всех лимфоцитов. Т-лимфоциты неоднородны. Среди них выделяют несколько групп: 1) хелперы (помощники) взаимодействуют с В-лимфоцитами и превращают их в плазматические клетки, синтезирующие антитела; 2) супрессоры - подавляют чрезмерные реакции В-лимфоцитов и поддерживают постоянное соотношение различных форм лимфоцитов; З) киллеры (убийцы) - взаимодействуют с чужеродными клетками и разрушают их; 4) амплифайеры - активируют киллеры; 5) клетки иммунной памяти В-лимфоциты образуются в костном мозге, дифференцируются у млекопитающих в лимфоидной ткани кишечника, червеобразного отростка, глоточных и небных миндалин. У птиц дифференцировка проходит в фабрициевой сумке. Сумка по латыни звучит как бурса, отсюда и В-лимфоциты. На их долю приходится 20-З0 % циркулирующих лимфоцитов. Основная функция В-лимфоцитов - выработка антител и создание гуморального иммунитета. После встречи с антигеном В-лимфоциты переселяются в костный мозг, селезенку, лимфатические узлы, где они размножаются и превращаются в плазматические клетки, образующие антитела, иммунные глобулины. В-лимфоциты специфичны: каждая группа их реагирует лишь с одним антигеном и отвечает за выработку антител только против него. Выделяют еще и так называемые нулевые лимфоциты, которые не проходят дифференцировку в органах иммунной системы, но при необходимости могут превращаться в Т- и В лимфоциты. Они составляют 10- 20 % лимфоцитов. Продолжительность жизни лейкоцитов. Большинство из них живет относительно недолго. При помощи методики меченых атомов установлено, что гранулоциты живут максимум 8-10 дн., чаще значительно меньше - часы и даже минуты. Средняя продолжительность жизни нейтрофилов у теленка составляет 5 ч. Среди лимфоцитов различают короткоживушие и долгоживущие формы. Первые (В-лимфоциты) живут от нескольких часов до недели, вторые (Т-лимфоциты) могут жить месяцы и даже годы. Кровяные пластинки (тромбоциты). У млекопитающих эти форменные элементы крови не имеют ядер, у птиц и всех низших позвоночных ядра есть. Кровяные пластинки обладают удивительным свойством менять форму и размеры в зависимости от местоположения. Так, в потоке крови они имеют форму шарика диаметром полмикрона (на границе разрешения оптического микроскопа). Но попав на стенку кровеносного сосуда или на предметное стекло, они распластываются, из круглых становятся звездчатыми, увеличивая площадь в 5-10 раз, диаметр их становится от 2 до 5 мкм. Количество кровяных пластинок зависит от вида животных. Оно возрастает при тяжелой мышечной работе, пищеварении, в период беременности. Отмечены также суточные колебания: днем их больше, чем ночью. Количество кровяных пластинок уменьшается при острых инфекционных заболеваниях, при анафилактическом шоке. В 1882 г. русский ученый В. П. Образцов впервые доказал, что тромбоциты - это самостоятельные элементы крови, происходящие из клеток красного костного мозга - мегакариоцитов (диаметр до 140 мкм). Мегакариоцит - клетка с огромным ядром. Долгое время была принята «теория взрыва», согласно которой «зрелый» мегакариоцит как бы взрывается, распадаясь на мелкие частицы - тромбоциты. Причем ядро мегакариоцита тоже распадается, передавая определенный запас вещества наследственности - ДНК - тромбоцитам. Однако тщательные исследования под электронным микроскопом не подтвердили эту гипотезу. Оказалось, что в цитоплазме мегакариоцита под управлением его гигантского ядра происходит зачатие и развитие 3-4 тыс. тромбоцитов. Затем мегакариоцит выпускает свои цитоплазматические отростки через стенки кровеносных сосудов. В отростках лежат созревшие кровяные пластинки, они отрываются, поступают в кровоток и начинают выполнять свои функции. Но мегакариоцит не прекращает своего существования. Его ядро наращивает новую цитоплазму, в которой проходит новый цикл за рождения, созревания и «рождения» пластинок. Таким образом, «теорию взрыва» сменила «теория рождения». Каждый мегакариоцит за время своего существования в костном мозге дает 8-10 поколений тромбоцитов. Пластинки выбрасываются в кровь из костного мозга в зрелом состоянии с полным набором органелл, но без ядра и ядерного наследственного материала (ДНК). Они существуют, ню не развиваются, тратят себя, но не восстанавливаются. В отсутствие ядра в токе крови возможен только синтез за счет запасов веществ и энергии, полученных от мегакариоцита. Вот почему в кровяном русле каждый тромбоцит живет недолго (3-5 сут). В световом микроскопе к пластинки выглядят как кусочки цитоплазмы с небольшим количеством зернышек внутри. С помощью электронного микроскоп было показано, что за мнимой простотой скрыта своеобразная и сложная организация. Очень сложным оказался и химический состав кровяных пластинок. Они содержат ферменты