|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ВКУСОВОЙ АНАЛИЗАТОРВкусовой анализатор сообщает животному о химических свойствах корма, поступающего в его ротовую полость. Вкусовые рецепторы в отличие от обонятельных сгруппированы в специальные образования, по форме напоминающие луковицы, — вкусовые луковицы. В свою очередь, подавляющее число вкусовых луковиц собраны в особых выростах эпителия языка — со-
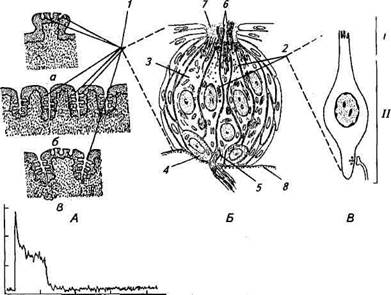
Г Рис.15.11. Структурная и функциональная организация вкусового рецептора: А. а — грибовидный сосочек; б — листовидные сосочки; в — желобовидный сосочек; прямыми линиями указано место расположения вкусовых луковиц (1) в сосочках. Б. Схема ультраструктурной организации вкусовой луковицы языка кролика: 1 — вкусовая луковица; 2 — рецептор-ная клетка; 3 — опорная клетка; 4— базальная клетка; 5—афферентный синапс; б—микроворсинки рецепторной клетки; 7—пора; 8— базальная мембрана. В. Рецепторная клетка: /— апикальная область рецепторной клетки, где происходит взаимодействие вкусовых веществ с белками мембраны микроворсинок. На апикальной мембране ионные каналы связаны с рецепцией кислых и соленых веществ; //— базолатеральная область рецепторной мембраны. Ионные каналы связаны с рецепцией сладких веществ, которые активируются внутриклеточным посредником — цАМФ; Г. Изменение частоты потенциалов действия афферентного нервного ствола (барабанной струны) при действии на вкусовые луковицы 0,1 м раствора хлорида натрия (соленый вкус). По оси ординат — частота, имп/с, по оси абсцисс — время, с; внизу—время действия солевого раствора 10 с сочках (рис. 15.11, А). Сосочки имеют различную форму, и их подразделяют на грибовидные, желобовидные и листовидные. Грибовидные сосочки выступают над поверхностью языка и напоминают по форме гриб; расположены в основном на передней дорсальной поверхности языка. Листовидные сосочки состоят из нескольких вертикальных складок, расположенных параллельно, в виде листочков; сконцентрированы по обеим сторонам задней поверхности языка. Желобовидные сосочки погружены в толщу слизистой языка и отделены от нее кольцевидным желобком; локализованы на дорсальной части у корня языка. Редкие вкусовые луковицы найдены также в слизистой оболочке
42 — 3389
Вкусовая луковица (см. рис. 15.11, Б) расположена в толще многослойного эпителия и ориентирована перпендикулярно слоям эпителия так, что апикальный отдел каждой луковицы сообщается с окружающей средой, а базальная часть граничит с соединительной тканью, отделенной базальной мембраной. В состав вкусовой луковицы входят несколько типов клеток: вкусовые рецепторные клетки, опорные клетки и базальные клетки. Вкусовая клетка имеет веретенообразную форму; диаметр в широкой части б.-ЛОмкм (см. рис. 15.11, В). Апикальный отросток более длинный и тонкий, базальный — короткий, иногда делится и формирует синапсы с афферентными волокнами. На вершине апикального отростка имеются микроворсинки диаметром около 0,2 и длиной 4 мкм. Вкусовые рецепторные клетки в верхней части образуют плотные контакты с опорными клетками так, что имеется барьер между наружной и межклеточной средой латерального и базального участков клеток. На верхней части опорных клеток микроворсинки, по размерам сходные с микроворсинками ре-цепторных клеток. Базальные клетки находятся в нижней части вкусовой луковицы и формируют плотные контакты с опорными клетками. Верхняя часть луковицы сообщается с наружной средой через узкий канал — пору. Во вкусовых луковицах идет постоянный процесс обновления клеток. Клетки непрерывно делятся ми-тотически и дегенерируют, заменяясь новыми. Так, вкусовая клетка в среднем живет (250 + 50) ч. Молодые вкусовые клетки движутся от периферии к центру, заменяя старые клетки. Кроме афферентных нервных волокон к вкусовым луковицам подходят и эфферентные нервные волокна, образуя соответственно на ба-зальных участках вкусовых клеток эфферентные синапсы (звено обратной связи). Большинство нервных волокон, образующих си-наптические контакты с рецепторными