|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
СЛУХОВОЙ АНАЛИЗАТОРСлуховой анализатор предназначен воспринимать механические колебания (звуковые волны) из внешней среды, создаваемые источниками, находящимися на удалении, иногда весьма значительном, от животного. Звукопередающей средой для наземных животных является воздух, а для водных животных — вода, в которой звук распространяется в 4 раза быстрее, чем в воздухе. Звуковые сигналы, воспринимаемые животными, подразделяют на звуки небиологического происхождения (зависящие от силы ветра, выпадения осадков, шума листвы деревьев и др.) и звуки биологического происхождения, связанные с сигналами акустического общения различных животных, а также шорохами и шумами, производимыми хищниками или их жертвами. Развитие слухового анализатора в процессе эволюции достигло наибольшего совершенства у млекопитающих. Периферическая часть слухового анализатора состоит из трех отделов: наружного, среднего и внутреннего уха (рис. 15.12). Наружное ухо включает в себя ушную раковину и наружный слуховой проход. По своей форме наружное ухо напоминает воронку, которая направляет звуковые волны в слуховой проход, т. е. обеспечивает улучшенное восприятие звуков. У многих животных рефлекторное управление подвижностью ушной раковины хорошо развито с помощью специальных мышц, что не требует поворота головы. Раз- |
1Рейснерова Сосудистая '.А Внутренние
Средняя \ лестница v-ygS (эндолимфа) ^3*5
Афферентные (перилимфа) нервные волокно Б Рис. 15.12. Ухо млекопитающих:
А. Общая схема строения (полукружные каналы не показаны); Б. Поперечный разрез через улитку; В. Схема кортиевого органа
меры и форма ушной раковины варьируют у животных разных видов в значительной степени: очень подвижны ушные раковины у кошек, лошадей, некоторых пород собак. Наружный слуховой проход представляет собой узкую, несколько искривленную трубку, по которой звуковые колебания проходят внутрь уха. Специальные железы в коже наружнего слухового прохода вырабатывают секрет —ушную серу, которая предохраняет ухо от загрязнения и препятствует высыханию барабанной перепонки, отделяющей наружное ухо от среднего. Барабанная перепонка крепится на внутреннем костном конце наруж- ного слухового прохода (см. рис. 15.12, Л), толщина г.. ,„ инмм. ■ 100...200 мкм. Назначение барабанной перепонки шмнпч.к пи и передаче без искажений частоты воздействующих на п<ч тумими колебаний. Необходимым условием для этого является uhvh i ни*, у нее резонансных явлений, т. е. она не усиливает звукош.к- ими-бания. Это возможно благодаря особенностям ее строения, а шк же расположению ее в слуховом проходе. Барабанная перепои ка — это соединительнотканная мембрана, образованная из радиальных и кольцевых волокон. Степень натяжения ее отдельных участков неодинакова; в звуковом проходе она расположена косо и середина ее втянута внутрь.
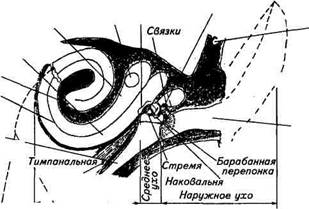
Среднее ухо — это трехкосточковая звукопередающая рычажная система, включающая в себя последовательно соединенные косточки: молоточек, наковальню и стремячко. Большая часть поверхности барабанной перепонки контактирует с ручкой молоточка, который сочленяется связками с наковальней. Наковальня соединена со стремечком, связанным с мембраной овального окошка, открывающегося в полость преддверия (внутреннее ухо), заполненной жидкостью. Расположение и механические свойства сруктур среднего уха обеспечивают оптимальную передачу звуковых колебаний из воздушной среды в жидкую среду внутреннего уха. В частности, эффективная часть барабанной перепонки (т. е. находящаяся в контакте с молоточком) примерно в 17 раз больше, чем площадь основания стремечка, кроме того, способ вращения косточек дополнительно увеличивает давление на овальное окно — отверстие, ведущее во внутреннее ухо. Благодаря всему этому давление на овальное окно по сравнению с барабанной перепонкой увеличивается примерно в 20 раз. Таким образом, рычажная система среднего уха уменьшает амплитуду перемещений барабанной перепонки, превращая их в незначительные по размаху, но в соответствующее число раз усиленных по давлению перемещений стремечка. Этот механизм увеличения давления является чрезвычайно важным приспособлением, обеспечивающим эффективную передачу акустической энергии из воздушной среды в жидкую. В системе среднего уха функционируют несколько механизмов, предохраняющих повреждение его структур от весьма сильных звуковых колебаний. Так, полость среднего уха не закрыта, а соединяется с полостью глотки особым каналом — евстахиевой трубой, служащей для уравнивания давления по обе стороны барабанной перепонки. Это имеет большое значение для предохранения барабанной перепонки при значительной разнице давления в барабанной полости и в окружающей среде. Такая разница может возникнуть при попадающей в ухо звуковой волны, например от раската сильного грома. Раскрытие евстахиевой трубы способствует выравниванию давления и предупреждает разрыв барабанной перепонки.