адреналин, норадреналин, лизоцим, АТФ, гранулы серотонина и целый ряд других веществ. Функции тромбоцитов. Тромбоциты выполняют различные функции. Прежде всего они участвуют в процессе свёртывания крови. Имея очень клейкую поверхность, они способны быстро прилипать к поверхности инородного предмета при соприкосновении с инородными телами или шероховатой поверхностью тромбоциты слипаются, а за тем распадаются на мелкие обломки, и при этом выделяются лежащие в митохондриях вещества - так называемые пластинчатые, или тромбоцитарные, факторы, которые принято обозначать арабскими цифрами. Они принимают участие во всех фазах свертывания крови. Тромбоциты служат строительным материалом для первичного тромба. При свертывании крови кровяные пластинки выпускают мельчайшие отростки - усики звездообразной формы, затем сцепляются ими, образуя каркас, на котором формируется сгусток крови - тромб. Тромбоциты выделяют также вещества, необходимые для уплотнения кровяного сгустка, - ретрактозимы. Важнейший из них - тромбостенин, который по своим свойствам напоминает актомиозин скелетных мышц. Из кровяных пластинок в раненую ткань выделяется тромбоцитарный фактор роста (ТФР), который стимулирует деление клеток, поэтому рана затягивается быстро. Тромбоциты укрепляют стенки кровеносных сосудов. Внутренняя стенка сосуда образована эпителиальными клетками, но прочность ее определяется сцеплением пристеночных тромбоцитов. А они всегда располагаются вдоль стенок кровеносных сосудов, служа своеобразным барьером. Когда прочность стенки сосуда повышена, то подавляющее большинство пристеночных тромбоцитов имеет дендрическую, самую «цепкую» форму, а многие из них находятся на разной стадии внедрения в эпителиальные клетки. Без взаимодействия с тромбоцитами эндотелий сосудов начинает пропускать через себя эритроциты. Тромбоциты переносят различные вещества. Например, серотонин, который адсорбируется пластинками из крови. Это вещество суживает кровеносные сосуды и уменьшает кровотечение. Тромбоциты переносят и так называемые креаторные вещества, необходимые для сохранения структуры сосудистой стенки. На эти цели используется около 15 % циркулирующих в крови тромбоцитов. Тромбоциты обладают способностью к фагоцитозу. Они поглощают и переваривают чужеродные частицы, в том числе и вирусы.
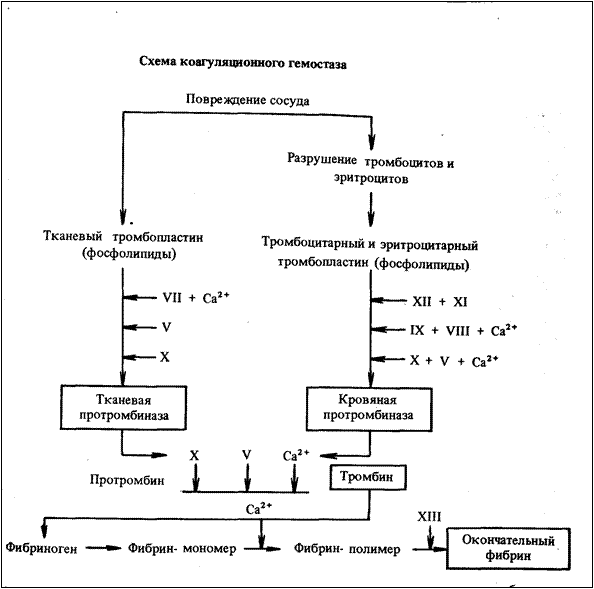
СВЕРТЫВАНИЕ КРОВИ При ранении кровеносного сосуда кровь свертывается, образуется тромб, который закупоривает дефект и препятствует дальнейшему кровотечению. Свертывание крови, или гемокоагуляция, предохраняет организм от кровопотери и является важнейшей защитной реакцией организма. При пониженной свертываемости крови даже ничтожное ранение может привести к смерти. Скорость свертывания крови у животных различных видов различна. Свертывание крови может происходить внутри кровеносных сосудов при повреждении их внутренней оболочки (интимы) или вследствие повышенной свертываемости крови. В этих случаях образуются внутри сосудистые тромбы, представляющие опасность для организма. Коагуляция крови обусловлена изменением физико-химического со стояния белка плазмы фибриногена, который при этом переход из растворимой формы в нерастворимую, превращаясь в фибрин. Тонкие и длинные нити фибрина образуют сеть, в петлях которой оказываются форменные элементы. Если вы пускаемую из сосуда кровь непрерывно помешивать метелочкой, то на ней осядут волокна фибрина. Кровь, из которой удален фибрин, называют дефибринированной. Она состоит из форменных элементов и сыворотки. Сыворотка крови - это плазма, в которой нет фибриногена и некоторых других веществ, участвующих в свертывании. Свертываться способна не только цельная кровь, но и плазма. Современная теория свертывания крови. В ее основу положена ферментативная теория А. Шмидта (1872 г.). Согласно последним данным, свертывание крови происходит в три фазы: 1 - образование протромбиназы, 2 - образование тромбина, 3 - образование фибрина. Кроме этого, выделяют предфазу и послефазу свертывания крови. В предфазу осуществляется так называемый сосудисто-тромбоцитарный, или микроциркуляторный, гемостаз. В послефазу входят два параллельных процесса: ретракция (уплотнение) и фибринолиз (растворение) кровяного сгустка. Гемостаз - это совокупность физиологических процессов, которые завершаются остановкой кровотечения при повреждении кровеносных сосудов. Сосудисто-тромбоцитарный, или микроциркуляторный, гемостаз - остановка кровотечения из мелких сосудов с низким кровяным давлением. Она слагается из двух последовательных процессов: спазм сосудов и формирование тромбоцитарной пробки. При травме рефлекторно происходит уменьшение просвета (спазм) мелких кровеносных сосудов. Рефлекторный спазм кратковременный. Более