|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ТЕМА 2. ТКАНИ ВЫСШИХ РАСТЕНИЙМИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА РОССИЙСКОЙ ФЕДЕРАЦИИ ФГБОУ ВПО «БРЯНСКАЯ ГОСУДАРСТВЕННАЯ СЕЛЬСКОХОЗЯЙСТВЕННАЯ АКАДЕМИЯ»
Кафедра биологии, кормопроизводства, селекции и семеноводства
БОТАНИКА (учебно-методическое пособие для лабораторно-практических занятий Раздел: «Цитология, гистология и органография вегетативных органов»)
Брянск 2014 УДК ББК М - 47
Милехина, Н.В. Ботаника (учебно-методическое пособие для лабораторно-практических занятий. Раздел: цитология, гистология и органография вегетативных органов). Предназначено для подготовки студентов по направлениям 110400 «Агрономия», 110100 «Агрохимия и агропочвоведение», 110900 «Технология производства и переработки с/х продукции»; квалификация (степень) выпускника – бакалавр). Брянск. Издательство Брянской ГСХА, 2014. – 61 с. Учебно-методическое пособие предназначено для выполнения лабораторно практических работ по дисциплине «Ботаника» Раздел: цитология, гистология и органография вегетативных органов. Разработано в соответствии с требованиями примерной программы по дисциплине «Ботаника», квалификация (степень) выпускника – бакалавр.
Рецензент: доцент кафедры экологии и рационального природопользования БГУ, к.с.-х. наук Н.А. Сковородникова
Рекомендовано к изданию решением методической комиссии агроэкологического института от года
©Брянская ГСХА, 2014 © Н.В. Милехина, 2014
СОДЕРЖАНИЕ Тема 1. РАСТИТЕЛЬНАЯ КЛЕТКА И ЕЕ КОМПОНЕНТЫ…………………………………………………………………… Тема 2. ТКАНИ ВЫСШИХ РАСТЕНИЙ…………………….… 2.1 ОБРАЗОВАТЕЛЬНЫЕ ТКАНИ (МЕРИСТЕМЫ)………….. 2.2 ПОКРОВНЫЕ ТКАНИ……………………………………….. 2.3 ОСНОВНЫЕ ТКАНИ………………………………………… 2.4 МЕХАНИЧЕСКИЕ ТКАНИ………………………………..... 2.5 ПРОВОДЯЩИЕ ТКАНИ И КОМПЛЕКСЫ………………... Тема 3. ВЕГЕТАТИВНЫЕ ОРГАНЫ РАСТЕНИЙ………..…. 3.1 КОРЕНЬ И КОРНЕВАЯ СИСТЕМА…………………..… 3.2 ПЕРВИЧНОЕ СТРОЕНИЕ КОРНЯ………………………. 3.3. ВТОРИЧНОЕ СТРОЕНИЕ КОРНЯ……………………… 3.4. КОРНЕПЛОДЫ…………………………………………… Тема 4. ПОБЕГ И СИСТЕМА ПОБЕГОВ……………………… 4.1 НАРАСТАНИЕ И ВЕТВЛЕНИЕ ПОБЕГОВ……………. ТЕМА 5. СТЕБЕЛЬ- ОСЬ ПОБЕГА……………………………. 5.1 АНАТОМИЧЕСКОЕ СТРОЕНИЕ СТЕБЛЕЙ…………… 5.2 АНАТОМИЧЕСКОЕ СТРОЕНИЕ СТЕБЛЯ ОДНОДОЛЬНЫХ РАСТЕНИЙ…………………………………………… 5.3 АНАТОМИЧЕСКОЕ СТРОЕНИЕ СТЕБЛЕЙ ТРАВЯНИСТЫХ ДВУДОЛЬНЫХ РАСТЕНИЙ…………………………. 5.4 АНАТОМИЧЕСКОЕ СТРОЕНИЕ СТЕБЛЯ ДРЕВЕСНОГО РАСТЕНИЯ………………………………………………….. Тема 6. ЛИСТ. МОРФОЛОГИЯ И АНАТОМИЯ ЛИСТА……………………………………………………………… 6.1 МОРФОЛОГИЯ И КЛАССИФИКАЦИЯ ЛИСТЬЕВ…….. 6.2 АНАТОМИЯ ЛИСТА………………………………………. 6.3 АНАТОМИЧЕСКОЕ СТРОЕНИЕ ЛИСТЬЕВ ОДНОДОЛЬНЫХ И ДВУДОЛЬНЫХ РАСТЕНИЙ……………………..
Источники рисунков………………………………………….. Список литературы……………………………………..……. ТЕМА 1. РАСТИТЕЛЬНАЯ КЛЕТКА И ЕЕ КОМПОНЕНТЫ
Клетка – основная структурная единица одноклеточных, колониальных и многоклеточных растений. Клетки многоклеточных организмов разнообразны по размеру, форме и внутреннему строению. Различают две основные формы клеток: паренхимную и прозенхимную (рис.1).
