|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
МЫШЕЧНОЕ ВЕРЕТЕНО
Мышечные веретена принадлежат к группе наиболее изученных рецепторов. Поэтому следует более подробно рассмотреть их характеристики с целью дальнейшего их использования при изучении свойств рецепторов других модальностей. Под световым микроскопом мышечное веретено представляет собой вытянутую структуру, расширенную в середине за счет капсулы и напоминающее по форме веретено, что и обусловило его название. Мышечные веретена у всех видов животных имеют примерно одинаковое строение (рис. 15.2). Вспомогательные структуры мышечных волокон представлены специализированными поперечнополосатыми мышечными волокнами, названными интрафузалъными (в отличие от обычных, экстрафузальных волокон), а также капсулой рецептора, за-
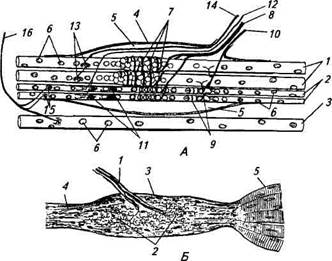
Рис. 15.2. Строение мышечного веретена (А) и сухожильного ре цептора Гольджи (Б): А. 1 — интрафузальное мышечное волокно с ядерной сумкой; 2 — интра-фузальное мышечное волокно с ядерной цепочкой; 3 — экстрафузальное мышечное волокно; 4— капсула; 5 — внутрикапсулярное пространство; 6— ядра мышечных волокон; 7—первичные афферентные окончания; 8— афферентное нервное волокно первого типа; 9— вторичные нервные окончания; 10— афферентное нервное волокно второго типа; 11 — у-кус-тиковидные эфферентные нервные окончания; 12— у2-двигательное нервное волокно; 13 — у-пластинчатые эфферентные нервные окончания; 14 — у [-двигательное нервное волокно; 15 — р-пластинчатые эфферентные нервные окончания; 16— р-двигательное нервное волокно; Б. 1 — афферентное нервное волокно; 2— афферентные нервные окончания; 3— капсула; 4— сухожилия; 5— мышечные волокна полненной жидкостью. В полярных областях интрафузальных волокон хорошо видны ядра, рассеянные по периферии волокна, как и в экстрафузальных. Ближе к центральной части, где волокно покрыто капсулой, появляются дополнительные ядра. Это и послужило основанием называть центральную зону волокна областью ядерной сумки, а сами интрафузальные волокна называют волокнами с ядерной сумкой; диаметр этих волокон составляет 20...25мкм. У более тонких волокон другого типа (диаметр около 10 мкм) ядра располагаются в ряд, образуя цепочку. Такие волокна получили название интрафузальных волокон с ядерной цепочкой. Интрафузальные волокна покрыты соединительнотканной капсулой. В центральной области веретена, где капсула отходит от интрафузальных волокон, имеется перикапсулярное пространство, заполненное жидкостью. К мышечному веретену подходит большое количество нервных волокон, разветвляющихся в разных его час- тях, но основная сеть разветвлений находится и ofttiaiin,.......... фм той капсулой. В мышечных веретенах было обнаружено штышьт типов афферентных и эфферентных двигательных окончании Последние согласно общей схеме строения анализатором можно отнести к звену обратной связи. Афферентные окончания представлены двумя типами. 11 с р вый тип (первичные окончания) образуется разветвлени ями довольно толстых миелинизированных волокон, достигающих в диаметре около 20 мкм. Нервное волокно проникает под капсулу и, многократно разветвляясь, постепенно теряет миелиновую оболочку. Первичные окончания локализуются в середине центрального участка интрафузальных волокон и имеют вид спиралей и колец, обвивающих волокна. Второй тип (вторичные окончания) образуется более тонкими миелинизи-рованными волокнами (диаметр 4... 12 мкм) и располагается ближе к периферии интрафузального волокна (см. рис. 15.2). Диаметр обоих типов окончаний в поперечном сечении составляет 0,5...3 мкм. Характерная их особенность — это большое число митохондрий во внутриклеточном пространстве. Мышечные веретена иннервируют также и несколько типов двигательных нервных волокон, получивших название фузимоторных (у-волокон). Фузимоторные синаптические окончания, образованные этими волокнами, подразделяются на у-кустиковид-ные и у-пластинчатые. Первые локализуются ближе к центру веретена, а у-пластинчатые — дистальнее. Кроме двигательных у-волокон мышечные веретена дополнительно иннервируют двигательные Р-волокна, которые одновременно образуют синаптические окончания на экстрафузальных мышечных волокнах, р-волокна на интрафузальных