длительный спазм сосудов поддерживается сосудосуживающими веществами (серотонин, норадреналин, адреналин), которые выделяются тромбоцитами и поврежденными клетками тканей. Спазм сосудов приводит лишь к временной остановке кровотечения. Образование тромбоцитарной пробки имеет основное значение для остановки кровотечения в мелких сосудах. Тромбоцитарная пробка образуется благодаря способности тромбоцитов прилипать к чужеродной поверхности (адгезия тромбоцитов) и склеиваться друг с другом (агрегация тромбоцитов). Затем образовавшийся тромбоцитарный тромб уплотняется в результате сокращения специального белка тромбостенина, содержащегося в тромбоцитах. Остановка кровотечения при ранении мелких сосудов происходит у животных в течение 4 мин. Этот гемостаз в сосудах с низким давлением называется первичным. Он обусловлен длительным спазмом сосудов и механической закупоркой их агрегатами тромбоцитов. Вторичный гемостаз обеспечивает плотное закрытие поврежденных сосудов тромбом. Он предохраняет от возобновления кровотечения из мелких сосудов и служит основным механизмом защиты от кровотечения при повреждении сосудов мышечного типа. При этом происходит необратимая агрегация тромбоцитов и образование сгустка. В крупных сосудах гемостаз так же начинается с образования тромбоцитарной пробки, но она не выдерживает высокого давления и вымывается. В этих сосудах имеет место коагуляционный (ферментативный) гемостаз, осуществляемый в три фазы. Первая фаза. Образование протромбиназы - наиболее сложная и продолжительная. Различают тканевую и кровяную и тканевую протромбиназы. Образование тканевой протромбиназы совершается за 5-10 с, а кровяной 5-10 мин. Процесс образования тканевой протромбиназы начинается с повреждения стенок сосудов и окружающих их тканей и выделения из них в кровь тканевого тромбина, который представляет собой осколки клеточных мембран (фосфолипиды). В этом процессе принимают также участие вещества, содержащиеся в плазме, так называемые плазменные факторы: VII - конвертин, V - глобулин - акцелератор, Х – тромботропин и IV - катионы кальция. Образование тканевой протромбиназы служит пусковым механизмом для последующих реакций. Процесс образования кровяной протромбиназы начинается с активирования особого вещества плазмы – фактора ХII, или фактора Хагемана. В циркулирующей крови он находится в неактивном состоянии, что обусловлено наличием в плазме антифактора, препятствующего его активизации. При соприкосновении с шероховатой поверхностью антифактор разрушается, и тогда фактор Хагемана активируется. Шероховатой поверхностью служат обнажающиеся при повреждении кровеносного сосуда волокна коллагена. С активации фактора Хагемана начинается цепная реакция. Фактор ХII делает активным фактор ХI - предшественник плазменного тромбопластина и образует с ним комплекс, называемый контактным фактором. Под влиянием контактного фактора активируется фактор IХ - антигемофильньй глобулин В, который вступает в реакцию с фактором VIII антигемофильный глобулин А - и ионами кальция, образуя кальциевый комплекс. Последний оказывает сильное действие на кровяные пластинки. Они склеиваются, набухают и выделяют гранулы, содержащие тромбоцитарный фактор З. Контактный фактор, кальциевый комплекс и тромбоцитарный фактор образуют промежуточный продукт, который активирует фактор Х. Последний фактор на осколках клеточных мембран, тромбоцитов и эритроцитов (кровяной тромбопластин) образует комплекс, соединяясь с фактором V и ионами кальция. Этим завершается образование кровяной протромбиназы. Основным звеном здесь служит активный фактор Х. Вторая фаза. Образовавшаяся протромбиназа адсорбирует неактивный фермент плазмы протромбин (фактор II) на своей поверхности и превращает его в активный фермент тромбин. Протромбин всегда присутствует в циркулирующей крови. Он синтезируется в печени при участии филохинона (витамин К). Для его превращения в тромбин под влиянием протромбиназы необходимы факторы V, Х, ионы кальция и факторы тромбоцитов 1 и 2. Вторая фаза образование тромбина протекает за 2-5 с. Третья фаза. Происходит образование нерастворимого фибрина из растворимого белка плазмы фибриногена (фактор 1). Этот процесс идет под влиянием тромбина при участии ионов кальция и факторов тромбоцитов в три этапа. Первый этап (протеолитический) - под влиянием тромбина от молекулы фибриногена отщепляются пептиды, и он превращается в золеобразный фибрин-мономер (профибрин). Второй этап (полимеризационный) - объедение растворимых молекул профибрина в молекулы фибрин-полимра. Полимеризация происходит без участия тромбина, но под влиянием ионов кальция. Третий этап идет при участии фактора Х - фибриназы тканей, тромбоцитов и эритроцитов. Фибриназа образует прочные