Рис. 1 Растительные клетки: а,б – паренхимные клетки; в- прозенхимные: 1- ядро с ядрышками; 2- цитоплазма; 3- вакуоль; 4- клеточная стенка Паренхимные клетки - многогранники. Размер их примерно одинаков во всех направлениях, длина не более чем в два-три раза превышает ширину. Паренхимные клетки, где откладываются запасные питательные вещества, являются наиболее крупными. Прозенхимные клетки - вытянутые, их длина превышает ширину и толщину в 5-100 раз и более. Растительная клетка эукариотическая и имеет оформленное ядро. Основными особенностями растительных клеток является: - наличие пластид, - целлюлозопектиновая жесткая клеточная стенка, - хорошо развитая система вакуолей с клеточным соком, - отсутствие центриолей при делении. У молодых вновь образовавшихся клеток полость заполнена густой цитоплазмой. Цитоплазма и ядро составляют живое содержимое клетки – протопласт. В протопласте происходят все основные процессы обмена веществ (рис. 2).
Рис. 2 Схема строения растительной клетки (электронная микроскопия): 1- ядро; 2- ядерная оболочка (две мембраны - внутренняя и внешняя – и перинуклеарное пространство); 3- ядерная пора; 4- ядрышко (гранулярный и фибриллярный компоненты); 5- хроматин (конденсированный и диффузный); 6- ядерный сок; 7- клеточная стенка; 8- плазмалемма; 9- плазмодесмы; 10- эндоплазматическая агранулярная сеть; 11- то же, гранулярная; 12- митохондрия; 13- свободные рибосомы; 14- лизосома; 15- хлоропласт; 16- диктиосома аппарата Гольджи; 17- гиалоплазма; 18- тонопласт; 19- вакуоль с клеточным соком
В состав протопласта входят различные соединения: белки, нуклеиновые кислоты, липиды, углеводы, вода. Цитоплазма – это обязательная часть живой клетки, где происходят процессы клеточного обмена, исключая синтез нуклеиновых кислот, совершающийся в ядре. Основу цитоплазмы составляет матрикс или гиалоплазма. С гиалоплазмой связано движение цитоплазмы. Ядро – важнейшая структура, регулирующая всю жизнедеятельность клетки (обмен веществ, деятельность других органелл). В ядре хранится и воспроизводится наследственная информация, определяющая признаки данной клетки и всего организма в целом. Клетка без ядра быстро погибает. Обычно клетка имеет одно ядро шаровидной формы, окруженное цитоплазмой. Ядро является коллоидной системой вязкой консистенции с высоким содержанием ДНК, а также в значительном количестве и -РНК и р-РНК. Структура ядра одинакова у всех эукариотических клеток: ядерная оболочка, ядерный сок, хромосомно-ядрышковый комплекс. Многообразные функции цитоплазмы выполняют специализированные обособленные органеллы - рибосомы, пластиды, митохондрии, и диктиосомы. Рибосомы. Содержатся во всех клетках и осуществляют биосинтез белка. Каждая рибосома состоит из двух субъединиц: большой и малой. Рибосомы по своей структуре являются полужидкими образованиями и легко нанизываются на молекулу информационной РНК. Формирование субъединиц происходит в ядре, сборка в цитоплазме на молекуле и-РНК. Пластиды. Органеллы характерны только для растительных клеток. В них происходит первичный и вторичный синтез углеводов. По наличию пигментов (окраске) различают три типа пластид: зеленые хлоропласты, желто-оранжевые и красные хромопласты, бесцветные лейкопласты. Возможно взаимопревращение пластид. Обычно в клетке встречается только один тип пластид. Пластиды имеют двойную мембрану и заполнены матриксом. В нем имеется кольцевая ДНК и рибосомы. Хлоропласты – это органеллы фотосинтеза. Хлоропласты высших растений имеют двояковыпуклую форму (рис 3). Располагаются в клетках растений в постенном слое цитоплазмы таким образом, что одна из плоских сторон обращена к освещенной стенке клетки. Положение хлоропластов меняется в зависимости от освещенности: при прямом солнечном свете они перемещаются к боковым стенкам. Хлоропласты содержат пигменты: хлорофиллы и каротиноиды. Они покрыты двойной мембранной оболочкой, в которой заключена бесцветная мелкозернистая строма – матрикс.
Рис. 3 Хлоропласт: схема строения: 1- мембрана оболочки хлоропласта наружная; 2- то же, внутренняя;3- перипластидное пространство; 4- рибосомы; 5- нить пластидной ДНК; 6- матрикс; 7- грана; 8- тилакоид граны; 9- тилакоид стромы; 10- пластоглобула; 11- крахмальное зерно
Из складок внутренней мембраны образуются плоские мешки - тилакоиды в них локализованы пигменты. Дисковидные тилакоиды собраны в стопки – граны. Лейкопласты – бесцветные округлые пластиды, в которых накапливаются запасные питательные вещества, в основном крахмал. В состав лейкопластов входят: ДНК, рибосомы, ферменты, осуществляющие синтез и гидролиз запасных веществ. Хромопласты – пластиды оранжево красного и желтого цвета, образуются из лейкопластов и хлоропластов. Они встречаются в клетках лепестков, зрелых плодов, корнеплодах и в осенних листьях. Митохондрии - округлые или цилиндрические органеллы. Имеют двумембранное строение, внутри – бесструктурный матрикс. Внутренняя мембрана образует складки – кристы, которые в растительных клетках имеют вид трубочек. В матриксе имеются молекулы ДНК и рибосомы (рис. 4).