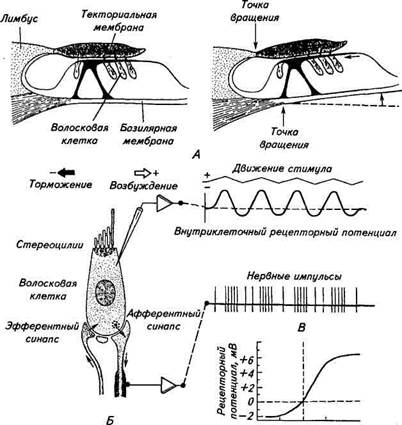
Другой охранительный механизм заключается в рефлекторном сокращении мышц среднего уха в ответ на чрезмерно сильные звуки. Связанная с молоточком тимпанальная мышца, сокращаясь, смещает барабанную перепонку внутрь, а одновременное сокращение мышцы стремечка ограничивает движение стремечка и мембраны овального окна, что в совокупности приводит к уменьшению давления, передаваемого рычажным механизмом. В костной перегородке, отделяющей среднее ухо от внутреннего, кроме овального окна имеется также круглое окно, или окно улитки. Оно затянуто тонкой соединительнотканной мембраной и может служить дополнительным приспособлением для передачи звуковых колебаний в случае повреждения барабанной перепонки и косточек. Внутреннее ухо расположено в каменистой части височной кости и состоит из костного и перепончатого лабиринтов (см. рис. 15.12, Б). В состав периферической части слухового анализатора входит лишь часть перепончатого лабиринта, который помещается в костном, как в футляре, и точно повторяет его форму. Звуковой лабиринт представляет собой суживающуюся к концу трубку, спирально закрученную наподобие раковины — улитки. Число витков улитки у различных млекопитающих бывает разное: у человека — 2,5 витка, у кошки — 3, у свиньи — 4. Костная улитка разделена вдоль костной спиральной стенкой, которая, однако, не доходит до противоположной стенки, а ее продолжает основная — базилярная мембрана. В начале улитки костная спиральная стенка имеет наибольшую ширину, а базилярная мембрана наименьшую (см. рис. 15.12, Б). По мере движения к вершине улитки ширина костной стенки уменьшается, а базилярной мембраны увеличивается. Комбинация костной перегородки с базилярной мембраной делит просвет улитки на два главных продольных канала: барабанная лестница и лестница преддверия. Они заполнены жидкостью, называемой перилимфой, и сообщаются между собой у верхушки улитки через небольшое отверстие — геликотрему, диаметром около 20мкм. Между этими каналами лежит третий канал — внутренняя лестница, отделенный от них базилярной и рейснеровой мембраной, заполненный жидкостью — эндолимфой. Перилимфа и эндолимфа у млекопитающих отличаются в значительной степени по ионному составу друг от друга, а также от плазмы крови. Так, концентрация ионов калия в эндолимфе в 30...35 раз превышает их концентрацию в плазме крови, а в пери-лимфе — примерно в 1,5...2,5 раза. И наоборот, концентрация натрия в эндолимфе примерно в такой же пропорции ниже, чем в плазме крови и перилимфе. Причина ионной асимметрии связана с функционированием электрогенных ионных насосов. Разница в концентрации заряженных частиц сопровождается возникновением разности по- тенциалов. В каналах улитки обнаружили положительные (относительно плазмы крови) эндокохлеарные и эндовестибулир-ные потенциалы. Роль их в работе слухового анализатор;! пока не выяснена. На базилярной мембране располагается рецептор звуковою анализатора — кортиев орган (см. рис. 15.12, В), состоящий hi четырех рядов волосковых клеток — одного внутреннего и трех наружных. На верхней части рецепторных клеток кортиеиого органа находится несколько десятков волосков — стереоцилии. Ниже рейснеровой мембраны (базилярной) от костной стенки улитки отходит текторальная (покровная) мембрана, нависающая над рядами расположенных под ней волосковых клеток. Стереоцилии волосковых клеток трех наружных рядов контактируют с текторальной мембраной, а внутреннего ряда с ней контакта не имеют. Все упоминаемые выше структуры представляют собой структуры вспомогательного аппарата (рис. 15.13). Они предназначены передавать с минимальными искажениями звуковые колебания к рецепторным волосковым клеткам. Волосковые клетки имеют длину 20...40мкм и диаметр 5...10мкм. Стереоцилии на апикальной части рецепторных клеток имеют длину 4...10мкм и диаметр 0,1...0,3мкм. Сердцевину стереоцилии образует пучок фибрилл, которые