пептидные связи между соседними молекулами фибрин-полимера, что цементирует фибрин, увеличивает его механическую прочность и устойчивость к фибринолизу. Таким образом, формированием нерастворимого фибрина завершается процесс образования кровяного тромба. Затем наступает послефаза свертывания крови, во время которой идут два процесса - ретракция и фибринолиз. Ретракция заключается в том, что образовавшийся тромб начинает уплотняться, сжиматься, в него выдавливается сыворотка. Этот процесс происходит под влиянием особых веществ - ретрактозимов, выделяемых кровяными пластинками. Благодаря ретракции тромб плотнее закупоривает поврежденный сосуд. Ретракция заканчивается через 2-3 ч после образования сгустка. Одновременно с ретракцией, но с меньшей скоростью начинается фибринолиз - расщепление фибрина, который составляет основу тромба. Назначение фибринолиза: восстановить просвет кровеносного сосуда, закупоренного сгустком. Расщепление фибрина осуществляется протеолитическим ферментом фибронолизином, или плазмином, который находится в плазме в виде профермента профибринолизина, или плазминогена. Его активация осуществляется при помощи веществ, содержащихся в крови и тканях. При недостатке или отсутствии хотя бы одного из указанных факторов плазмы или тромбоцитов свертывание крови сильно замедляется либо становится совершенно невозможным. С давних времен известно наследственное заболевание людей - гемофилия, при котором резко понижена свертываемость крови. Оно встречается почти исключительно у мужчин, но передают его женщины. Оказалось, что гемофилия обусловлена дефицитом VIII и I факторов, которые названы антигемофильными. Подобное заболевание, связанное с нарушением синтеза одного из факторов плазмы, участвующего в образовании протромбиназы, встречается и у собак. Описаны случаи наследственной кровоточивости свиней. Больные поросята истекают кровью от ничтожных ранений и погибают. Болеют свиньи обоих полов, передают болезнь и самки, и самцы. В начале 20-х годов нашего века в Северной Америке была зарегистрирована массовая гибель крупного рогатого скота от нарушения свертываемости крови. Было замечено, что к этому заболеванию приводит кормление гниющим сеном или силосом из медового клевера донника. Добавление в корм люцерны (травы, богатой витамином К) предохраняет от заболевания. Впоследствии установили, что плохо силосованный гниющий медовый клевер содержит токсическое вещество, разрушающее филохинон (витамин К). Это вещество было выделено, изучены его строение, свойства и названо дикумарином. Дикумарин и его синтетические производные (неодикумарин, пелентан и др.) используют в клинике в качестве антикоагулянтов. Свертывание крови, взятой из кровеносного сосуда, можно предотвратить добавлением к ней следующих веществ: 1) солей (лимоннокислого, щавелевокислого, фтористого, сернокислого или углекислого натрия), которые связывают и удаляют из плазмы ионы кальция; 2) гепарина - препарата, полученного из печени; 3) пептонов, стимулирующих образование гепарина; 4) гирудина - противосвертывающего вещества, выделяемого слюнными железами пиявок.
Противосвертывающая система. Не менее важную роль в организме, чем свертывающая система, играет система противодействия свертыванию крови - противосвертывающая. Она является важным фактором в предупреждении внутрисосудистой коагуляции крови и растворении образовавшихся сгустков. Две системы - свертываювющая и антисвертывающая - находятся в организме в постоянной взаимосвязи и взаимодействии. В состав противосвертывающей системы входит ряд веществ: антипротромбопластин - плазменный ингибитор фактора Хагемана; антитромбопластины, действие которых направлено против образования тканевой и кровяной протромбиназы (антикефалин, липоидный ингибитор и др.). К ингибиторам фазы превращения протромбина в тромбин относят: гепарин - антикоагулянт с многообразным действием, тормозит действие образовавшейся протромбиназы, препятствует образованию протромбиназы и угнетает фазу формирования фибрина, постоянно образуется в тучных клетках тканей и базофилах крови, особенно богаты им печень, легкие, мышцы; антиконвертин - игибитор фактора VII; ингибитор фактора 1. Антитромбины - вещества, инактивирующие и разрушающие тромбин. Быстрое исчезновение тромбина после завершения свертывания крови обусловлено наличием в крови антитромбинов: антитромбин 1 - фибрин, который адсорбирует на своей поверхности значительное количество образовавшегося тромбина; антитромбин II - образует комплекс с гепарином, в его присутствии увеличивается количество тромбина, оседающего на фибрине, препятствует действию тромбина на фибриноген; антитромбин III - ускоряет распад тромбина; антитромбин III - блокирует активность тромбина.