а – в объемном изображении; б- на срезе; 1- наружная мембрана; 2- внутренняя мембрана с кристами в виде трубочек; 3- матрикс; 4- перимитохондриальное пространство; 5- митохондриальные рибосомы; 6- нить митохондриальной ДНК
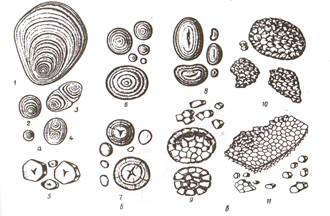
Митохондрии являются энергетическим центром клетки. В них проходит процесс внутриклеточного дыхания, т.е идет окисление органических веществ кислородом воздуха до простых неорганических веществ - углекислого газа и воды, при этом выделяется энергия АТФ. Диктиосомы – это органеллы, которые входят в состав аппарата Гольджи, который также включает пузырьки Гольджи. Они представляют собой стопки плоских округлых цистерн, ограниченных мембраной и заполненных матриксом. Цистерны диктиосом – последний участок многих обменных реакций. Здесь накапливаются, конденсируются и упаковываются вещества, подлежащие удалению из цитоплазмы. Они упаковываются в пузырьки и поступают в вакуоли. Клеточная стенка. Клетки растений в отличие от животных имеют твердые клеточные стенки. Они придают клеткам определенную форму, защищают их от внешних факторов, обеспечивают механическую прочность. Клеточные стенки бесцветны и легко пропускают солнечный свет. Стенки соседних клеток соединены пектиновой срединной пластинкой. В состав клеточной стенки входят полисахариды – пектин, гемицеллюлоза и целлюлоза. Они придают прочность и эластичность. Клеточная стенка создается протопластом. В процессе жизнедеятельности клетки целлюлозная клеточная стенка может претерпевать изменения: Одревеснение или лигнификация – это отложение в межмицеллярные промежутки лигнина. В результате возрастает твердость и прочность стенки, но уменьшается ее пластичность. Опробковение или суберинизация - отложение в клеточную стенку жироподобного аморфного вещества – суберина. При этом клеточные стенки становятся непроницаемыми для воды и газов. Клетки с опробковевшими клеточными стенками защищают растения от испарения. Кутинизация – отложение кутина – вещества, близкого к суберину, в поверхностных слоях наружных клеточных стенок и на их поверхности; образующаяся при этом пленка (кутикула) препятствует испарению. Минерализация – отложение в клеточных стенках солей кальция и кремнезема. Отложение кремнезема характерно для клеток эпидермы хвощей, осок и злаков. Ослизнение – превращение целлюлозы и пектина в слизи и близкие к ним камеди, которые при соприкосновении с водой способны сильно набухать. Явление можно наблюдать в кожуре семян, например у льна. Запасные питательные вещества. Растения в процессе жизнедеятельности накапливают продукты обмена веществ. Включения клетки представляют собой вещества, временно выведенные из обмена, или конечные его продукты. Запасные питательные вещества являются продуктами первичного обмена веществ, остальные группы – вторичного. Запасные питательные вещества накапливаются в клетках в течение вегетационного периода и используются частично зимой, весной и во время цветения. Широко распространено у растений отложение запасных жиров в виде липидных капель в цитоплазме. Наиболее богаты ими плоды и семена. Запасные белки (протеины) наиболее часто встречаются в виде алейроновых зерен (образуются при созревании семян из высыхающих вакуолей) в клетках семян бобовых, гречишных, злаков и других растений. Они имеют различную форму и размеры. Алейроновое зерно, окруженное тонопластом и содержащее аморфный белок альбумин, в который погружены белковые кристаллы глобулина, является сложным зерном (семена льна, подсолнечника, тыквы). Алейроновые зерна, содержащие только аморфный белок, называют простыми (бобовые, рис, кукуруза, гречиха). Крахмал – наиболее распространенное запасное вещество растений. Следует различать крахмал ассимиляционный (или первичный), запасной (или вторичный) и транзиторный. Ассимиляционный образуется в процессе фотосинтеза в хлоропластах из глюкозы. Запасной крахмал откладывается в лейкопластах (амилопластах) в виде крахмальных зерен (рис. 5).