клетками вкусовых луковиц, локализованных в передних двух третях языка, входят в состав язычного нерва. Нервные волокна, иннервирующие вкусовые луковицы задней трети языка, идут в ЦНС в составе языко-глоточного нерва. Небольшая часть афферентных волокон от вкусовых клеток включается в состав других черепномозговых нервов: тройничного, лицевого и блуждающего. Афферентные волокна от вкусовых луковиц, идущие в составе язычного нерва, на некотором расстоянии выходят из него и образуют основную часть нервного ствола, получившего название барабанной струны — horda tympani. Поскольку horda tympani состоит главным образом из вкусовых афферентных нервных волокон, ее используют для электрофизиологического исследования функций вкусовых рецепторных клеток. При регистрации электрической активности от нервных волокон horda tympani (см. рис. 15.11, Г) было обнаружено, что в них в отсутствие влияния раздражителя на вкусош.н' нумипши и -in |>н руются редкие потенциалы действия. Это служи i in. №................. „, казательством того, что клеточная мембрана вкусоиои ► и к и и. поляризована и происходит спонтанный выброс мпшницш»н пресинаптической области вкусовой клетки. Микро^лгкфмммыг измерения показали, что мембранный потенциал клеток елнимуi на 40...50мВ в сторону деполяризации по сравнению с paimoiu-i ным потенциалом для ионов калия, а это способствует увеличс нию чувствительности рецептора. Что же служит адекватным раздражителем для вкусовых клеток языка? Для человека и животных выделяют пять основных вкусовых ощущений: сладкое, горькое, кислое, соленое и вкус пищи животного происхождения, обусловленный присутствием аминокислот, таких, как, например, глютамат. Иногда это вкусовое ощущение называют японским термином умами. Вкусовые ощущения возникают в том случае, если вещество растворяется в слюне и затем воздействует на апикальную область вкусовых рецепторных клеток. На действие веществ, обладающих различным вкусом, клетки реагируют деполяризацией мембраны — генерацией рецеп-торного потенциала, связанной с изменением ионной ее проводимости. Рецепторный потенциал увеличивает амплитуду с повышением концентрации стимулирующего вещества; эта зависимость носит логарифмический характер. Несмотря на трудности с нанесением дозированных химических стимулов на рецептор, удалось показать, что у рецепторного потенциала вкусовых клеток имеются динамическая и статическая фазы и соответственно быстрая и медленная адаптации. Особенно это хорошо заметно при регистрации афферентных потенциалов действия в нервных волокнах horda tympani (рис. 15.11, Г). Частота афферентных потенциалов действия максимальна в начале ответа, далее следует спад, сначала сравнительно быстрый, а затем частота потенциалов действия медленно снижается до уровня, близкого к исходному значению частоты спонтанной импульсации. Несомненно, с адаптацией связано постепенное снижение и исчезновение в некоторых случаях ощущения вкуса того или иного вещества. Учитывая большие вариации в химической природе веществ, вызывающих вкусовые ощущения, встает вопрос, за счет каких рецепторов апикальной мембраны и каким образом осуществляется рецепция специфических веществ. Ответы на эти вопросы были частично получены в конце XX в. в опытах на изолированных вкусовых клетках различных животных. Оказалось, что рецепция химических веществ осуществляется несколькими механизмами. Известно, что кислый вкус связан с воздействием на вкусовые рецепторы кислот, причем степень «кислости» зависит в основном от концентрации ионов водорода. Действие кислот сопровождается возникновением деполяризационного потенциала 42* 659