волокнах формируют окончания пластинчатого типа, располагающиеся в самых ди-стальных областях интрафузальных волокон. В исследованиях реакции интрафузальных мышечных волокон на раздражение эфферентных волокон было обнаружено, что способностью к сокращению обладают только дистальные участки, а центральная часть несократима. При регистрации электрической активности в афферентных нервных волокнах, идущих от мышечных веретен, в состоянии покоя были обнаружены нерегулярные низкочастотные потенциалы действия, получивших название спонтанных. Принимая во внимание механизмы генерации потенциалов действия в нервных клетках и волокнах (см. гл. 2), можно полагать, что мембрана рецеп-торной области исходно деполяризована и значение мембранного потенциала находится в области порога возникновения потенциалов действия. Таким образом, даже самое маленькое увеличение энергии раздражающего стимула (в данном случае механического) будет повышать частоту импульсации рецептора и его чувствительность очень высока.
Рис. 15.3. Электрические ответы мышечного веретена на ступенчатообраз-ное механическое растяжение: А. / — электрические ответы первичных афферентных нервных окончаний; 2 — отметка ступенчатообразного механического растяжения (калибровка времени: внизу горизонтальная линия соответствует 0,5 с); 3 — электрические ответы вторичных афферентных нервных окончаний; 4 — отметка ступенчатообразного механического растяжения (калибровка времени: внизу горизонтальная линия — 0,5 с); стрелками отмечены спонтанные потенциалы действия. Б. Потенциалы действия заблокированы введением в раствор родотоксина. Рецепторные потенциалы первичных (1, 2, 3) и вторичных (4, 5, 6) афферентных нервных окончаний мышечного веретена в ответ на растяжение ступенчатообразными механическими стимулами с различной скоростью нарастания (7, 8, 9) переднего фронта механического растяжения; калибровка времени: горизонтальная линия соответствует 0,5 с Растяжение мышечного веретена приводит к возникновению электрической активности — рецепторного потенциала и потенциалов действия (рис. 15.3). При легком ступенчатообразном растяжении мышечного веретена в первичных и во вторичных афферентных окончаниях регистрируется длительная серия потенциалов действия. Частота разрядов зависит от степени растяжения мышцы. На рисунке видно, что потенциалы действия находятся на своеобразном деполяризационном пьедестале, представляющем собой рецепторный потенциал, т. е. изменение мембранного потенциала афферентных окончаний. Более сильное растяжение приводит к более значительной деполяризации и увеличению частоты афферентных потенциалов действия. Частота потенциалов действия мышечного веретена на фоне неизменной амплитуды растяжения изменяется со временем. Начало растяжения сопровождается повышением частоты импульсов с последующим быстрым спадом до некоторого уровня (динамическая часть ответа). Далее частота продолжает уже медленнее снижаться до значения, определяемого величиной растяжения (статическая часть ответа). Процесс снижения частоты потенциалов действия получил название адаптации. Таким образом, в ответе мышечного веретена четко выражены динамическая и статическая фазы, во время которых соответственно происходит быстрая и медленная адаптация. Подобная реакция на длительный адекватный стимул характерна и для рецепторов других модальностей. Данный тип рецептором получил название медленноадаптирующихся. Необходимо отмети., что явление адаптации в анализаторах имеет чрезвычайно илжпое значение для процесса восприятия раздражающего стимула, (о гласно закону специфических энергий, изменения в частого им пульсной активности афферентного нейрона означают изменение ощущения, возникающего в ЦНС. Работа двигательного анализатора у человека в нормальных условиях не сопровождается возникновением каких-либо определенных ощущений. Косвенные данные указывают на то, что и у животных двигательный анализатор функционирует сходным образом. Поэтому адаптация рецепторов мышц не проявляется в ощущении. Однако в других анализаторах явление адаптации весьма заметным образом влияет на восприятие раздражения. Например, в зрительном, обонятельном, кожном анализаторах соответственно интенсивность ощущения