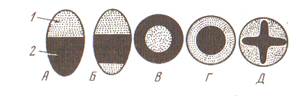
Рис. 5 Крахмальные зерна: а- из клеток клубней картофеля: 1- простое эксцентрическое; 2- простые концентрические; 3- сложное; 4- полусложное; б- простые зерна из клеток эндосперма: 5- кукурузы; 6- пшеницы; 7- ржи; 8- фасоли; в – сложные зерна из клеток эндосперма:9- овса; 10- риса; 11- гречихи
Крахмальные зерна бывают простые, сложные и полусложные. Простые зерна имеют один центр крахмалообразования, вокруг которого формируются слои крахмала (пшеница, рожь, кукуруза). У сложных зерен в одном лейкопласте несколько центров, имеющих свои собственные слои (рис, овес, гречиха). В полусложных зернах также несколько центров, но кроме слоев крахмала, возникших возле каждого центра, по периферии зерна имеются общие слои. В клубнях картофеля встречаются все три типа крахмальных зерен. Крахмальные зерна представляют собой сферокристаллы, состоящие из игольчатых кристаллов. Продуктами вторичного происхождения для растительных клеток служат соли щавелевой кислоты, дубильные вещества, алкалоиды, эфирные масла, смолы, оксалат кальция и др. Клетки многоклеточного организма, образовавшиеся путем митоза, обладают одинаковыми наследственными свойствами. Свойство клеток реализовывать всю генетическую информацию ядра, обеспечивающую их дифференцировку и развитие до целого организма, называют тотипотентностью. В многоклеточном организме клетки выполняют различные функции и имеют разное строение, приобретенное в ходе онтогенеза, или жизненного цикла. Жизненный цикл клетки - это развитие ее от момента возникновения в результате предшествующего деления до разделения на две новые клетки или до ее смерти. ТЕМА 2. ТКАНИ ВЫСШИХ РАСТЕНИЙ
Ткани – это группы клеток, сходных по строению, происхождению и приспособленных к выполнению одной или нескольких определенных функций. Наиболее часто различают шесть типов тканей: образовательные, или меристематические, и постоянные – покровные; основные; механические; проводящие; выделительные. У растений происходит постоянное деление клеток по типу митоза. В зависимости от происхождения различают первичные и вторичные меристемы.
2.1 ОБРАЗОВАТЕЛЬНЫЕ ТКАНИ (МЕРИСТЕМЫ)
Первичные меристемы. Из первичной образовательной (эмбриональной) ткани состоит зародыш семени. У взрослых растений первичные меристемы сохраняются лишь на самой верхушке стебля и вблизи кончика корня. Остальные клетки перестают делиться и превращаются в клетки постоянных тканей. Вторичные меристемы образованы или первичными меристемами, или постоянными тканями. По положению в растении выделяют: верхушечные (апикальные), боковые (латеральные), вставочные (интеркалярные) меристемы. Верхушечные меристемы находятся на полюсах зародыша– кончике корешка и почечке. Они обеспечивают рост корня и побега в длину. Меристемы являются первичными и образуют конусы нарастания корня и побега. Боковые меристем ы располагаются по окружности осевых органов и обеспечивают утолщение корня и стебля. Первичные боковые меристемы – прокамбий и перицикл - возникают под апексами. Вторичные – камбий и феллоген (пробковый камбий формируются позднее из меристем или постоянных тканей путем их дифференцировки. Из прокамбия и камбия образуются проводящие ткани, из феллогена – пробка. Вставочные меристемы располагаются в основаниях междоузлий, черешков листьев. Особенно хорошо заметны у злаков и обеспечивают поднятие хлебов после полегания. Раневые меристемы образуются при повреждении тканей и органов. Они образуют каллус – плотную ткань беловатого или желтоватого цветов, состоящую из паренхимных клеток, расположенных беспорядочно. Из каллуса может возникнуть любая ткань или орган растения. В нем могут закладываться придаточные корни и почки. Каллус образуется из раневых меристем при прививках, обеспечивая срастание привоя с подвоем; в основании черенков.
