|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ТЕРМОРЕЦЕПТОРЫ КОЖИ
Способность гомойотермных животных поддерживать постоянную температуру тела несмотря на резкие колебания температуры окружающей среды — одно из важнейших биологических регу-ляторных явлений. Процессы терморегуляции протекают независимо от сознания и не прекращаются во время сна. Начальным звеном терморегулирующей системы кожи являются многочисленные терморецепторы. Поскольку изменения температуры окружающей среды создают субъективные ощущения тепла или холода, то можно предположить, что в коже имеются соответственно тепловые и холодовые рецепторы. Действительно, в первых же объективных исследованиях терморецепции кожи, проведенных в начале 20-х годов XX в., были обнаружены особые точки, в которых при нагревании или охлаждении происходило увеличение частоты афферентных потенциалов действия в нервных волокнах, иннервирующих эти участки. По реакции на адекватный стимул температурные рецепторы были похожи на медленноадаптирую-щиеся механорецепторы кожи (рис. 15.9). В покое у обоих типов терморецепторов была зарегистрирована спонтанная импульсная активность. В ответ на ступенчатообразное изменение температуры (повышение или понижение) частота афферентных потенциалов действия быстро повышалась, достигая максимального значения, и затем сравнительно быстро понижалась до определенного уровня, а далее процесс замедлялся. Действие температурных стимулов с различными скоростями нарастания позволяет отнести
Рис.15.9. Терморецепторы кожи: A. Схематическое изображение одиночного холодового рецептора на безволосой коже носа B. Зависимость между средней частотой импульсной активности холодового (квадраты) и теп температурные рецепторы к детекторам амплитуды скорости. Исследование зависимости частоты афферентных потенциалов действия от величины температуры показало, что средняя частота импульсной активности при нагревании или охлаждении возрастает
Каковы же морфологические корреляты тепловых и Холодовых рецепторов? С конца XIX в. по 60-е годы XX в. существовало мнение, основанное на субъективных данных исследований на человеке, что холодовым рецепторам соответствуют инкапсулированные тельца Краузе, напоминающие по своему строению тельца Пачини, а тепловым — тельца Руффини. Однако объективные исследования с использованием регистрации афферентной импульсации показали, что эти рецепторы являются кожными механо-рецепторами (см. предыдущий раздел). Эксперименты с одновременной регистрацией импульсной активности и морфологической маркировкой выявили, что терморецепторам соответствуют не-инкапсулированные нервные окончания, локализованные в обоих слоях кожи. Например, холодовый рецептор безволосой части носа кошки (см. рис. 15.9, А) представляет собой неразветвлен-ное немиелинизированное окончание в эпидермисе диаметром 2,5...4мкм; расширенный его кончик заполнен мелкими митохондриями. Терморецепторы образованы тонкими миелинизиро-ванными нервными волокнами диаметром 1,5...5 мкм. Изменению частоты афферентных потенциалов действия в нервных волокнах терморецепторов должно предшествовать изменение мембранного потенциала афферентной области (рецепторно-го потенциала) (см. рис. 15.1). Известно, что формирование мембранного потенциала обусловлено пассивным и активным ионным транспортом. Величина пассивной компоненты мембранного потенциала вычисляется с помощью уравнения Голдмана. Поэтому можно предположить, что вариации в температуре будут непосредственно изменять пассивный транспорт ионов и соответственно влиять на амплитуду мембранного потенциала. Однако простые расчеты показывают, что колебание температуры на 10 °С изменяет мембранный потенциал всего лишь на 2 мВ. Если учесть, что терморецепторы обладают спонтанной активностью, т. е. мембранный потенциал находится в области порога генерации потенциалов действия, то данной величины изменения мембранного потенциала все же будет недостаточно для заметного увеличения частоты импульсации. Нужно отметить,что у некоторых видов животных тепловые рецепторы имеют чрезвычайно высокую чувствительность. Так, в лицевой ямке гремучей змеи находятся разветвленные нервные окончания, реагирующие на повышение температуры всего на 0,002 °С. Исследования последних лет позволяют полагать, что в рецепторной мембране имеются каналы, которые активируются повышением или понижением температуры. Дополнительно, что касается рецепции холода, то обширный экспериментальный материал указывает на участие в этом процессе активного ионного транспорта — ионного насоса. В частности, предполагается, что в Холодовых рецепторах при обычной температуре тела животного мембранный потенциал и рецепторных окончаниях сдвинут в сторону гиперпшшрт.шии электрогенным ионным насосом. Работа ионной помпы minimi от температуры, поэтому ее снижение, охлаждение нервной кт> минали будет затормаживать работу помпы, что вызовет деполяризацию мембраны и соответственно увеличение частоты афферентных потенциалов действия. Совокупность тепловых и Холодовых рецепторов обеспечивает функционирование системы терморегуляции организма животного. Так, интенсивное охлаждение тела возбуждает холодо-вые рецепторы. Посредством автономной нервной системы они вызывают сужение кожных кровеносных сосудов, что уменьшает интенсивность кровотока в коже и сохраняет теплоту внутри организма. Снижение теплоотдачи достигается также уменьшением поверхности тела (свертывание в клубок) и изменением положения волос или перьев на теле животного. Одновременно через систему двигательных нервов увеличивается выделение теплоты в мышцах за счет напряжения мышц и мышечной дрожи, а также посредством «разогревающих» движений. Теплопродукция возрастает, кроме того, благодаря усилению обмена веществ во внутренних органах. При внешнем перегревании тела животного тепловые рецепторы кожи вызывают процессы, обратные происходящим при переохлаждении: расширение кровеносных сосудов, усиление потоотделения, учащение дыхания. Дата добавления: 2015-12-15 | Просмотры: 885 | Нарушение авторских прав |
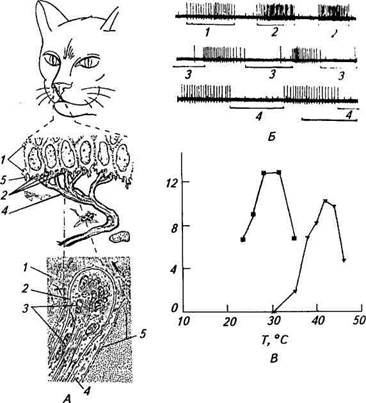
 до максимального значения, а затем в обоих случаях начинает уменьшаться до полного исчезновения. Таким образом, соотношение носит куполообразный характер (см. рис. 15.9, В).
до максимального значения, а затем в обоих случаях начинает уменьшаться до полного исчезновения. Таким образом, соотношение носит куполообразный характер (см. рис. 15.9, В).