|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
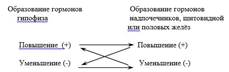
Регуляция образования гормоновКонцентрация гормонов в крови может изменяться в зависимости от скорости их образования. Существуют специальные сенсорные механизмы, которые следят либо за уровнем самого гормона в крови (1), либо за уровнем, регулируемых этим гормоном гомеостатических констант (2). При отклонении соответствующего параметра в сторону повышения или понижения происходит уменьшение или увеличение секреции гормона – это регуляция по принципу обратной связи. Например, повышение в крови уровня образующегося в коре надпочечников кортизола уменьшает выделение из гипофиза кортикотропина, который стимулировал секрецию кортизола. В результате снижения продукции кортикотпропина выделение кортизола из надпочечников уменьшается и соответственно нормализуется уровень этого гормона в крови. Пониженный уровень кортизола приводит к увеличению образования кортикотропина, он стимулирует деятельность коры надпочечников, выделение кортизола в кровь повышается. По такому же принципу гипофиз взаимодействует со щитовидной и половыми железами: при высоком уровне их гормонов в крови в гипофизе уменьшается синтез тропных гормонов (соответственно тиреотропного и гонадотропных), а низкий уровень тироксина или половых стероидов, наоборот, ведёт к увеличенной секреции тиреотропного или гонадотропных гормонов:
В 1928 году Эрнст и Берта Шаррер (Scharrer E. B.) впервые обнаружили явление нейросекреции – способность нервных клеток синтезировать и выделять определённые вещества. Дальнейшие исследования позволили создать концепцию, согласно которой некоторые нейроны могут действовать как нейроэндокринные преобразователи, трансформирующие кратковременные нервные импульсы в длительные гуморальные влияния на ткани организма. В 1955 году Джеффри Харрис (Harris G.) сумел объяснить назначение необычной сосудистой связи между гипоталамусом и гипофизом, получившей название портальной (т.е. воротной) системы. Густая сеть капилляров гипоталамической области собирается в т.н. портальные вены, они доставляют кровь к передней доле гипофиза, где снова распадаются на множество мелких капилляров (рис.12.2). Харрис, на основе проведённых экспериментов, показал, что продукты нейросекреции клеток гипоталамуса доставляются по портальной системе в аденогипофиз (передняя доля гипофиза, состоящая из железистой ткани) и регулируют образование его гормонов.
Первые из них назвали либеринами или рилизинг-гормонами (от лат. libero или англ. release – освобождать), а вторые – ингибирующими гормонами или статинами. Известные к настоящему времени либерины и статины представлены в таблице 12.1 вместе с гормонами аденогипофиза, на образование которых они влияют. Следует отметить, что соматостатин тормозит ещё и продукцию тиреотропина, а тиреотропин-либерин стимулирует секрецию пролактина, тогда как тормозит её не нейросекрет, а хорошо известный нейротрансмиттер дофамин. Чтобы понять функциональный смысл взаимоотношений гипоталамуса и гипофиза, следует сопоставить два обстоятельства. Во-первых, в гипоталамус, как уже рассматривалось в главе 11, поступает необходимая информация о состоянии гомеостаза и внутренних органов, он имеет двусторонние связи с различными подкорковыми структурами и корой, что позволяет ему координировать всю регуляцию вегетативных функций. Во-вторых, передняя доля гипофиза – классическая железа внутренней секреции, образованная несколькими разновидностями железистых клеток, она выделяет гормоны, которые контролируют деятельность других важнейших эндокринных желёз: коры надпочечников (кортикотропин), щитовидной (тиреотропин) и половых.(два гонадотропных гормона: лютеотропин и фоллитропин). Ещё два гормона гипофиза – соматотропин и пролактин также играют важную роль в эндокринной регуляции функций. Таким образом, выделяя в капилляры портальной системы свои нейросекреты – гипофизотропные гормоны, гипоталамус держит под контролем большинство желёз внутренней секреции. Однако отношения между гипоталамусом и гипофизом не следует рассматривать, как улицу с односторонним движением: повышение в крови уровня гипофизарных гормонов и гормонов зависимых от него желёз заставляет гипоталамус уменьшать секрецию либеринов и/или увеличивать выделение статинов. Дата добавления: 2015-12-15 | Просмотры: 872 | Нарушение авторских прав |
 Если же секреция гормонов зависит от управляемого параметра, как, например, в случае образования гормонов поджелудочной железы, контролирующих уровень глюкозы в крови, то схему регуляции можно изобразить так:
Если же секреция гормонов зависит от управляемого параметра, как, например, в случае образования гормонов поджелудочной железы, контролирующих уровень глюкозы в крови, то схему регуляции можно изобразить так: По такой же схеме регулируется продукция двух гормонов, контролирующих уровень кальция в крови: тирокальцитонин выделяется особыми клетками щитовидной железы в большем количестве, если нужно понизить уровень кальция в крови, а когда кальция в крови становится мало, активируется образование паратгормона в околощитовидных железах: и в том, и в другом случае результатом изменения секреции гормонов будет нормализация уровня кальция в крови. Регуляция по принципу обратной связи осуществляется быстро: темп секреции того или иного гормона может измениться уже за несколько минут, благодаря чему константы гомеостаза удерживаются в относительно узких пределах.
По такой же схеме регулируется продукция двух гормонов, контролирующих уровень кальция в крови: тирокальцитонин выделяется особыми клетками щитовидной железы в большем количестве, если нужно понизить уровень кальция в крови, а когда кальция в крови становится мало, активируется образование паратгормона в околощитовидных железах: и в том, и в другом случае результатом изменения секреции гормонов будет нормализация уровня кальция в крови. Регуляция по принципу обратной связи осуществляется быстро: темп секреции того или иного гормона может измениться уже за несколько минут, благодаря чему константы гомеостаза удерживаются в относительно узких пределах.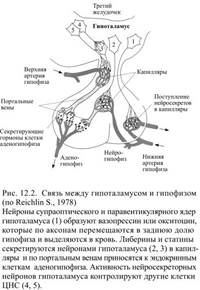 12.3. Роль гипоталамуса в регуляции образования гормонов передней доли гипофиза (гипоталамо-аденогипофизарная система)
12.3. Роль гипоталамуса в регуляции образования гормонов передней доли гипофиза (гипоталамо-аденогипофизарная система) Химическую природу нейросекретов, выделяющихся в чрезвычайно малом количестве, удалось обнаружить лишь в 1969 сразу двум группам исследователей, которыми руководили Роже Гиймен (Guilleman R.) и Эндрю Шелли (Schally A.), получившие за это открытие Нобелевскую премию в 1977 году. Выделяемые сравнительно небольшими, т.н. парвоцеллюлярными гипоталамическими нейронами вещества оказались пептидами, одни из которых стимулировали, а другие тормозили образование гормонов передней доли гипофиза.
Химическую природу нейросекретов, выделяющихся в чрезвычайно малом количестве, удалось обнаружить лишь в 1969 сразу двум группам исследователей, которыми руководили Роже Гиймен (Guilleman R.) и Эндрю Шелли (Schally A.), получившие за это открытие Нобелевскую премию в 1977 году. Выделяемые сравнительно небольшими, т.н. парвоцеллюлярными гипоталамическими нейронами вещества оказались пептидами, одни из которых стимулировали, а другие тормозили образование гормонов передней доли гипофиза.