2.2 ПОКРОВНЫЕ ТКАНИ
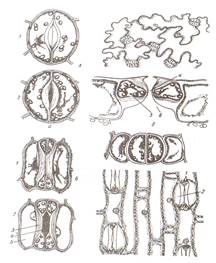
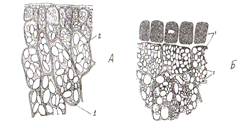
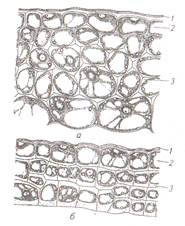
Покровные ткани расположены снаружи всех органов растений на границе с внешней средой и предохраняет их от неблагоприятных воздействий окружающей среды. Эпиблема (ризодерма) – это первичная однослойная проверхностная ткань корня. Основной функцией эпиблемы является всасывание и поглощение из почвы воды с растворенными в ней минеральными веществами. В свою очередь через эпиблему могут выделяться биологически активные вещества, например кислоты. Клетки эпиблемы тонкостенные, лишены кутикулы, с вязкой цитоплазмой и содержат большое число митохондрий. Поглощающая поверхность эпиблемы может увеличиваться за счет образования корневых волосков. Они представляют собой выросты клетки длиной 1-3 мм. Эпидерма (кожица) – первичная покровная ткань образуется из наружного слоя клеток конуса нарастания побега на листьях, стеблях, цветках, плодах и семенах. Ткань защищает внутренние ткани от высыхания и повреждений. В состав эпидермы входят разные по морфологии клетки: основные клетки эпидермы, замыкающие и побочные клетки устьиц, трихомы. Основные клетки эпидермы плотно сомкнуты, межклетников нет, боковые стенки клеток извилистые. Клеточные стенки могут быть пропитаны кремнеземом (хвощи, осоки, злаки) или содержать слизи (семена льна). С наружной стороны клетки ткани покрыты сплошным слоем кутикулы. В ее состав может входить воск, образуя сплошной налет, состоящий из чешуек, палочек и других структур. Хорошо заметен сизый налет на листьях капусты, плодах сливы и винограда. Устьица являются специализированными структурами, осуществляющими газообмен и транспирацию. Устьице состоит из двух замыкающих клеток, между ними находится устьичная щель. Под ней расположена дыхательная подустьичная полость. К замыкающим клеткам могут примыкать побочные клетки, которые совместно представляют собой устьичный аппарат (рис.6). Стенки замыкающих клеток содержат много хлоропластов и митохондрий и неравномерно утолщены: брюшные (обращенные к щели) толще спинных (примыкающих к эпидерме). При повышении тургора тонкие стенки растягиваются, увлекая за собой толстые, и устьичная щель увеличивается. Роль хлоропластов заключается в том, что в процессе фотосинтеза повышается содержание сахаров и осмотическое давление. В результате в клетку засасывается большое количество воды, увеличивается тургор и размер вакуоли устьице открывается. В темноте устьичная щель закрыта за счет недостатка воды. В условиях достаточного увлажнения замыкающие клетки располагаются на одном уровне с основными клетками эпидермы, при избыточном – приподняты, при недостаточном – залегают глубже.
Рис. 6 Эпидерма листа гороха (А) и пшеницы (Б): 1- устьице закрытое; 2- устьице открытое (а- в плане, б- в разрезе); 3- замыкающие клетки; 4- устьичная щель; 5- побочные клетки; 6- подустьичная полость
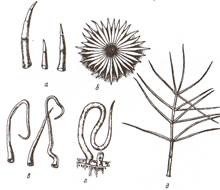
Трихомы - различные по форме, строению и функциям выросты эпидермы. Они имеют форму волосков (кроющие и железистые, одноклеточные, многоклеточные неразветвленные и разветвленные, звездчатые) (рис 7).
Рис. 7 Эпидермальные трихомы: а - простые многоклеточные на листе картофеля; б - звездчатые на листе лоха; в - простые одноклеточные на листе яблони; г - то же на семени хлопчатника; д - ветвистые многоклеточные на листе коровяка
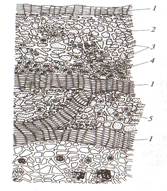
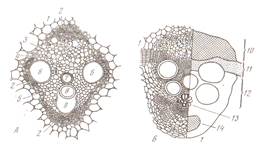
Волоски долго могут оставаться живыми, но при отмирании протопласта они заполняются воздухом и защищают растения от излишнего испарения и перепадов температуры. Жесткие, колючие, жгучие волоски защищают растения от насекомых, способствую опылению и распространению плодов. На эпидерме могут образовываться эмергенцы, их образуют ткани расположенные более глубоко (крапива, роза, ежевика). Эпидерма функционирует, как правило, один вегетационный период, обычно к осени ее заменяет пробка. Пробка (феллема) – вторичная покровная ткань развивается из клеток пробкового камбия, феллогена. Феллоген (образовательная ткань) – слой клеток боковой вторичной меристемы. Он возникает из основной паренхимы, лежащей под эпидермой (бузина) или более глубоко (смородина, вишня, малина), а иногда в самой эпидерме (груша, ива). При делении клеток феллогена кнаружи откладываются клетки феллемы, внутрь – феллодермы (основная ткань, хлорофиллоносная паренхима). Феллема, феллоген и феллодерма – это единый покровный комплекс – перидерма (рис 8).
Рис. 8 Перидерма стебля бузины: А- чечевичка; Б- участок перидермы; 1- выполняющая ткань; 2- остатки эпидермы;3- пробка (феллема); 4- пробковый камбий (феллоген); 5- феллодерма Перидерма образуется на ветвях, стволах, корнях и почечных чешуях (у древесных); на некоторых плодах; на корнях, гипокотиле, иногда на корневищах и клубнях (у трав). Клетки пробки соединены очень плотно, без межклетников. Сначала клеточные стенки тонкие, а затем утолщаются и состоят из чередующихся слоев суберина и воска, не пропуская воду и воздух. По мере того как эпидерму сменяет перидерма, зеленый цвет побегов приобретает бурый цвет. Газообмен и транспирация в органах покрытых перидермой, происходят через чечевички. Чечевички – это участок перидермы с рыхло расположенными клетками пробки с поверхности выглядит как бугорок. С наступлением холодов феллоген откладывает под выполняющей тканью замыкающий слой из плотно соединенных клеток, препятствующий испарению. Весной этот слой разрывается за счет вновь формирующихся клеток. По мере утолщения ветвей чечевички растягиваются. У большинства древесных растений по мере утолщения перидерма заменяется коркой. Она состоит из нескольких слоев пробки и заключенных между ними отмерших тканей (рис.9). Если образование перидерм происходит не по всей окружности ствола, а отдельными полудугами такая корка называется чешуйчатой (большинство растений).
