|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ВИСЦЕРАЛЬНЫЕ ХЕМОРЕЦЕПТОРЫ
Благодаря электрофизиологическим исследованиям афферентных рецепторов внутренних органов были обнаружены рецепторы, реагирующие на изменение химического состава окружающей их среды. Среди висцеральных хеморецепторов можно выделить следующие рецепторы: изменяющие свою электрическую активность на сдвиг рН среды; реагирующие на изменение концентрации глюкозы — глюкорецепторы; рецепторы некоторых аминокислот; реагирующие на содержание кислорода или диоксида углерода: в крови — это так называемые каротидные и аортальные тельца и во вдыхаемом воздухе — нейроэпителиальные тельца; сигнализирующие об изменении осмотического давления внутренней среды. За исключением каротидных, аортальных и нейро-эпителиальных телец, о структуре и механизмах рецепции в висцеральных хеморецепторах известно сравнительно немного, хотя расположены они в различных участках тела. Щелочно- и кислоточувствительные рецепторы. Обнаружены в слизистой оболочке желудочно-кишечного тракта животных. Они, по-видимому, являются не просто детекторами рН среды, поскольку их реакция зависит еще и от типа кислоты или щелочи. Кислотные рецепторы увеличивают частоту импульсации с повышением концентрации ионов водорода в среде (т. е. снижением рН), а щелочные рецепторы повышают частоту афферентных потенциалов действия с ростом концентрации ОН~-группы. 44 — 3389 689
Глюкорецепторы. Обнаружены в желудочно-кишечном тракте и печени животных. Увеличение концентрации глюкозы вызывает повышение частоты афферентной импульсации в нервных волокнах, образующих эти рецепторы в желудке и тонком кишечнике (рис. 15.20, А). Однако известно, что изменение концентрации глюкозы влияет на осмотическое давление раствора, т. е. глюкорецепторы могут быть просто осморецепторами. При повышении осмотичности раствора за счет других веществ, например таких, как хлорид натрия или калия, не обнаружили какой-либо реакции со стороны глюкорецепторов (рис. 15.20, Б, В). Возбуждение глю-корецепторов адекватными стимулами приводило к быстрому повышению уровня инсулина в крови у животных. Надо отметить, что некоторые глюкорецепторы возбуждают и другие углеводы, причем выражена специфичность реакции различных рецепторов на тип углевода. Реакция глюкорецепторов печени зависит от концентрации глюкозы в крови: при низких концентрациях глюкозы в афферентных волокнах регистрируются потенциалы действия с определенной частотой, а ее повышение вызывает снижение частоты импульсации. Аминокислотные висцеральные рецепто-р ы. Рецепторные структуры, реагирующие изменением импульсной активности на аминокислоты, были обнаружены в кишечнике плотоядных животных. Эти рецепторы не активировались другими химическими веществами, а также механическим раздражением. Короткий латентный период ответов на глюкозу и аминокислоты позволяет считать, что эти рецепторы также находятся в слизистой оболочке внутренних органов животных. Осморецепторы. Во внутренних органах были обнаружены рецепторы, реагирующие на изменение осмотического давления окружающей их жидкости. Реакция данных рецепторов не зависела от природы вещества, с помощью которого было вызвано повышение осмотичности раствора.
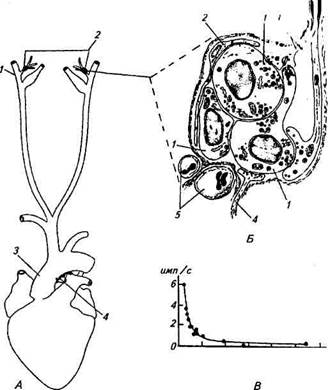
Рис. 15.20. Гистограммы афферентной импульсной активности, иллюстрирующие специфичность глюкорецепторов кишечника: А. Реакция глюкорецепторов на раствор глюкозы. Б, В. Реакция на растворы NaCl и КС1 (осмотичность как раствора глюкозы); стрелками показано начало действия растворов. Г. Реакция на сильное растяжение стенки кишки; горизонтальной линией отмечено время механического растяжения Рис. 15.21. Схема расположения каротидного и аортального телец (Л), участок каротидного тельца (Б) и зависимость частоты афферентных потенциалов действия каротидного тельца от содержания кислорода в крови (В): А. 1 — сонная артерия; 2 — место расположения каротидного тельца; 3 — аорта; 4— место расположения аортальных телец. Б. 1 — хеморецептор-ная клетка; 2— опорная клетка; 3 — афферентное нервное волокно; 4— эфферентное нервное волокно; 5— капилляры. В. По оси ординат — частота потенциалов действия в афферентном нервном волокне каротидного тельца, по оси абсцисс — напряжение кислорода в крови Каротидные и аортальные тельца. Структура и функция этих хеморецепторов наиболее изучены среди висцеральных рецепторов. Они являются хеморецепторами артериального русла и сосредоточены в двух зонах: при ветвлении общей сонной артерии между наружной и внутренней ее ветвями {каротидные тельца) и дуги аорты {аортальные тельца) (рис. 15.21, А). В основном эти рецепторы чувствительны к со-
44*