Фибринолитическая система. В составе белков плазмы крови найдены вещества, растворяющие образовавшийся фибрин. К ним относится фибринолизин, или плазмин, находящийся в плазме в неактивном состоянии в форме профибринолизина, или плазминогена. Под действием активаторов фибринокиназ, содержащихся во многих тканях, профибринолизин переходит в активную форму - фибринолизин. Активаторы профибринолизина появляются в плазме после усиленной физической работы, при болевом раздражении, в случае острой интоксикации, при эмоциях и т. д. Регуляция свертывания крови. Постоянное взаимодействие свертывающей и противосвертывающей систем находится под контролем ней нейро-гуморальных механизмов. Если возникает необходимость в образовании тромба, чтобы прекратить кровотечение, усиливается действие свертывающей системы, и наоборот, когда появляется опасность образования сгустков в сосудах, активируется противосвертывающая система.
Боль, температурные воздействия, раздражение симпатических нервов, эмоции страха, ярости неизменно вызывают быстро наступающее ускорение свертывания крови. Во всех этих случаях возбуждена симпатическая нервная система и в кровь поступают большие количества норадреналина и адреналина. Усиленное их выделение и служит действующим началом, так как они из стенок кровеносных сосудов, освобождают тромбопластины, способствующие быстрому образованию тканевой протромбиназы; кроме того, адреналин прямо в кровотоке активирует фактор Хагемана, начинающий процесс образования кровяной протромбиназы. Адреналин усиливает освобождение тромбопластинов из форменных элементов крови тромбоцитов и эритроцитов, а также активирует тканевые липазы, усиливающие расщепление жиров и поступление в кровь жирных кислот, обладающих тромбопластической активностью. Раздражение блуждающего нерва или внутривенное введение ацетилхолина также вызывает выделение из стенок кровеносных сосудов тромбопластинов, и скорость свертывания крови увеличивается. Таким образом, в процессе эволюции сформировалась одна защитно-приспособительная реакция ускорение свертывания крови. Обычно после прекращения действия раздражителя на организм ускоренное свертывание крови – гиперкоагулемия - сменяется замедлением свертывания крови - гипокоагулемией вследствие значительного расходования факторов свертывания. Гипокоагулемия всегда вторична, то есть наступает после гиперкоагулемии. Можно и условнорефлекторно влиять на свертываемость крови. Так, путем многократного сочетания условного раздражителя, например звонка с болевым, вырабатывается условный рефлекс на звон и включение одного лишь звонка будет ускорять коагуляцию крови. Условнорефлекторный механизм изменения свертывания крови имеет исключительное значение, так как обеспечивает подготовку организма к защите от кровопотери. В регуляции свертывания крови различают две фазы: рефлекторную (кратковременную) и рефлекторно-гуморальную (более продолжительную). Начальным процессом ускорения свертывания крови при болевом раздражении служит поступление нервных импульсов из центральной нервной системы к кроветворным органам и депо крови. Это вызывает увеличение количества кровяных пластинок в периферической крови, главным образом за счет депонированной крови из печени, селезенки, кожи. Одновременно происходит активация плазменных факторов свертывания крови. Все это создает условия для быстрого образования протромбиназы. Затем включается и гуморальное звено, поддерживающее и продолжающее активирование свертывающей системы. Параллельно с этим снижается активность антисвертывающей системы. Мобилизация противосвертывающей системы происходит при медленном внутривенном введении крысам умеренных доз тромбопластина или тромбина, и внутрисосудистые тромбы не образуются. Если же тромбин вводить животным в состоянии наркотического сна или при местной анестезии, быстро наступает свертывание циркулирующей крови. Очевидно, в кровеносных сосудах имеются хеморецепторы, реагирующие на повышенное количество тромбина, и рефлекторным путем усиливается вы деление противосвертывающих веществ. Следовательно, свертывание крови процесс не автономный, он находится под регулирующим влиянием центральной нервной системы.
ГРУППЫ КРОВИ Агглютинация. В начале ХХ века было открыто явление агглютинации (склеивание) эритроцитов. Агглютинация наступает в результате взаимодействия содержащихся в эритроцитах антигенов агглютиногенов - и имеющихся в плазме антител - агглютининов. Явление агглютинации лежит в основе определения групп крови. В 1901 г. К. Ландштейнер открыл в человеческих эритроцитах два агглютинируемых фактора, которым дали название агглютиноген А и агглютиноген В. Оказалось, что в крови одних людей совсем нет агглютиногенов (группа 0), в крови других содержится только агглютиноген А (группа I, а у третьих - только агглютиноген В (группа III). Таким образом, К. Ландштейнер выделил три группы крови. Впоследствии К. Янский открыл IV группу крови, эритроциты которой содержат оба агглютиногена - А и В. В плазме крови было соответственно открыто два агглютинирующих агента - агглютинин и агглютинины. В крови каждого человека никогда не встречаются одновременно агглютиноген А с агглютинином с и агглютиноген в с агглютинином, поэтому в организме агглютинации собственных эритроцитов не происходит. Реакция агглютинации может произойти при переливании крови от одного человека другому. В прошлом попытки переливания крови нередко вызывали тяжелые заболевания и гибель вследствие агглютинации и последующего гемолиза чужеродных эритроцитов. Агглютинация наступает при взаимодействии одноименных агглютинина и агглютиногена А с А, В с В. Кроме того, для нее необходимо достаточное количество агглютининов. Кровь донора (Дающего кровь) переливают реципиенту медленно и в небольших количествах, поэтому в организм реципиента вводится, как правило, небольшое количество агглютининов. Следовательно, при переливании крови агглютинины донора не будут иметь значение, поэтому нужно обращать внимание на его агглютиногены, а у реципиента важно учитывать агглютинины. Людям с первой группой крови можно переливать кровь только первой группы. Кровь же первой группы можно переливать людям всех групп. Поэтому люди с первой группой крови являются универсальными донорами. Нельзя переливать кровь второй группы в третью и наоборот. Кровь четвертой группы можно вводить только четвертой, а она может получать кровь всех четырех групп (универсальный реципиент). Однако от