сверху окружены клеточной мембраной. У основания стереоцилии фибриллы сливаются и оканчиваются внутри клетки в кутикулярной пластинке. Стереоцилии у волосковых клеток трех наружных рядов располагаются в виде подковообразной фигуры, причем самые длинные находятся на вершине, а внутреннего ряда — на апикальной поверхности в несколько рядов. Кроме волосковых клеток в кортиевом органе имеются несколько типов опорных клеток. Первичный афферентный нейрон контактирует с волосковыми клетками с помощью синапти-ческих окончаний. К рецепторным клеткам подходят и эфферентные нервные волокна, образуя соответственно эфферентные синапсы (звено обратной связи). Афферентные волокна слухового нерва генерируют спонтанные потенциалы действия, и частота их иногда бывает достаточно высокой (более 10 имп/с). Наличие спонтанной импульсации связывается с фоновым выделением медиатора в синаптической области волосковых клеток, что, в свою очередь, предполагает деполяризацию мембраны волосковых клеток. Микроэлектродные измерения показали, что действительно мембранный потенциал волосковых клеток сдвинут на 40...60 мВ в деполяризационную сторону от равновесного калиевого потенциала, что может быть одной из причин высокой чувствительности слухового анализатора. Колебания жидкости в улитке, вызванные стимуляцией стремечком мембраны овального окна, передаются базилярной мемб-
-0,1 0 +0,1 Смещение, мкм Г Рис. 15.13. Схема возбуждения волосковых клеток и генерации в них электрической активности: А. Возникновение сдвигающей силы, воздействующей на стереоцилии рецепторных клеток при отклонении базилярной мембраны вверх. Скольжение текторальной мембраны по кортие-вому органу обусловлено тем, что текторальная и базилярная мембраны имеют разные точки вращения. Перемещения мембраны даны в сильно увеличенном масштабе; Б. Волосковая ре-цепторная клетка и ее электрическая активность при механическом раздражении; В. Рецептор-ный потенциал и афферентная импульсная активность при разном направлении механического раздражения, воздействующего на стереоцилии; Г. Зависимость между амплитудой смещения стереоцилии и величиной рецепторного потенциала ране, на которой покоится кортиев орган. При этом происходит механическое смещение волосков на апикальном конце рецептор-ной клетки, активация механоактивируемых каналов и возникновение рецепторного потенциала. Полагают, что сдвиг стереоци- лий обусловлен различным расположи и иг м мсмцмт и|мн«ним ш зилярной мембраны и текторальной мембраны, i ин.ц.нм и|.м креплены наружные волосковые клетки (см. рис I'» И, Ь I. t.u.i даря такому расположению отклонение базилярной мемприиы вверх вызывает смещение кончиков стереоцилии в напржим ним их вершины. При этом происходит усиление натяжения мсмп раны и активация механоактивируемых ионных каналов. И ни оборот, смещение базилярной мембраны вниз вызовет наклон стереоцилии в противоположном направлении, что уменьшит натяжение рецепторной мембраны. Об этом свидетельствуют данные, полученные при изучении одиночных волосковых клеток (см. рис. 15.13,5). Смещение пучка стереоцилии в сторону наиболее длинных стереоцилии, находящихся на вершине подковообразного расположения стереоцилии, вызывает деполяризацию мембраны рецепторной клетки. Смещение же стереоцилии в другую сторону гиперполяризует мембрану (см. рис. 15.13, В). Таким образом, в покое часть механоактивируемых каналов открыта и ослабление натяжения мембраны вызывает их инактивацию и уменьшение ионного тока. На изолированных волосковых клетках с помощью методики локальной фиксации было изучено действие отдельных механоактивируемых каналов. Реакция механоактивируемых каналов слуховых волосковых клеток на механическое раздражение, как и других ионных каналов, представляет собой ступенчатообразные колебания тока различной длительности, но одинаковой амплитуды. Зависимость суммарного ионного тока волосковых клеток от амплитуды механического