|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Концентрационный и электрический градиентыДля того, чтобы понять, как и почему возникает возбуждение в нервных или мышечных клетках, необходимо прежде всего, уяснить основные правила обмена веществами между клеткой и окружающей её средой, поскольку ионы и небольшие молекулы одновременно растворены в водной среде клетки и во внеклеточном пространстве, где их концентрация отличается от внутриклеточной. В среде биологов иногда говорят, что для исследования любой биологической проблемы Бог создал идеальный организм. Эксперименты, положенные в основу мембранной теории, были проведены в 40-х годах ХХ века на гигантских аксонах кальмара.
Как видно из таблицы, концентрация ионов калия внутри клетки намного больше, чем во внеклеточной жидкости, а концентрация ионов натрия и хлора, напротив, значительно больше во внеклеточной жидкости. Органические анионы представляют собой крупные молекулы, которые не проходят через клеточную мембрану. Корректно или нет делать какие-либо выводы о мембранах клеток теплокровных животных, тем более человека, изучая нервные клетки кальмара? Сравним их гигантские аксоны, например, с мышечными клетками теплокровных (таблица 4.2). Результаты измерений концентраций ионов в разных клетках животных, относящихся к разным видам, дают, разумеется и разные значения этих концентраций, но общим для всех клеток, у всех видов животных является одно: концентрация ионов калия всегда больше в клетке, а концентрация ионов натрия и хлора – во внеклеточной жидкости. Эта разница концентраций или концентрационный градиент является движущей силой для диффузии растворённых ионов в область меньшей концентрации или, в соответствии со вторым законом термодинамики, – к меньшему энергетическому уровню. Ещё раз взглянув на представленные в таблицах цифры, можно безошибочно предсказать, что катионы натрия должны диффундировать в клетку, а катионы калия – из неё. Однако не всё так просто, поскольку надо учесть и проницаемость клеточной мембраны для различных ионов, а она изменяется в зависимости от состояния активности клетки. В покое у плазматической мембраны открыты лишь ионные каналы для калия, через которые не могут проходить другие ионы. Значит ли это, что через мембрану покоящейся клетки ионы калия могут выходить беспрепятственно? Выходя из клетки, катионы калия уменьшают в ней количество положительных зарядов и одновременно увеличивают их количество на наружной поверхности мембраны. Остающиеся в клетке органические анионы начинают ограничивать дальнейший выход катионов калия, поскольку между анионами внутренней поверхности мембраны и катионами её наружной поверхности возникает электрическое поле и появляется электростатическое притяжение. Сама же клеточная мембрана оказывается поляризованной: на наружной её поверхности группируются положительные заряды, на внутренней – отрицательные.
Если подставить в уравнение значение констант и концентрацию ионов, то равновесный потенциал мембраны аксона кальмара для ионов калия окажется равным – 75 мВ (для мышечной мембраны теплокровных – -97 мВ). Это означает, что при такой трансмембранной разности потенциалов и при таких значениях внутри- и внеклеточной концентрации ионов калия их ток из клетки становится равным току в клетку. Если трансмембранная разность потенциалов сделается меньше, то ионы калия станут выходить из клетки до тех пор, пока не восстановится значение равновесного потенциала. У находящихся в состоянии покоя клеток глии мембрана пропускает только ионы калия, поэтому реальная трансмембранная разность потенциалов у них совпадает с расчётной, т.е. со значением равновесного потенциала для калия – 75 мВ. Но у большинства нейронов дело обстоит иначе, поскольку их мембрана в покое пропускает не только ионы калия, но в небольшом количестве и ионы натрия и хлора. В связи с этим трансмембранная разность потенциалов оказывается несколько меньше, чем равновесный калиевый потенциал, но незначительно, так как проницаемость для ионов калия в покое намного выше, чем для ионов натрия и хлора. Пользуясь уравнением Нернста нетрудно найти значение равновесных потенциалов для любых ионов (для натрия и хлора они приводятся в таблице 1). Равновесный потенциал для натрия равен + 55 мВ, а его концентрация во внеклеточной среде намного больше, чем в клетке; и то, и другое побуждает ионы натрия входить в клетку. Но в состоянии покоя клеточная мембрана не даёт им такой возможности: её проницаемость для ионов натрия крайне мала. 4.2. Активный транспор т Диффузия ионов должна уменьшать концентрационный градиент, но концентрационное равновесие означало бы для клетки гибель. Не случайно она более 1/3 своих энергетических ресурсов тратит на поддержание градиентов, на сохранение ионной асимметрии. Перенос ионов через клеточную мембрану против концентрационных градиентов является активным, т.