Рис. 9 Схема строения корки: 1- пробка; 2- паренхима коры; 3- склереиды; 4- лубяные волокна; 5- клетки с друзами кристаллов Кольцевая корка формируется по всей окружности ствола, периодически сбрасывается на всем протяжении стебля, снижая его массу (виноград, лимонник, жимолость). Корка предохраняет стволы от механических повреждений, погрызов животными, пожаров, перепадов температур. 2.3 ОСНОВНЫЕ ТКАНИ
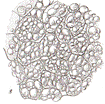
Основные ткани составляют большую часть тела растения. Клетки ткани живые паренхимные, тонкостенные с простыми порами. В зависимости от выполняемой функции различают основную (типичную), ассимиляционную, запасающую и воздухоносную паренхиму. Основная паренхима не имеет строго определенных функций, располагается внутри тела растения. Она заполняет сердцевину стебля, внутренние слои коры стебля и корня. Клетки ткани образуют вертикальные и горизонтальные тяжи, по которым в радиальном направлении перемещаются вещества. Из основной паренхимы могут возникнуть вторичные меристемы. Ассимиляционная паренхима (хлоренхима) осуществляет процесс фотосинтеза. Ткань обычно располагается под эпидермой, хорошо развита в листьях, меньше в молодых стеблях. Клетки тонкостенные с межклетниками имеют большое число хлоропластов. Запасающая паренхима служит для отложения избытка питательных веществ. Клетки ткани живые тонкостенные. Особенности строения клеток зависят от характера запасных веществ. Если это крахмал, клетки содержат много лейкопластов; если сахара или инулин, то крупные вакуоли; если белок, то много мелких вакуолей, образуют алейроновые зерна; если гемицеллюлоза, то толстые клеточные стенки. Запасающие ткани широко распространены и развиваются в разных органах (клубни картофеля, корнеплоды свеклы, моркови, луковицы лука, зерновки злаков, семена подсолнечника, клещевины) (рис. 10). У растений засушливых мест (агавы, алоэ, кактусы) в клетках запасающей паренхимы накапливается вода.
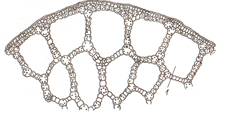
Рис. 10 Запасающая паренхима: А- семядоли гороха, Б- эндосперм пшеницы: 1- алейроновые зерна; 2- крахмальные зерна; В- клубень картофеля Воздухоносная паренхима (аэренхима) выполняет вентиляционные, дыхательные функции, обеспечивая ткани кислородом. Ткань хорошо развита у растений погруженных в воду (кувшинка, белокрыльник, рдест и др.) (рис. 11).
Рис. 11 Аэренхима стебля рдеста: 1- межклетник
2.4 МЕХАНИЧЕСКИЕ ТКАНИ
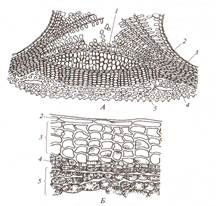
Механические ткани обеспечивают прочность растения и играют роль скелета. Клетки имеют утолщенные стенки. Различают колленхиму, склеренхиму и склереиды. Колленхима развивается в стеблях и черешках листьев двудольных растений под эпидермой или несколько глубже. Клетки ткани вытянутые, живые, часто содержат хлоропласты. Клеточные стенки неравномерно утолщены. В утолщениях чередуются слои целлюлозы и сильно обводненные слои, богатые пектином и гемицеллюлозой. Функции опорной ткани колленхима может выполнять только в состоянии тургора. В зависимости от характера утолщения стенок и их соединения различают уголковую, пластинчатую и рыхлую колленхиму (рис 12). Уголковая колленхима имеет стенки, утолщенные в углах клеток. Часто ткань встречается под эпидермой над главной жилкой листьев, по ребрам травянистых стеблей. Хорошо развита уголковая колленхима в стеблях тыквы, георгины, черешке листа свеклы. Пластинчатая колленхима имеет утолщенные тангентальные (параллельные поверхности органа) стенки клеток. Часто образует в стебле сплошное кольцо (подсолнечник, клевер, баклажан).