яркого света, сильного запаха или пониженной температуры со временем снижается. Изучение механизмов адаптации в рецепторах различных анализаторов показало, что адаптация может происходить на разных этапах восприятия раздражения (см. рис. 15.1): от воздействия стимула на вспомогательный аппарат до генерации афферентных потенциалов действия. Причем у различных рецепторов вклад в адаптацию структур, входящих в их состав, неодинаков. Первым звеном, которое определяет адаптацию у некоторых рецепторов, служит вспомогательный аппарат. У мышечных веретен это интрафузальные мышечные волокна. Как известно (см. гл. 2), мышечные волокна включают в себя эластичный и вязкий элементы, по-разному реагирующие на растяжение. Под действием растяжения вначале происходит удлинение центральной части с соответствующей реакцией на это афферентных окончаний данной области, а затем дистальных участков веретена с раздражением контактирующих с ними афферентных окончаний: При этом длина центральной части уменьшается. Соответствующим образом изменяются амплитуда рецепторного потенциала и частота афферентных потенциалов действия в нервных волокнах. Например, в первичных окончаниях, расположенных преимущественно в центре веретена, за первоначальным повышением рецепторного потенциала и частоты потенциалов действия следует быстрое снижение (быстрая адаптация). В дальнейшем натяжение будет ослабляться медленнее и частота потенциалов действия начнет снижаться также медленнее (медленная адаптация). Параллельно с чисто механическим фактором в адаптации мышечного веретена принимают участие и ионные процессы, лежащие в основе генерации рецепторного потенциала. Сравнительные исследования реакции первичных и вторичных афферентных окончаний мышечных веретен показали, что на стационарную часть растяжения они реагируют примерно одинаково (см. рис. 15.3). Однако на возрастание растяжения в начале действия стимула ответы их отличаются. Как видно на рисунке, у первичных окончаний динамическая часть ответа более выражена. С возрастанием скорости растяжения увеличиваются амплитуда динамической части рецепторного потенциала и соответственно частота потенциалов действия, приходящихся на этот период. У вторичных окончаний динамическая часть ответа выражена в меньшей степени. Таким образом, вторичные афферентные окончания главным образом кодируют (детектируют) амплитуду растяжения — детекторы амплитуды, а первичные окончания — амплитуду и скорость нарастания растяжения — детекторы скорости амплитуды. При введении в раствор, омывающий мышечное веретено, тет-родотоксина — специфического блокатора потенциалзависимых натриевых каналов (см. гл. 2) — потенциалы действия подавляются. Однако рецепторный потенциал, имеющий ступенчатообразную форму с характерным деполяризационным выбросом, в начале своего возникновения (динамическая часть ответа) сохраняется и не изменяется со временем. Это указывает на разные механизмы генерирования рецепторного потенциала и потенциалов действия, возникающих по закону «все или ничего». На этом мы остановимся подробнее далее. Усиление интенсивности адекватного стимула (растяжения) сопровождается повышением как амплитуды рецепторного потенциала, так и частоты афферентных потенциалов действия. Диапазон интенсивностей стимула, в котором рецептор способен кодировать амплитуду стимула, генерируя с увеличением энергии стимула все более интенсивные сигналы, называется динамическим диапазоном. В мышечных веретенах амплитуда рецепторного потенциала приблизительно пропорциональна логарифму интенсивности стимула (рис. 15.4.) В тоже время средняя частота афферентных импульсов связана более или менее линейной зависимостью с амплитудой рецепторного потенциала вплоть до того значения, ког-
Логарифм интенсивности стимула
В
Рис. 15.4. Зависимость между амплитудой рецепторного потенциала и логарифмом величины растяжения (А), частотой импульсов и амплитудой рецепторного потенциала (Б), частотой импульсов и логарифмом величины растяжения (В) в мышечных веретенах да явление рефрактерности потенциалов действия (см. гл. 2) ним нет ограничивать интервал между потенциалами дейстнин и, следовательно, частоту импульсации. Исходя из этих двух чаши и мостей, частота потенциалов действия является функцией лот рифма интенсивностей стимула в медленноадаптирующихси рецепторах. Учитывая тот факт, что степень растяжения мышцы может изменяться в довольно широких пределах, становится понятным, насколько важна для мышечных веретен логарифмическая зависимость между амплитудой стимула и частотой афферентных потенциалов действия. Эта зависимость также имеет чрезвычайно большое значение для восприятия раздражения у рецепторов других модальностей. Так, например, солнечный свет примерно в 1019 раз ярче лунного, а слуховая система человека и животных способна воспринимать без значительного искажения звуки, энергия которых может различаться в 1012 раз. Здесь надо отметить, что способность анализаторов функционировать в больших диапазонах интенсивности внешних стимулов определяется не только большим диапазоном первичного (рецепторного) преобразования стимулов, но и наличием в центральных структурах анализатора механизмов, дополнительно расширяющих динамический диапазон. Логарифмическая зависимость «раздражение — ответ» (см. рис. 15.4) «сжимает» область высоких интенсивностей, тем самым значительно расширяя диапазон, в котором возможно различение стимулов по амплитуде. Вследствие логарифмической зависимости между величиной рецепторного потенциала и силой раздражения данное относительное усиление стимула приводит к одинаковому приращению рецепторного потенциала в очень широком диапазоне интенсивностей. Например, удвоение силы слабого раздражающего стимула приведет к такому же приращению амплитуды рецепторного потенциала, как и при удвоении силы стимула в области высоких интенсивностей. Если обозначить интенсивность стимула /, а приращение стимула Д/, то отношение приращения стимула к его амплитуде будет величиной постоянной, т. е. Al/I= const. Какова же природа рецепторного потенциала мышечных веретен? Эксперименты на мышечных веретенах с вариацией ионного состава в наружном растворе показали, что рецепторный потенциал генерируется в основном за счет ионов натрия. Однако в растворе с тетродотоксином рецепторный потенциал не подавляется (см. рис. 15.3, Б). Это свидетельствует о том, что натриевые каналы в рецепторной мембране имеют другую структуру, чем потенциалзависимые натриевые каналы нервных волокон. По-видимому, их воротный механизм отличается от такового в потенциалзависимых натриевых каналах. Активация его происходит
/
Дата добавления: 2015-12-15 | Просмотры: 1029 | Нарушение авторских прав |



 через конформационные (пространственные) изменения, вызванные механическим воздействием. Такие каналы получили название механоактивируемых. К сожалению, методические трудности не позволяют пока изучать одиночные каналы рецепторной мембраны мышечных веретен. Однако удалось с помощью метода локальной фиксации (см. гл. 2) зарегистрировать ответы механоактивируемых ионных каналов в мембране механорецепторного дендрита мышечного рецептора растяжения речного рака. Оказа-лось,что ионный ток через одиночный механоактивируемый канал, как и в случае с потенциалзависимым и ацетилхолинактиви-руемым каналами (см. гл. 2), имеет ступенчатообразную форму различной длительности. С увеличением растяжения мембраны вероятность открытого состояния канала повышалась, причем зависимость между интенсивностью раздражающего стимула и вероятностью открытия канала была логарифмической.
через конформационные (пространственные) изменения, вызванные механическим воздействием. Такие каналы получили название механоактивируемых. К сожалению, методические трудности не позволяют пока изучать одиночные каналы рецепторной мембраны мышечных веретен. Однако удалось с помощью метода локальной фиксации (см. гл. 2) зарегистрировать ответы механоактивируемых ионных каналов в мембране механорецепторного дендрита мышечного рецептора растяжения речного рака. Оказа-лось,что ионный ток через одиночный механоактивируемый канал, как и в случае с потенциалзависимым и ацетилхолинактиви-руемым каналами (см. гл. 2), имеет ступенчатообразную форму различной длительности. С увеличением растяжения мембраны вероятность открытого состояния канала повышалась, причем зависимость между интенсивностью раздражающего стимула и вероятностью открытия канала была логарифмической.