Каротидное тельце образовано соединительнотканной капсулой, внутри которой располагаются клетки двух типов: хемо-рецепторы (гломерулярные клетки) и опорные (рис. 15.21, Б). Хеморецепторные клетки округлые, диаметр их Ю...15мкм; часто они имеют цитоплазматические отростки различной длины. У опорных клеток также есть пальцевидные отростки, которые частично оплетают рецепторные клетки. Внутри капсулы рецепторные клетки с прилегающими к ним опорными клетками располагаются островками — кластерами, отделенными друг от друга соединительнотканными перегородками. Соотношение между числом рецепторных и опорных клеток внутри кластера примерно 5:1. Хеморецепторные клетки образуют плотные контакты друг с другом. Кровоснабжение каротидных телец осуществляется тонкими артериями, отходящими от основной артерии. Чрезвычайно густая сеть капилляров проходит через стенки капсулы каждого тельца в кластеры; внутри телец капилляры переходят в венулы. Через соединительнотканную стенку телец проникают также афферентные и эфферентные волокна, которые под капсулой теряют миелиновую оболочку, пересекают стенку кластера и образуют разнообразные по форме афферентные и эфферентные синаптические окончания с хеморецепторными клетками. Таким образом, рецептором каро-тидного тельца является специальная клетка, синаптически связанная с первым афферентным нейроном. В афферентных нервных волокнах каротидного тельца при нормальном напряжении кислорода в крови животного (80...90 мм рт. ст.) регистрируется спонтанная импульсная активность. Это указывает на то, что хеморецепторная клетка изначально деполяризована и из ее пресинаптической области происходит выброс медиатора. Микроэлектродные измерения подтвердили данное предположение. В большинстве случаев мембранный потенциал рецепторных клеток сдвинут на 30...40 мВ в сторону деполяризации от равновесного калиевого потенциала, что в основном определяет ее высокую чувствительность к химическому стимулу. При ступенчатообразном уменьшении напряжения кислорода в крови через 1 с начинает увеличиваться частота афферентных потенциалов действия, которая в течение 3 с достигает максимальной величины для данного значения напряжения кислорода в крови. В дальнейшем частота импульсации остается на неизменном уровне, т. е. адаптация у каротидных рецепторов отсутствует. Спад напряжения кислорода до исходного значения сопровождается снижением в течение 2...3 с частоты импульсации до первоначального уровня. Зависимость между уменьшением напряжения кислорода в крови и средней частотой афферентных потенциалов действия имеет вид гиперболы (рис. 15.21,5). Физиологическое пороговое значение напряжения кислорода находится на уровне 75 мм рт. ст. Максимальная частота потенциалов действия в нервных волокнах каротидного тельца наблюдается при снижении напряжения кислорода до 10 мм рт.ст. При увеличении напряжения диоксида углерода в крови до 100 мм рт. ст. или уменьшении рН крови до 6,9 происходит также усиление частоты афферентных потенциалов действия в нервных волокнах каротидного тельца. Зависимость между частотой и напряжением диоксида углерода и рН в данных пределах близка к линейной. Однако дальнейшее увеличение этих значений сопровождается лишь незначительным приростом частоты афферентных потенциалов действия. В экспериментах, проведенных на изолированных хеморецеп-торных клетках каротидного синуса кроликов, было выявлено в мембране клеток несколько типов ионных каналов, и в том числе каналов, чувствительных к содержанию кислорода в крови. В частности, было показано, что при снижении напряжения кислорода блокируется одна из компонент ионного тока, переносимого ионами калия. Полагают, что в мембране хеморецепторной клетки имеются калиевые каналы с рецепторным белком, активируемым кислородом. При нормальном содержании кислорода в крови белок активирован и калиевые каналы открыты. Снижение концентрации кислорода в крови приводит к инактивации белка, закрытию калиевых каналов и к сдвигу мембранного потенциала в сторону деполяризации. Деполяризация мембраны вызывает дополнительный выход медиатора из клетки и возбуждение афферентных синапсов с последующим увеличением частоты потенциалов действия в нервном волокне. Вопрос о механизмах рецепции напряжения диоксида углерода в крови и рН остается менее изученным. Нервные волокна от каротидных телец формируют так называемый синусный нерв, представляющий собой ветвь языкоглоточ-ного нерва. Тела нервных клеток, образующих волокна каротидных телец, находятся в шейном ганглии. Эффект стимуляции каротидных рецепторов проявляется в увеличении дыхательного объема и удлинении фазы вдоха. Импульсация от каротидных рецепторов вызывает возбуждение нейронов, ответственных за фазу вдоха, и задерживает его выключение. Нейроэпителиальные тельца воздухоносных путей легких. Аналогом каротидных и аортальных телец в воздухоносных путях легких являются нейроэпителиальные тельца. Они обнаружены у различных видов дышащих воздухом позвоночных животных — от млекопитающих до амфибий.