Рис. 12 Колленхима: а- уголковая колленхима в черешке листа свеклы; б- пластинчатая колленхима в стебле подсолнечника: 1- кутикула; 2- эпидерма; 3- утолщенные стенки клеток колленхимы Склеренхима. Первичная склеренхима развита во всех вегетативных органах однодольных, реже двудольных растений; вторичная – у двудольных. Клетки склеренхимы – волокна, имеющие равномерно утолщенные, как правило, одревесневшие стенки. Протопласт отмирает рано, опорную функцию выполняют мертвые клетки (рис. 13). Склеренхимные волокна – сильно вытянутые прозенхимные клетки, обеспечивают прочность органов растений на растяжение, сжатие и изгиб. Концы клеток часто заострены (лен), могут быть ветвистыми (конопля), тупыми (крапива).
Рис. 13 Склеренхима и склереиды: а- волокна склеренхимы; б- каменистые клетки из плода груши; в- склеренхима на поперечных срезах стебля льна Склеренхимные волокна располагаются в растении в виде отдельных клеток (элементарное волокно) или, соединяясь друг с другом по длине, образуют пучок (техническое волокно). Склереиды – чаще всего имеют паренхимную форму и располагаются в растении плотными группами или одиночно. Окончательно сформировавшиеся ткани это мертвые клетки с толстыми одревесневшими стенками, пронизаны поровыми каналами, нередко ветвистыми. Поры простые. Склереиды имеют первичное происхождение. К ним относят каменистые и ветвистые клетки (рис.13). Каменистые клетки – округлые, обычно встречаются группами. Из них состоят косточки вишни, сливы, персика и скорлупа ореха. Встречаются также в сочных плодах груши, айвы, рябины, в корнях хрена. Ветвистые клетки имеют причудливую форму, выполняют роль опоры в листьях чая, камелии, маслины, в стеблях водных растений.
2.5 ПРОВОДЯЩИЕ ТКАНИ И КОМПЛЕКСЫ
Они образуют в теле растения не прерывную разветвленную систему, соединяющую все органы. В растениях все вещества транспортируются в двух направлениях: от корней к листьям поднимается восходящий ток водных растворов минеральных солей. В этом участвуют трахеальные элементы (трахеиды и сосуды) (рис. 14);
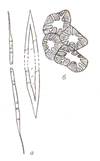
Рис. 14 Трахеальные элементы: А- типы сосудов: кольчатый (1), спиральный (2…4), сетчатый (5); б- закупорка сосуда (1) выростами клеток древесинной паренхимы – тилами (2,3)
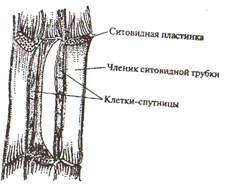
от листьев к корням идет нисходящий ток органических веществ, который обслуживают ситовидные элементы (ситовидные клетки и ситовидные трубки) (рис. 15). Трахеида представляет собой удлиненную клетку с острыми или округлыми концами и одревесневшими стенками. Поры только окаймленные. Живое содержимое трахеи постепенно отмирает. Большая часть окаймленных пор находится на концах клеток, где раствор переходит из одной поры в другую. Трахеиды встречаются у всех высших растений, а у большинства хвощей, плаунов, папоротников и голосеменных являются единственной проводящей тканью. Трахея (сосуд) состоит из многих клеток-члеников. Они располагаются друг над другом и образуют полые трубки. Поперечные стенки соприкасающихся клеток местами растворяются и возникают отверстия (перфорации), по которым и перемещается влага из одного членика сосуда в другой. Растворы передвигаются и в поперечном направлении через неутолщенные участки боковых стенок или поры в них. В зависимости от характера утолщения боковых стенок различают: кольчатые, спиральные, сетчатые, лестничные и точечно-поровые трахеиды и сосуды (рис.14). Сосуды функционируют ограниченное время. Они прекращают свою деятельность, если их закупоривают тилы (выросты соседних клеток, проникающие в полость сосуда через поры. В дальнейшем сосуды выполняют механические функции. В обеспечении восходящего тока участвует и ксилема (древесина). В ксилеме имеются живые клетки древесинной паренхимы и древесинные волокна (либриформ). По паренхиме, окружающей трахеальные элементы и контактирующей с ними, происходит ближний радиальный транспорт. В этих клетках накапливаются запасные вещества. Весной они превращаются в растворы сахаров и поступают в сосуды. Основная функция сосудов – проведение минеральных веществ и воды. Нисходящий ток обеспечивают ситовидные клетки и ситовидные трубки с клетками спутницами. Они сохраняют живой протопласт, клеточные стенки целлюлозные. Ситовидная клетка сильно вытянута в длину, концы заостренные. В зрелых клетках сохраняется ядро. Такие клетки характерны для высших споровых и голосеменных растений. Ситовидные трубки состоят из удлиненных живых клеток– члеников, расположенных друг над другом. Поперечные стенки члеников ситовидных трубок представляют собой ситовидные пластинки. Они пронизаны множеством мелких отверстий (перфораций), образующих канальцы. Через эти отверстия из клетки в клетку идут тяжи цитоплазмы. Ядра в этих клетках отсутствуют. Рядом с члеником ситовидной трубки располагаются одна или несколько клеток-спутниц с густой цитоплазмой, ядром, митохондриями и рибосомами. Ситовидные трубки с клетками – спутницами характерны для покрытосеменных (рис. 15). Осенью ситовидные пластинки затягиваются каллезой и трубка перестает функционировать. У некоторых древесных растений ситовидные трубки действуют в течение трех-четырех лет (липа), у других - два года (виноград, пихта), у большинства растений - в течение одного вегетационного периода.
