|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Роль миндалин в образовании мотиваций
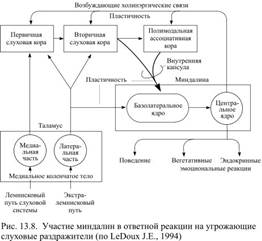
Есть два важных источника афферентной информации для миндалин: сенсорные ядра таламуса и кора, преимущественно вторичные слуховые области и полимодальные ассоциативные поля (рис. 13.8). При действии внешних стимулов сенсорная информация раньше поступает от таламуса и вызывает примитивную реакцию, на фоне которой приходит информация, уже переработанная в коре. Оба афферентных потока направляются к базолатеральным ядрам миндалин, где пришедшие сигналы перерабатываются и передаются нейронам центрального ядра миндалин. От клеток центрального ядра начинаются эфферентные пути миндалин, которые направляются к латеральному гипоталамусу и стволу мозга. При возникновении страха активация миндалин приводит к последующему быстрому повышению частоты сокращений сердца и артериального давления, учащению дыхания, выделению гормонов и другим проявлениям симпатоадреналовой реакции и стресса. Такими же последствиями сопровождается раздражение центрального ядра миндалины электрическим током, тогда как его разрушение приводит к исчезновению вегетативных реакций в ответ на действие стимулов, обычно вызывающих страх. Помимо этого существует эфферентный путь от миндалин к коре, в особенности к орбитофронтальной области и поясной извилине, этот путь важен для осознания переживаемых ощущений. Кроме того, ядра миндалин имеют реципрокные соединения с гиппокампом, а также получают афферентную проекцию от гипоталамуса, хотя она и уступает по мощности эфферентным влияниям миндалин на гипоталамус. И ещё необходимо отметить связь миндалин с мезолимбической системой, прежде всего с прилегающим ядром (n. accumbens). У кроликов можно выработать условный рефлекс на нейтральный звуковой стимул, подкрепляя его болевым действием электрического тока. Болевое раздражение всегда сопровождается повышением частоты сокращений сердца, артериального давления, учащением дыхания и т.п. Когда образуется условный рефлекс, такая же реакция наблюдается и на прежде нейтральный звуковой раздражитель, который начинает вызывать у животного состояние страха. Если теперь удалить или разрушить миндалины, либо ввести в них специфические блокаторы синаптической передачи – реакция страха исчезнет. Таким образом, не только врождённые, но и приобретённые мотивации страха связаны с обязательным участием миндалин. Роль миндалин не сводится только к мотивации страха. Хотя при их повреждении не исчезают совсем мотивации голода или жажды, но в пищевом и питьевом поведении наступают заметные перемены. Подопытные животные перестают отличать предпочитаемую раньше вкусную пищу от невкусной и постоянно берут в рот даже несъедобные предметы. У них пропадает врождённая или выработанная до операции разборчивость в питье, и, например, подопытные крысы начинают пить растворы с неприятным для нормальных животных запахом. У них изменяется половое поведение: животные становятся гиперсексуальными и готовы спариваться даже с представителями другого вида. Таким образом, значение миндалин становится особенно заметным в тех случаях, когда для формирования мотивированного поведения одновременно требуется учесть существование нескольких внешних факторов. Дата добавления: 2015-12-15 | Просмотры: 806 | Нарушение авторских прав |
 Если гипоталамус является важнейшей мотивирующей структурой мозга, активирующейся при изменении постоянства внутренней среды, то на внешние стимулы, вызывающие мотивацию или же изменяющие уже мотивированное поведение, раньше гипоталамуса откликаются нейроны миндалевидных ядер или миндалин мозга, которые относятся к лимбической системе и находятся в височных долях больших полушарий. Электрическое раздражение миндалин у людей чаще всего вызывает чувство страха и тревоги, двустороннее удаление миндалин у экспериментальных животных по-видимому лишает их чувства страха. Каков же механизм формирования такой мотивации? Поиск ответа на этот вопрос побудил к исследованию афферентных и эфферентных связей миндалин.
Если гипоталамус является важнейшей мотивирующей структурой мозга, активирующейся при изменении постоянства внутренней среды, то на внешние стимулы, вызывающие мотивацию или же изменяющие уже мотивированное поведение, раньше гипоталамуса откликаются нейроны миндалевидных ядер или миндалин мозга, которые относятся к лимбической системе и находятся в височных долях больших полушарий. Электрическое раздражение миндалин у людей чаще всего вызывает чувство страха и тревоги, двустороннее удаление миндалин у экспериментальных животных по-видимому лишает их чувства страха. Каков же механизм формирования такой мотивации? Поиск ответа на этот вопрос побудил к исследованию афферентных и эфферентных связей миндалин.