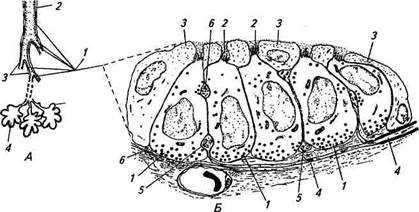
Рис. 15.22. Схема расположения неироэпителиальных телец (Л) в воздухоносных путях и строения нейроэпителиального тельца (Б): А. 1 — легкие; 2—бронхи; 3 — бронхиола; 4— альвеола. Б. 1 — хеморецепторные клетки; 2—микровиллы; 3 — Клара-клетки; 4— нервное волокно; 5 — афферентное нервное окончание; б— эфферентное нервное окончание да в окружающей среде. Недостаток кислорода вызывает закрытие калиевых ионных каналов хемочувствительных клеток и соответственно деполяризацию их мембран. Деполяризация, в свою очередь, усиливает выброс медиатора из клеток с последующим возбуждением афферентных синапсов и генерации в отходящих от них нервных волокнах серии потенциалов действия. Надо отметить, что помимо участия в рефлекторной регуляции функционирования легких через центральные нервные структуры нейроэпителиальные тельца могут регулировать работу легочных структур. Оказалось, что хеморецепторные клетки, образующие нейроэпителиальные тельца, одновременно являются нейро-эндокринными клетками. Они могут осуществлять синтез и секрецию таких биологически активных веществ, как серотонин, кальцитонин-либерин, бомбезин, гастрин-либерин и др. Выделяясь из клеток под воздействием различных факторов, находящихся во вдыхаемом воздухе (в том числе и под влиянием гипоксии), в близлежащие капилляры или межклеточное пространство, они могут вызывать расслабление или сжатие кровеносных сосудов и бронхов. Кроме того, ряд веществ, секретирующихся в клетках неироэпителиальных телец, ускоряют в постнатальном периоде формирование легких.
Нейроэпителиальные тельца детектируют содержание кислорода в полостях бронхов и бронхиолей и располагаются в эпителиальной выстилке слизистой оболочки их стенок (рис. 15.22, А). Нейроэпителиальное тельце (см. рис. 15.22, Б) образовано группой плотно прилегающих друг к другу хеморецепторных клеток с явно выраженной поляризацией. На апикальном конце клеток расположены реснички — микровиллы. Как правило, верхняя часть клеток неироэпителиальных телец у различных животных покрыта слоем уплощенных Клара-клеток так, что микровиллы ре-цепторных клеток сообщаются с воздушной полостью через заполненные слизью узкие поры между Клара-клетками. В базаль-ной части хеморецепторных клеток содержится большое количество гранулированных везикул. Иннервация неироэпителиальных телец осуществляется нервными волокнами, входящими в состав блуждающего нерва. Тела нервных клеток, образующих эти волокна, находятся в узловатом ганглии. Нервные волокна формируют на клетках неироэпителиальных телец афферентные и эфферентные синапсы. Причем часто от афферентного аксона, образующего афферентный синапс, отходит колатераль, заканчивающаяся эфферентным синапсом. К базальным областям неироэпителиальных телец подходят капилляры. Так же как у хемочувствительных клеток каротидных телец, в мембране клеток неироэпителиальных телец были обнаружены калиевые ионные каналы, реагирующие на изменение концентрации кислоро- Дата добавления: 2015-12-15 | Просмотры: 898 | Нарушение авторских прав |


 держанию кислорода в омывающей их крови, но вместе с тем они обладают четко выраженной реакцией на содержание С02 и изменение рН в кровяном русле. Каротидные и аортальные тельца по своему строению и функции имеют много общего, однако у большинства животных главенствующее положение занимают каротидные тельца.
держанию кислорода в омывающей их крови, но вместе с тем они обладают четко выраженной реакцией на содержание С02 и изменение рН в кровяном русле. Каротидные и аортальные тельца по своему строению и функции имеют много общего, однако у большинства животных главенствующее положение занимают каротидные тельца.