Рис. 15 Элементы флоэмы на продольном срезе
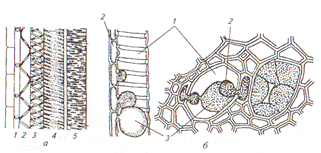
Ситовидные элементы - основные компоненты проводящего комплекса, который получил название флоэма (луб). Живые тонкостенные клетки лубяной паренхимы участвуют в ближнем транспорте ассимилятов, в них откладываются запасные вещества. Лубяные волокна играют опорную роль. Ксилема и флоэма образуются в результате работы специальных меристем – прокамбия (первичные) и камбия (вторичные). Проводящие пучки образуются за счет располагающихся рядом друг с другом тяжей ксилемы и флоэмы (рис. 16).
Рис. 16 Проводящие пучки (схема поперечных разрезов): А- коллатеральный, Б- биколлатеральный, В, Г, - концентрические (В- с наружной ксилемой, Г- с наружной флоэмой), Д- радиальный; 1- флоэма, 2- ксилема
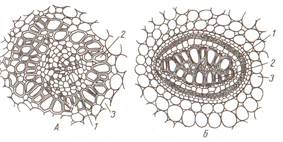
Развитие проводящих пучков начинается под конусом нарастания из клеток прокамбия. Часть клеток, обращенные к периферии органа, превращается в элементы первичной флоэмы, а остальные - в элементы первичной ксилемы. Между ними не остается меристематических клеток, которые могли бы дать новые проводящие элементы. Такие пучки закончили свой рост, их называют закрытыми (рис. 17 А ). Они свойственны однодольным и папоротникообразным растениям, но встречаются и у двудольных.
Рис. 17 Сосудисто-волокнистые пучки в поперечном разрезе: А- коллатеральный закрытый стебля кукурузы, Б- биколлатеральный открытый стебля тыквы (слева- детальный, справа – схематичный); 1- тонкостенная основная паренхима стебля, 2- склеренхима, 3- сопровождающая клетка, 4- ситовидная трубка, 5- древесинная паренхима, 6- сетчатые сосуды, 7- кольчато-спиральный сосуд, 8- кольчатый сосуд, 9- полость, 10- наружная флоэма, 11- камбий, 12- вторичная ксилема, 13- первичная ксилема, 14- внутренняя флоэма
В большинстве случаев после образования первичных флоэмы и ксилемы между ними остаются меристематические клетки. Они начинают делиться и образуется камбий. Благодаря делению клеток камбия к периферии нарастают элементы вторичной флоэмы, а к центру - вторичной ксилемы. Такой пучок открыт для дальнейшего роста и его называют открытым (рис. 17 Б). По взаиморасположению ксилемы и флоэмы различают пучки нескольких типов: - Коллатеральные (флоэма лежит кнаружи от ксилемы) пучки могут быть открытыми и закрытыми (встречаются наиболее часто). - Биколлатеральные пучки имеются у семейства Пасленовые, Тыквенные, Вьюнковые. В таких пучках флоэма расположена с обеих сторон ксилемы, пучки открытые (подсолнечник). Наружная флоэма – первичная и вторичная – отделена от ксилемы камбием, внутренняя флоэма - только первичная. - Концентрические пучки, в которых или ксилема окружает флоэму, или флоэма - ксилему, всегда закрытые (рис. 18).
Рис. 18 Концентрические проводящие пучки: А- с наружной ксилемой в корневище ландыша, Б- с наружной флоэмой в корневище папоротника- орляка; 1- флоэма, 2- ксилема, 3- основная паренхима стебля.
В молодых корнях у всех растений развиваются радиальные закрытые пучки. В них ксилема и флоэма расположены по радиусам (рис 16).
Дата добавления: 2016-06-05 | Просмотры: 1459 | Нарушение авторских прав |
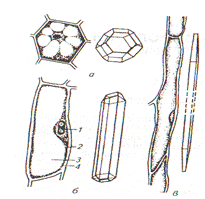

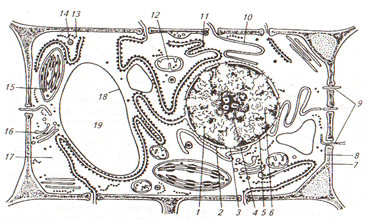
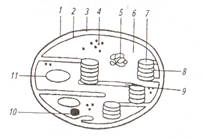
 Рис. 4 Схема строения митохондрий:
Рис. 4 Схема строения митохондрий: