|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Процессы торможения в центральной нервной системе.В центральной нервной системе постоянно функционируют два основных, взаимосвязанных процесса - возбуждение и торможение. Торможение - это активный биологический процесс, направленный на ослабление, прекращение или предотвращение возникновения процесса возбуждения. Явление центрального торможения, т. е. торможения в ЦНС, было открыто И. М. Сеченовым в 1862 г. в опыте, получившим название "опыт сеченовского торможения". Суть опыта: у лягушки на срез зрительных бугров накладывали кристаллик поваренной соли, что приводило к увеличению времени двигательных рефлексов, т. е. к их торможению. Время рефлекса - это время от начала раздражения до начала ответной реакции. Торможение в ЦНС выполняет две основные функции. Во-первых, оно координирует функции, т. е. оно направляет возбуждение по определенным путям к определенным нервным центрам, при этом выключая те пути и нейроны, активность которых в данный момент не нужна для получения конкретного приспособительного результата. Важность этой функции процесса торможения для функционирования организма можно наблюдать в эксперименте с введением животному стрихнина. Стрихнин блокирует тормозные синапсы в ЦНС (в основном глицинергические) и тем самым устраняет основу для формирования процесса торможения. В этих условиях раздражение животного вызывает некоординированную реакцию, в основе которой лежит диффузная (генерализованная) иррадиация возбуждения. При этом приспособителъная деятельность становится невозможной. Во-вторых, торможение выполняет охранительную или защитную функцию, пред охраняя нервные клетки от перевозбуждения и истощения при действии сверхсильных и длительных раздражителей. Теории торможения. Н. Е. Введенским (1886) было показано, что очень частые раздражения нерва нервно-мышечного препарата вызывают сокращения мышцы в виде гладкого тетануса, амплитуда которого мала. Н. Е. Введенский полагал, что в нервно-мышечном препарате при частом раздражении возникает процесс пессимального торможения, т. е. торможение является как бы следствием перевозбуждения. Сейчас установлено, что его механизм заключается в длительной, застойной деполяризации мембраны, вызванной избытком медиатора (ацетилхолина), выделяющегося при частой стимуляции нерва. Мембрана полностью теряет возбудимость из-за инактивации натриевых каналов и не в состоянии ответить на приход новых возбуждений выделением новых порций медиатора. Таким образом, возбуждение переходит в противоположный процесс - торможение. Следовательно, возбуждение и торможение являются как бы одним и тем же процессом, возникают в одних и тех же структурах, с участием одного и того. же медиатора. Данная теория торможения называется унитарно-химической или монистической. Медиаторы на постсинаптической мембране могут вызывать не только деполяризацию (ВПСП), но и гиперполяризацию (ТПСП). Эти медиаторы увеличивают проницаемость субсинаптической мембраны для ионов калия или хлора, в результате чего постсинаптическая мембрана гиперполяризуется и возникает ТПСП. Данная теория торможения получила название бинарно-химической, согласно которой торможение и возбуждение развиваются по разным механизмам, с участием тормозных и возбуждающих медиаторов соответственно. Классификация центрального торможения. Торможение в ЦНС можно классифицировать по различным признакам: • по электрическому состоянию мембраны - деполяризационное и гиперполяризационное; • по отношению к синапсу - пресинаптическое и постсинаптическое; • по нейрональной организации - поступательное, латеральное (боковое), возвратное, реципрокное. Постсинаптическое торможение развивается в условиях, когда медиатор, выделяемый нервным окончанием, изменяет свойства постсинаптической мембраны таким образом, что способность нервной клетки генерировать процессы возбуждения подавляется. Постсинаптическое торможение может быть деполяризационным, если в его основе лежит процесс длительной деполяризации, и гиперполяризационным, если - гиперполяризации. Пресинаптическое торможение обусловлено наличием вставочных тормозных нейронов, которые формируют аксо-аксональные синапсы на афферентных терминалях, являющихся пресинаптическими по отношению, например, к мотонейрону. В любом случае активации тормозного интернейрона, он вызывает деполяризацию мембраны афферентных терминалей, ухудшающей условия проведения по ним ПД, что таким образом уменьшает количество выделяемого ими медиатора, и, следовательно, эффективность синаптической передачи возбуждения к мотонейрону, что уменьшает его активность (рис. 14). Медиатором в таких аксо-аксональных синапсах является, по-видимому, ГАМК, которая вызывает повышение проницаемости мембраны для ионов хлора, которые выходят из терминали и частично, но длительно ее деполяризуют.
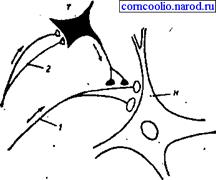
Рис. 14. Пресинаптическое торможение (схема): Н - нейрон, возбуждаемый афферентными импульсами, приходящими по волокну 1; Т - нейрон, образующий тормозные синапсы на пресинаптических разветвлениях волокна 1; 2 - афферентные волокна, вызывающие активность тормозного нейрона Т. Поступательное торможение обусловлено включением тормозных нейронов на пути следования возбуждения (рис. 15).
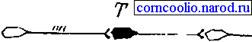
Рис. 15. Схема поступательного торможения. Т - тормозньй нейрон Возвратное торможение осуществляется вставочными тормозными нейронами (клетками Реншоу). Импульсы от мотонейронов, через отходящие от его аксона коллатерали, активируют клетку Реншоу, которая в свою очередь вызывает торможение разрядов данного мотонейрона (рис. 16). Это торможение реализуется за счет тормозных синапсов, образованных клеткой Реншоу на теле активирующего ее мотонейрона. Таким образом, из двух нейронов формируется контур с отрицательной обратной связью, которая дает возможность стабилизировать частоту разряда мотонейрона и подавлять избыточную его активность.
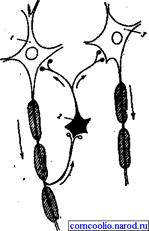
Рис. 16. Схема возвратного торможения. Коллатерали аксона мотонейрона (1) контактируют с телом клетки Реншоу (2), короткий аксон которой, разветвляясь, образует тормозные синапсы на мотонейронах 1 и 3. Латеральное (боковое) торможение. Вставочные клетки формируют тормозные синапсы на соседних нейронах, блокируя боковые пути распространения возбуждения (рис. 17). В таких случаях возбуждение направляется только по строго определенному пути.
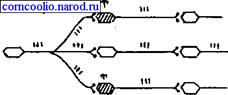
Рис. 17. Схема латерального (бокового) торможения. Т - тормозный нейрон. Именно латеральное торможение обеспечивает, в основном, системную (направленную) иррадиацию возбуждения в ЦНС. Реципрокное торможение. Примером реципрокного торможения является торможение центров мышц-антагонистов. Суть этого вида торможения заключается в том, что возбуждение проприорецепторов мышц-сгибателей одновременно активирует мотонейроны данных мышц и вставочные тормозные нейроны (рис. 18). Возбуждение вставочных нейронов приводит к постсинаптическому торможению мотонейронов мышц-разгибателей.
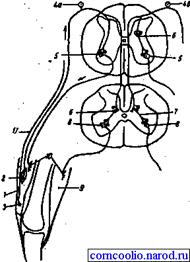
Рис. 18. Схема реципрокного торможения. 1 - четырехглавая мышца бедра; 2 - мышечное веретено; 3 - сухожильный рецептор Гольджи; 4 - рецепторные клетки спиномозгового ганглия; 4а - нервная клетка, воспринимающая импульсы от мышечного веретена; 4б - нервная клетка, воспринимающая имульсы от рецептора Гольджи; 5 - мотонейроны, иннервирующие мышцы-разгибатели; 6 - тормозный промежуточный нейрон; 7 - возбуждающий промежуточный нейрон; 8 - мотонейроны, иннервирующие мышцы-сгибатели; 9 - мышца-сгибатель; 10 - моторные нервные окончания в мышцах; 11 - нервное волокно от сухожильного рецептора Гольджи. Дата добавления: 2015-12-15 | Просмотры: 992 | Нарушение авторских прав |