При действии на язык хлорида натрия возникает «классический» соленый вкус. Другие соли (по крайней мере для человека) имеют различные привкусы (горечи и т. д.). Рецепция соленого связана с наличием на апикальной мембране потенциалнеза-висимых ионных натриевых каналов. Увеличение концентрации ионов натрия в слюне сопровожается появлением входящего внутрь клетки ионного натриевого тока и деполяризацией мембраны. Рецепцию веществ, имеющих сладкий вкус, обеспечивают специальные рецепторные белки, находящиеся на апикальной мембране. Так же как и в обонятельных клетках ионные каналы, за счет которых происходит деполяризация мембраны, пространственно отделены от «сладких» рецепторных белков. В передаче сигнала участвует внутриклеточный посредник — цАМФ, который снижает калиевую проводимость мембраны посредством блокирования калиевых каналов, расположенных в базолатеральной мембране клетки, в отличие от рецепции кислого. Механизм рецепции горького и аминокислотного (умами) вкуса расшифрован в меньшей мере. В апикальной мембране вкусовых клеток имеются рецепторные белки для этих веществ: внутриклеточным посредником для рецепции горького вкуса выступают ионы кальция, а для аминокислот — цАМФ. При проведении экспериментов на одиночных вкусовых клетках по изучению ионных каналов в различных участках мембраны была обнаружена еще одна особенность. В базальных областях клеток имеются потенциалзависимые каналы, за счет которых вкусовая клетка в ответ на деполяризацию генерирует потенциалы действия. Таким образом, вкусовая клетка представляет собой видоизмененный первичный афферентный нейрон и последовательность событий в ней при рецепции химического стимула совпадает с таковой в обонятельных клетках. Наряду с вкусовыми клетками, реагирующими на одно специфическое вкусовое вещество, были обнаружены клетки, которые возбуждаются двумя или тремя вкусовыми веществами. Животные часто по-другому реагируют на вкусовые вещества, чем человек, субъективные реакции которого используются для объяснения работы вкусовых клеток. Так, неприятный для человека горький вкус не вызывает у некоторых видов животных отрицательного отношения к нему: кролики и лошади охотно поедают горькую осиновую кору; кислый вкус лимона весьма неприятен для лошадей, коров, собак. Центральные отростки нейронов, иннервирующие вкусовые луковицы, оканчиваются в продолговатом мозге в ядре одиночного пучка, где происходит переключение на нейроны следующею порядка. Аксоны этих нейронов образуют частичный перекрест и ретикулярной формации и достигают вентромедиального комплекса ядер таламуса. Из таламуса аксоны нейронов третьего порядка направляются в кору головного мозга в корковый центр вкуса, локализация которого пока точно не определена. Чувствительность вкусовых клеток контролирует ЦНС с помощью эфферентной цепи обратной связи посредством эфферентных синапсов. У животных вкусовая чувствительность меняется в зависимости от функционального состояния желудочно-кишечного тракта. Сигналы с рецепторов слизистой желудка идут в ЦНС, которая регулирует возбудимость вкусовых рецепторов. Развитие вкусового анализатора у животных определяется составом поедаемой пищи. В наибольшей степени развит вкусовой анализатор у травоядных животных, так как их пища включает большое количество трав различных видов, среди которых они должны выбирать съедобные растения. У плотоядных животных пища более однообразна: мясо не может принести им вреда, поэтому и не возникает необходимость тонко различать его вкус. Дата добавления: 2015-12-15 | Просмотры: 833 | Нарушение авторских прав |
 задней поверхности ротовой полости, на стенках глотки, миндалин, надгортанника и гортани.
задней поверхности ротовой полости, на стенках глотки, миндалин, надгортанника и гортани. во вкусовых клетках. Причиной деполяризации рецепторной мембраны является блокирование ионами водорода потенциал-зависимых калиевых каналов, расположенных на апикальной мембране вкусовой клетки. Согласно уравнению Голдмана (см. гл. 2) уменьшение проницаемости для ионов калия сдвигает мембранный потенциал в сторону деполяризации.
во вкусовых клетках. Причиной деполяризации рецепторной мембраны является блокирование ионами водорода потенциал-зависимых калиевых каналов, расположенных на апикальной мембране вкусовой клетки. Согласно уравнению Голдмана (см. гл. 2) уменьшение проницаемости для ионов калия сдвигает мембранный потенциал в сторону деполяризации.