этих правил в настоящее время отказались и переливают только одногруппную кровь. Одной из причин отказа от классических правил послужила необходимость массивного переливания крови при ряде хирургических операций. Кроме того, было обнаружено, что переливание крови первой группы людям с другими группами крови в 10- 20 % случаев вызывает тяжелые осложнения. В 1940 г. Ландштейнер и Винер открыли в эритроцитах людей еще один агглютиноген, который получил название резус-фактора. Исследователи вводили кровь обезьян (макака-резус) и кроликам и получали от них сыворотку, содержащую антитела против агглютиногенов макаки. Оказалось, что эта сыворотка дает резко положительную реакцию агглютинации эритроцитов не только макаки, но и большого количества людей (85 %) - резусные, у 15 % обследованных этот антиген не был обнаружен - резус-отрицательные. После переливания резус-положительной крови резус-отрицательному человеку у послед него образуются специфические антитела к резус-антигену. Поэтому повторное введение резус-положительной крови может вызвать у него агглютинацию эритроцитов и тяжелый гемотрансфузионный шок. Особое значение имеет образование резус-антител у резус-отрицательных матерей, вынашивающих резус-положительный плод. При этом резус-фактор плода диффундирует через плаценту в кровь матери и вызывает образование специфических антирезусных веществ. Последние проникают через плаценту в кровь плода и вызывают агглютинацию и гемолиз его эритроцитов. У ребенка развивается тяжелое гемолитическое заболевание, и только полная замена крови может предотвратить смерть новорожденного. К настоящему времени в эритроцитах человека найдено более 200 различных агглютиногенов, 140 из которых объединены в 20 систем. Определение групп крови имеет большое практическое значение, так как дает возможность проводить переливание крови без каких-либо последствий. Группы крови сельскохозяйственных животных. В эритроцитах сельскохозяйственных животных обнаружено большое количество антигенных факторов. Антигены, обусловливающие группы крови, обозначают заглавными буквами латинского алфавита (А, В, С,..., Р, К и т. д.) в соответствии с международной номенклатурой. Полное написание формулы группы крови учитывает как антигены (А, В и т. д.), так и антитела. Однако у большинства видов сельскохозяйственных животных при огромном богатстве антигенных факторов в эритроцитах очень мало, часто совсем нет естественных антител в плазме крови. Поэтому о группах крови животных делают заключение только по антигенной характеристике эритроцитов. Разнообразные сочетания антигенов создают десятки и сотни групп крови. Антигены, наследование которых взаимообусловленно, составляют систему групп крови. Наличие у животных антигенных факторов в эритроцитах определяют по их реакции с соответствующими антителами. Антитела, которые находятся в плазме крови неиммунизированных животных, называют нормальными или естественными. Однако основную массу антигенных факторов у сельскохозяйственных животных определяют с помощью антител, обнаруженных в плазме крови после предварительной иммунизации животных, то есть введения им эритроцитов того же вида. Наиболее изучены группы крови крупного рогатого скота и cвиней. С помощью иммунных специфических антисывороток у крупного рогатого скота изучены и идентифицированы 100 антигенных факторов, объединенных в 12 систем. Особенно обширна система В, в нее входит 50 антигенных факторов. Количество известных групп крови в системе В составляет несколько сотен, в системе С - 35, в системе А - 10 и т. д. Для антигенов групп крови крупного рогатого скота не хватило букв алфавита, поэтому стали использовать тот же алфавит с первой буквы, но с надстрочными индексами: А’, В’, С’ и т. д. У свиней обнаружены 50 антигенов, образующих 14 систем. К наиболее простым системам групп крови свиней относят системы В, У, 1, обусловливаемые двумя аллелями (в каждой системе потри группы крови). Более сложны в иммуногенетическом отношении полиаллельные системы АО, Е, Н, К и др. У овец найдено семь систем групп крови; наибольшее количество антигенных факторов в системе В (52), наименьшее в системах А и О. Интересно, что при наличии системы В у животных обнаруживают наиболее низкую активность фермента аденозинтрифосфатазы и наименьшее содержание калия в эритроцитах. У лошадей от крыто 10 естественных агглютиногенов, с помощью иммунизации у них удалось получить еще 19 агглютиногенов, а агглютинины в плазме выявляют редко (до 6 %). Агглютиногены лошадей образуют восемь систем групп крови. В системах А, О, Р по четыре группы крови. Система С - наиболее сложная (восемь групп крови). Системы С, К, Т представлены одним антигенным фактором, имеющим два аллеля, обусловливающих две группы крови. У кур найдено 60 антигенных факторов, сгруппированных в 14 систем. В каждой системе известно по одному (системы К, Р), два и более 20 (система В) антигенов, от которых зависит групповая дифференциация этого вида. Все эти данные пока нельзя считать окончательными. Наибольшее практическое значение анализ групп крови имеет в скотоводстве для установления происхождения животных, особенно при определении происхождения телят в связи с широким применением искусственного осеменения, когда коров осеменяют спермой различных быков. Анализ групп крови используют для селекционных целей. Изучение межпородных различий по группам крови позволяет уточнить происхождение пород и генетические связи между ними, а также степень при менявшегося при выведении породы близкородственного разведения. Так как у животных отсутствует система крови, аналогичная системе человека, в ветеринарии нет определенных данных о ценности тех или иных доноров применительно к определенным реципиентам. Однократное переливание крови животному, как правило, безопасно при условии, что реципиенту кровь до этого не переливали. Однако перед переливанием всё же следует проверить совместимость крови донора и реципиента. Для животноводства существенное значение имеет изучение возможных генетических связей групп крови с хозяйственно полезными признаками сельскохозяйственных животных. Например, при наличии в крови крупного рогатого скота системы К регистрируют высокое содержание жира в молоке. Установлена корреляция между группами крови и жизнеспособностью, живой массой и яйценоскостью кур. Осуществляя селекцию под иммуногенетическим контролем, с помощью определения групп крови созданы высокопродуктивные линии кур.
КРОВЕТВОРЕНИЕ И РЕГУЛЯЦИЯ СИСТЕМЫ КРОВИ Кроветворение, или гемопоэз, - процесс образования, развития и созревания клеток крови. Различают эритропоэз - образование эритроцитов, лейкопоэз - образование лейкоцитов и тромбоцитопоэз образование тромбоцитов. Кроветворение - одна из наиболее рано возникающих функций организма. Во время большинство исследователей придерживаются унитарной теории кроветворения. Согласно этой теории, все форменные элементы крови имеют общее происхождение. Их родоначальником является крупная клетка - гемоцитобласт. Во внутри утробный период образование и развитие эритроцитов, зернистых лейкоцитов и мегакариоцитов происходит в печени. В конце внутриутробного периода кроветворение в печени прекращается и единственным органом образования этих клеток становится красный костный мозг. Согласно современным представлениям, предшественники лимфоцитов образуются в костном мозге. Оттуда часть клеток переносится в лимфоузлы и селезенку, где идет их дозревание и размножение. Это так называемые В-клетки. Другие клетки- предшественники поступают из костного мозга с кровью в вилочковую железу, или тимус. Они размножаются, проходят первую стадию созревания и становятся Т-лимфоцитами. Затем они снова переходят в кровь и направляются в лимфатические узлы. Форменные элементы крови живут недолго, поэтому непрерывно об разуются новые и разрушаются старые клетки. Разрушение эритроцитов идет тремя путями. Один из них – фрагментоз - разрушение только что вышедших из костного мозга, наиболее неустойчивых молодых эритроцитов вследствие механического разрушения при циркуляции по кровеносным сосудам. Значительная часть эритроцитов подвергается фагоцитозу клетками мононуклеарной фагоцитарной системы в печени и селезенке. Эти органы называют «кладбищем эритроцитов». Третий путь разрушения - гемолиз старых эритроцитов прямо в циркулирующей крови. Разрушение эритроцитов так же, как и образование, идет в очень больших количествах. Так, если общее количество эритроцитов в организме коровы составляет примерно 300 триллионов, а средняя продолжительность их жизни 100 дн., то 3 триллиона должно образовываться и разрушаться каждый день, или около 35 млн за каждую секунду. При разрушении эритроцитов освобождается гемоглобин, который распадается на гем и глобин. От гема отделяется железо. При помощи меченых атомов установлено, что железо сразу используется для образования гемоглобина. Избыток же его вступает в соединение со специфическими белками крови, образуя ферритин и гемосидерин. Последние служат резервом железа и откладываются в печени, селезенке и в меньших количествах в слизистой тонкого кишечника. Транспорт железа из кишечника, где всасывается железо пищи, происходит с помощью белка трансферрина. Разрушение и образование лейкоцитов также совершаются непрерывно. Местом их разрушения являются клетки мононуклеарной системы печени и селезенки. Регуляция кроветворения. Она осуществляется нейро-гуморальным путем. Впервые идею нервной регуляции кроветворения и перераспределения элементов крови выдвинул С. П. Боткин еще в 80-х годах прошлого века. В дальнейшем в основу понимания этих процессов была положена концепция И. П. Павлова о кортиковисцеральных взаимоотношениях, развитая К. М. Быковым и его учениками. Наличие в кроветворных органах интерорецепторов служит доказательством того, что эти органы включены в систему рефлекторных взаимоотношений. Это подтверждается многими факторами. Так, при длительном раздражении блуждающего нерва происходит перераспределение лейкоцитов в крови: увеличивается их содержание в сосудах желудочно-кишечного тракта и уменьшается в других сосудистых областях. Раздражение симпатического нерва приводит к противоположному эффекту. При активном пищеварении, мышечной работе, болевых раздражениях, эмоциональном возбуждении количество лейкоцитов в крови возрастает за счет выхода их из лимфоузлов и синусов костного мозга. Такой лейкоцитоз называется перераспределительным. Он может быть вызван условнорефлекторным путем, что свидетельствует об участии в этих процессах коры больших полушарий. Экспериментальные и клинические данные показывают, что особое значение в регуляции кроветворения и перераспределения элементов крови имеет гипоталамическая область промежуточного мозга, осуществляющая свое действие через гипофиз и центры вегетативной нервной системы. На кроветворение оказывают влияние и железы внутренней секреции. Так, оно усиливается соматотропным и адренокортикотропным гормонами передней доли гипофиза, надпочечников, щитовидной железы. Мужские половые гормоны усиливают