е. энергозатратным видом транспорта, его обеспечивает натрий-калиевый насос. Это большой интегральный белок клеточной мембраны, который непрерывно выносит из клетки ионы натрия и одновременно закачивает в неё ионы калия. Этот белок обладает свойствами АТФ-азы, фермента, расщепляющего АТФ на внутренней поверхности мембраны, там же белок присоединяет три иона натрия. Освободившаяся при расщеплении молекулы АТФ энергия используется для фосфорилирования определённых участков белка-насоса, после которого меняется конформация белка и он выносит три иона натрия из клетки, но одновременно забирает снаружи и вносит в клетку два иона калия (рис. 4.1). Таким образом, за один цикл работы насоса выносятся из клетки три иона натрия, вносятся в неё два иона калия, а на эту работу тратится энергия одной молекулы АТФ. Именно так поддерживается высокая концентрация калия в клетке, а натрия – во внеклеточном пространстве. Если учесть, что и натрий, и калий являются катионами, т.е. несут положительные заряды, то суммарным итогом одного цикла работы насоса для распределения электрических зарядов является удаление одного положительного заряда из клетки. В результате такой деятельности мембрана становится чуть более отрицательной изнутри и поэтому натрий-калиевый насос можно считать электрогенным. За 1 секунду насос способен вынести из клетки около 200 ионов натрия и одновременно перенести в клетку приблизительно 130 ионов калия, а на одном квадратном микрометре мембранной поверхности может разместиться 100- 200 таких насосов. Кроме натрия и калия насос переносит в клетку против концентрационных градиентов глюкозу и аминокислоты; этот, как бы попутный транспорт, получил название: симпорт. Производительность натрий-калиевого насоса зависит от концентрации в клетке ионов натрия: чем больше она, тем быстрее работает насос. Если же концентрация ионов натрия в клетке понизится, то и насос уменьшит свою деятельность. Наряду с натрий-калиевым насосом в клеточной мембране существуют специальные насосы для ионов кальция. Они тоже используют энергию АТФ для выноса ионов кальция из клетки, в результате создаётся значительный концентрационный градиент кальция: вне клетки его значительно больше, чем в клетке. Это заставляет ионы кальция постоянно стремиться войти в клетку, но в состоянии покоя клеточная мембрана эти ионы почти не пропускает. Однако порой мембрана раскрывает каналы для этих ионов и тогда они играют очень важную роль в освобождении медиаторов или в активации некоторых ферментов. Таким образом, активный транспорт создаёт концентрационные и электрические градиенты, которые играют выдающуюся роль во всей жизни клетки. Дата добавления: 2015-12-15 | Просмотры: 792 | Нарушение авторских прав |
 Диаметр этих аксонов достигает 1 мм, их можно увидеть даже невооружённым глазом, в них легко вводить электроды, чтобы исследовать возникновение электрических сигналов – потенциалов действия. Именно на таком объекте работали основоположники мембранной теории, британские физиологи Алан Ходжкин и Эндрю Хаксли (Hodgkin A., Huxley A.), лауреаты Нобелевской премии 1963 года. Цитоплазма гигантских аксонов кальмара отличается от окружающей внеклеточной жидкости по концентрации некоторых ионов (таблица 4.1).
Диаметр этих аксонов достигает 1 мм, их можно увидеть даже невооружённым глазом, в них легко вводить электроды, чтобы исследовать возникновение электрических сигналов – потенциалов действия. Именно на таком объекте работали основоположники мембранной теории, британские физиологи Алан Ходжкин и Эндрю Хаксли (Hodgkin A., Huxley A.), лауреаты Нобелевской премии 1963 года. Цитоплазма гигантских аксонов кальмара отличается от окружающей внеклеточной жидкости по концентрации некоторых ионов (таблица 4.1). Равновесный потенциал – такое значение трансмембранной разности электрических зарядов, при котором ток ионов в клетку и из неё становится одинаковым, т.е. фактически ионы не перемещаются.
Равновесный потенциал – такое значение трансмембранной разности электрических зарядов, при котором ток ионов в клетку и из неё становится одинаковым, т.е. фактически ионы не перемещаются.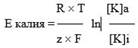 Таким образом, если мембрана готова пропустить какие-либо ионы, то направление ионного тока будут определять два обстоятельства: концентрационный градиент и действие электрического поля, причём концентрационный градиент может направлять ионы в одном направлении, а электрическое поле – в другом. Когда эти две силы уравновешиваются, ток ионов практически прекращается, поскольку количество входящих в клетку ионов становится равным количеству выходящих. Это состояние называется равновесным потенциалом (Е), а его значение можно вычислить с помощью уравнения Нернста (Nernst W., 1888): где R – газовая постоянная, Т – абсолютная температура (310 при температуре тела), z – валентность иона (для калия = 1), F – константа Фарадея, [K ]a – концентрация ионов калия вне клетки, [K]i – концентрация ионов калия в клетке.
Таким образом, если мембрана готова пропустить какие-либо ионы, то направление ионного тока будут определять два обстоятельства: концентрационный градиент и действие электрического поля, причём концентрационный градиент может направлять ионы в одном направлении, а электрическое поле – в другом. Когда эти две силы уравновешиваются, ток ионов практически прекращается, поскольку количество входящих в клетку ионов становится равным количеству выходящих. Это состояние называется равновесным потенциалом (Е), а его значение можно вычислить с помощью уравнения Нернста (Nernst W., 1888): где R – газовая постоянная, Т – абсолютная температура (310 при температуре тела), z – валентность иона (для калия = 1), F – константа Фарадея, [K ]a – концентрация ионов калия вне клетки, [K]i – концентрация ионов калия в клетке.