стимула носит логарифмический характер (см. рис. 15.13, Г). Другой этап работы периферического звена слухового анализатора заключается в кодировании информации о высоте (частоте) звука. Животные способны воспринимать звуки с частотой 15...80000 Гц. При частоте звука, превышающей 1000 Гц, однозначного соответствия между звуковыми колебаниями и частотой афферентных потенциалов действия не может быть, что обусловлено наличием абсолютного рефрактерного периода после каждого потенциала действия. Поэтому информация о частоте звука должна кодироваться не частотой потенциалов действия, а с помощью другого механизма. Изучая морфологические и механические свойства базилярной мембраны, Г. Бекеши в 60-х годах XX в. пришел к выводу, что амплитуда смещения различных ее участков зависит от частоты звука. Причем колебания в базилярной мембране распространяются в виде бегущей волны. Постепенное увеличение механической податливости базилярной мембраны от ее узкого конца к широкому в дистальной части улитки приводит к тому, что амплитуда бегущих по ней волн изменяется (рис. 15.14). Точка, в которой смещение мембраны имеет наибольшую амплитуду (и соответственно происходит максимальная стимуля-
ция стереоцилий волосковых клеток), зависит от частоты бегущих волн, а следовательчастоты звука. При воздействии высоких час- смещение базилярной мембраны ближе к овальному окну. Для низкочастотных сигналов область наибольшего смещения базилярной мембраны сдвигается к дальнему концу улитки. Таким образом, принцип кодирования частоты звука — пространственный. В то же время кодирование громкости (амплитуды) звукового стимула осуществляется частотным кодом. Степень смещения базилярной мембраны в любой точке ее длины определяет силу стимуляции волосковых клеток и тем самым амплитуду рецепторного потенциала, что, в свою очередь, обусловливает частоту импульсов в афферентных нервных волокнах, иннервирующих данную группу клеток. У волосковых клеток, как и в других механорецепторах, в ответ на длительное действие механического стимула наблюдается адаптация рецепторного потенциала. Это вызывает уменьшение выброса медиатора и афферентной импульсации. Данное явление лежит в основе адаптации слуха, т. е. временном снижении чувствительности слухового анализатора при длительном действии звукового сигнала. Первыми афферентными нейронами слуховой системы млекопитающих являются клетки спирального ганглия улитки. Периферические отростки этих клеток направляются в виде спиральных и радиальных пучков к рецепторным клеткам кортие-вого органа и образуют на них синапсы. Аксоны этих нейронов формируют улитковую, или кохлеарную, ветвь слухового нерва, который, пройдя через внутренний слуховой проход, входит в мозговой ствол и заканчивается в кохлеарных ядрах мозга. Улитка проецируется на кохлеарные ядра таким образом, что волокна от базального высокочастотного завитка улитки оканчиваются в дорсомедиальной части кохлеарных ядер, а от апикального — низкочастотного — в вентролатеральной части. Тела афферентных нейронов третьего порядка находятся в задних буграх четверохолмия и в медиальном коленчатом теле тала-мической области. Внутреннее коленчатое тело — это талами-
ческий центр слуховой системы. Аксоны кисни, шимккипм коленчатого тела оканчиваются на клетках с-иумити ш.ы. ш коры — конечного этапа слухового пути, распиши,ионн м*. ч и височной доле больших полушарий. Благодаря наличии) л\\\\ ушей животные точно определяют направление, откудп i........ 1 сится звук. Это явление получило название бинауралышгп >ф фекта. В височных долях коры происходит анализ звуковых ко лебаний и создается представление о направленности звука При двустороннем разрушении слуховой коры пространственный слух нарушается. Дата добавления: 2015-12-15 | Просмотры: 1034 | Нарушение авторских прав |

 /V**"""°V^\ Текториальная клетки волосковые клетки
/V**"""°V^\ Текториальная клетки волосковые клетки