эритропоэз, а женские половые гормоны (эстрогены) тормозят его. По-видимому, этим отчасти объясняется более высокое количество эритроцитов у самцов, чем у самок. Нервные и эндокринные влияния осуществляются за счет специфических посредников гемопоэтинов. Эритропоэтины стимуляторы эритропоэза. Они образуются в печени, селезенке, но главным образом в почках. После удаления почек эритропоэтинов не обнаруживают, поэтому почки считают главным местом их образования. Они являются полипептидами относительно небольшой молекулярной массы. Их количество увеличивается при кислородном голодании, вызванном различными причинами (потерей крови, разрушением эритроцитов под влиянием некоторых ядов, продолжи тельным пребыванием на большой высоте и т. д.). Образование лейкоцитов стимулируется лейкопоэтинами, которые появляются у животных после быстрого удаления из крови большого количества лейкоцитов. Среди лейкопоэтинов выделяют нейтрофило-, базофило-, эозинофило-, моноцито- и лимфоцитопоэтины. Стимулирующее влияние на лейкопоэз оказывают продукты распада самих лейкоцитов и тканей, возникающие при их повреждении, нуклеиновые кислоты, микробы и их яды, а также некоторые гормоны. Так, под влиянием гормонов передней доли гипофиза (соматотропного и адренокортикотропного) увеличивается количество нейтрофилов и уменьшается число эозинофилов в крови. Все эти вещества действуют на лейкопоэз не прямо, а за счет лейкопоэтинов. В плазме крови найдены также тромбоцитопоэтины. Под их влиянием через несколько часов после острой кровопотери количество кровяных пластинок может увеличиться вдвое. Химическая природа, место образования, механизм действия этих веществ еще не установлены. Кроме того, для нормального созревания эритроцитов необходимы витамины (цианкобаламин, пиридоксин, фолиевая кислота). Цианкобаламин (витамин B12), так называемый внешний фактор кроветворения, поступает в организм с кормом. Он всасывается и усваивается только в том случае, если слизистая оболочка пилорической части желудка выделяет особое вещество — так называемый внутренний фактор кроветворения, или фактор Кэсла. При отсутствии этого вещества нарушается всасывание цианкобаламина и образование эритроцитов тормозится. Пиридоксин (витамин В6) необходим для синтеза гема. Фолиевая кислота, содержащаяся в растительных продуктах, нужна для синтеза нуклеиновых кислот и глобина в ядерных предстадиях эритроцитов. Рибофлавин (витамин В2) участвует в процессе формирования липидной стромы эритроцитов, пантотеновая кислота — в синтезе фосфолипидов. Для эритропоэза нужен также витамин С. Он усиливает всасывание железа из кишечника, способствует образованию тема, стимулирует действие фолиевой кислоты.
Кроме витаминов, животные должны получать с кормом достаточное количество белков и минеральных веществ. Особое значение для эритропоэза имеют такие элементы, как железо, медь, марганец, кобальт. Следовательно, полноценное кормление — необходимое условие образования и созревания эритроцитов и других форменных элементов крови.
Возрастные особенности системы крови. Возрастные изменения морфологического состава крови отчетливо проявляются у всех видов животных. В очень раннем возрасте функция кроветворения неустойчива и легко нарушается, что обусловлено недостаточным развитием нервной системы. С возрастом интенсивность гемопоэза постепенно уменьшается. При старении в костном мозге уменьшается количество кроветворных элементов, которые замещаются жировыми клетками. В лимфоузлах возникают атрофические изменения, ведущие к фиброзу. Возрастные изменения имеют видовые особенности. Так, у крупного рогатого скота и лошадей количество эритроцитов в молодом возрасте очень большое, затем в течение нескольких месяцев, а иногда 1—2 лет, оно постепенно уменьшается. По истечении этого срока количество эритроцитов вновь несколько увеличивается и долго сохраняется на одном уровне. Однако общий объем эритроцитарной массы с возрастом изменяется мало. Это обусловлено тем, что у молодых животных эритроциты имеют меньший размер, чем у взрослых. Возрастные изменения содержания гемоглобина соответствуют изменениям количества эритроцитов.
У свиней эти изменения носят несколько иной характер. У поросят, начиная с седьмого дня после рождения и до девяти месяцев, число эритроцитов в крови постепенно возрастает. Но так как молоко свиноматок очень бедно железом, то, несмотря на увеличение числа эритроцитов, количество гемоглобина в крови падает, если поросята дополнительно не получают препараты железа.
Дата добавления: 2015-12-15 | Просмотры: 1282 | Нарушение авторских прав 12 |